
Investigating the Impacts of UVC Radiation on Natural and Cultured Biofilms: An assessment of Cell Viability



Abstract
:1. Introduction
2. Materials and Methods
2.1. Cultured Biofilms
2.1.1. Navicula Incerta
2.1.2. UVC Exposure
2.1.3. XTT Assay
2.2. Field Biofilms
2.3. Statistical Analysis
3. Results
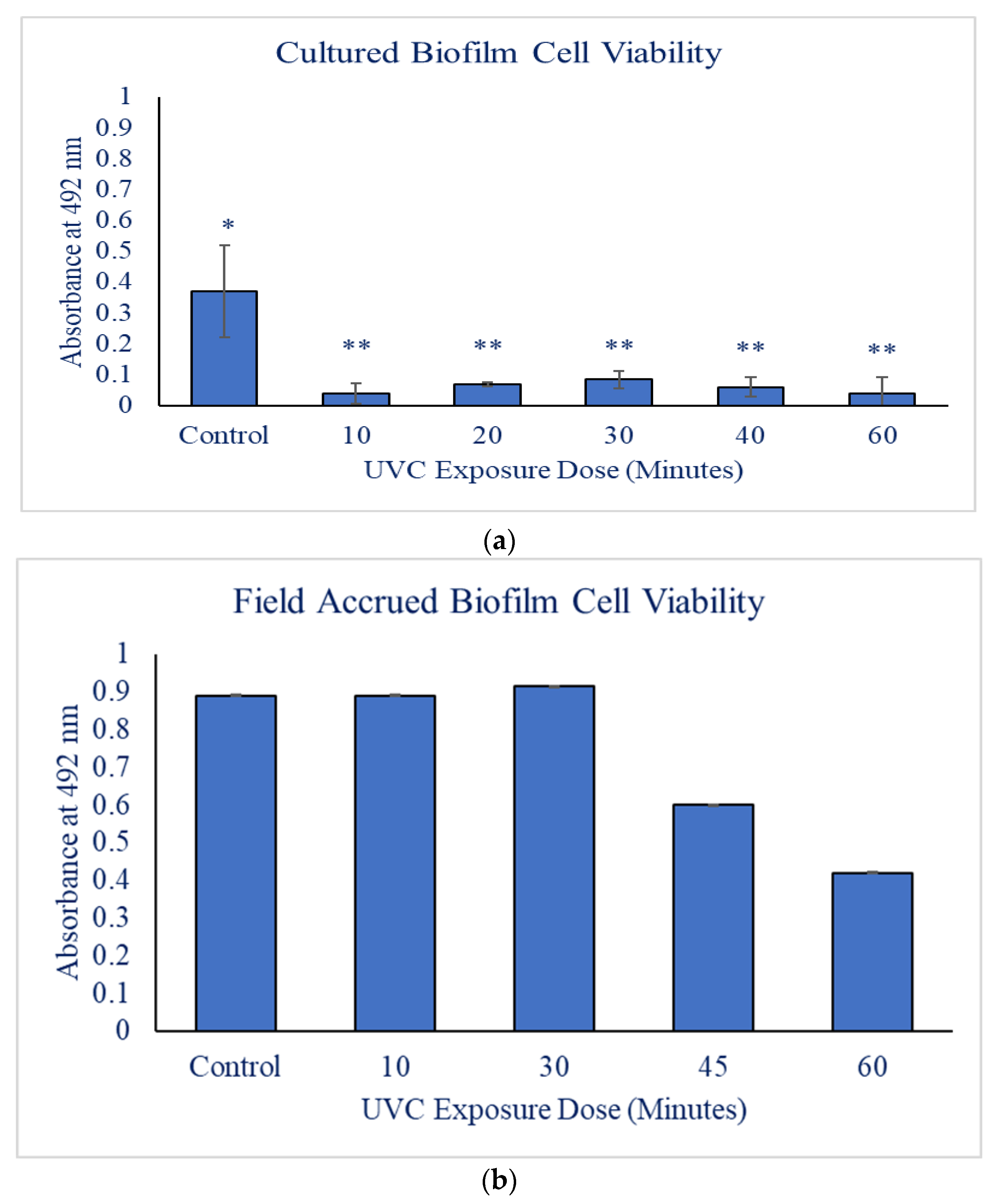
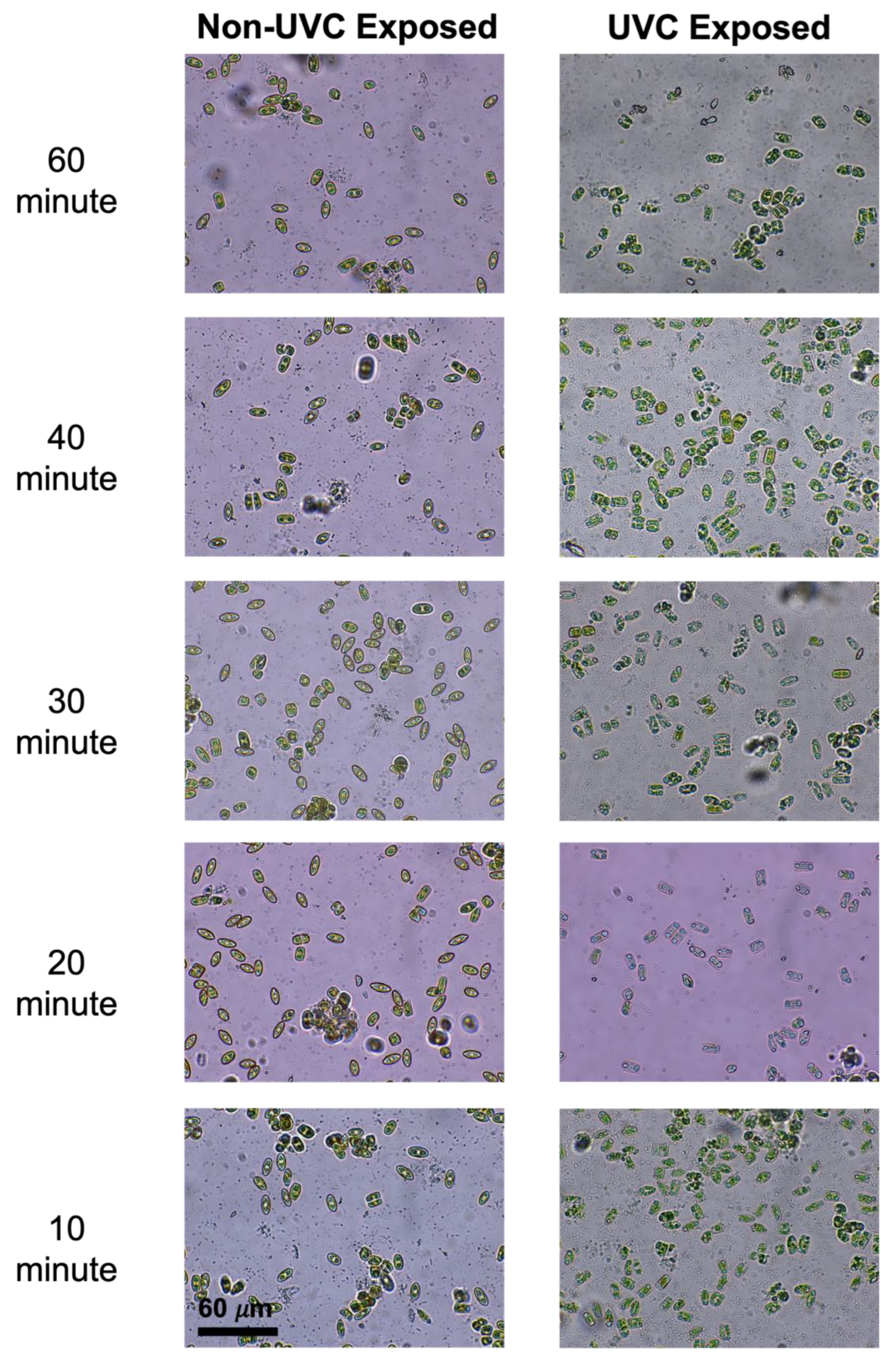
3.1. Cultured Biofilms
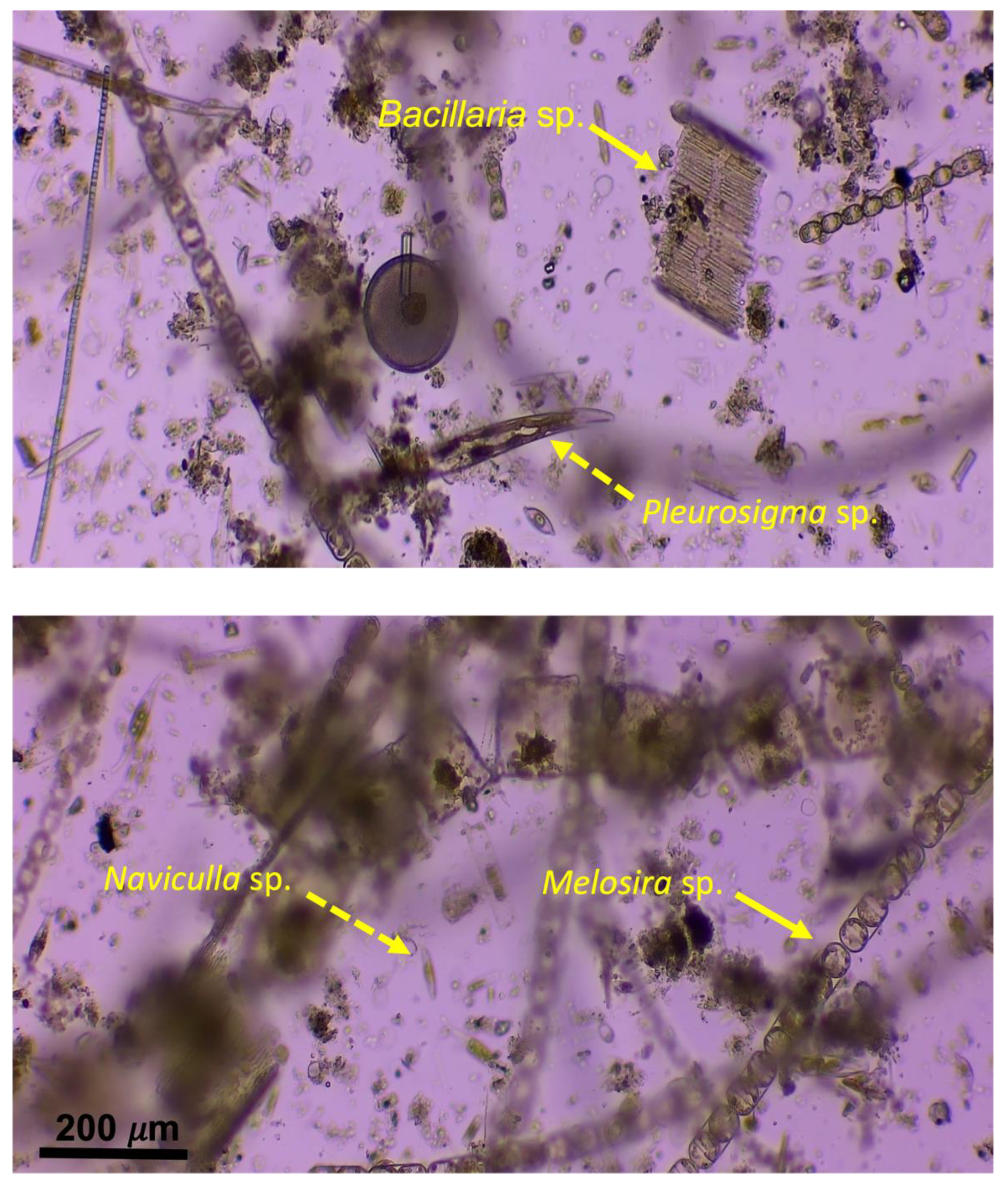
3.2. Field Biofilms
4. Discussion
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chambers, L.D.; Stokes, K.R.; Walsh, F.C.; Wood, R.J. Modern approaches to marine antifouling coatings. Surf. Coat. Technol. 2006, 201, 3642–3652. [Google Scholar] [CrossRef]
- Flemming, H.C.; Wingender, J. The biofilm matrix. Nat. Rev. Microbiol. 2010, 8, 623–633. [Google Scholar] [CrossRef] [PubMed]
- Lejars, M.; Margaillan, A.; Bressy, C. Fouling release coatings: A nontoxic alternative to biocidal antifouling coatings. Chem. Rev. 2012, 112, 4347–4390. [Google Scholar] [CrossRef]
- Qian, P.Y.; Cheng, A.; Wang, R.; Zhang, R. Marine biofilms: Diversity, interactions and biofouling. Nat. Rev. Microbiol. 2022, 20, 671–684. [Google Scholar] [CrossRef]
- Qian, P.Y.; Lau, S.C.; Dahms, H.U.; Dobretsov, S.; Harder, T. Marine biofilms as mediators of colonization by marine macroorganisms: Implications for antifouling and aquaculture. Mar. Biotechnol. 2007, 9, 399–410. [Google Scholar] [CrossRef]
- Mack, D.; Rohde, H.; Harris, L.G.; Davies, A.P.; Horstkotte, M.A.; Knobloch, J.M. Biofilm formation in medical device-related infection. Int. J. Artif. Organs 2006, 29, 343–359. [Google Scholar] [CrossRef]
- Agarwal, A.; Singh, K.P.; Jain, A. Medical significance and management of staphylococcal biofilm. FEMS Immunol. Med. Microbiol. 2010, 58, 147–160. [Google Scholar] [CrossRef]
- Analysing the Impact of Marine Biofouling on the Energy Efficiency of Ships and the GHG Abatement Potential of Biofouling Management Measures; GloFouling Partnerships and GIA for Marina Biosafety: London, UK, 2022.
- Schultz, M.P.; Walker, J.M.; Steppe, C.N.; Flack, K.A. Impact of diatomaceous biofilms on the frictional drag of fouling-release coatings. Biofouling 2015, 31, 759–773. [Google Scholar] [CrossRef] [PubMed]
- Hunsucker, K.Z.; Vora, G.J.; Hunsucker, J.T.; Gardner, H.; Leary, D.H.; Kim, S.; Lin, B.; Swain, G. Biofilm community structure and the associated drag penalties of a groomed fouling release ship hull coating. Biofouling 2018, 34, 162–172. [Google Scholar] [CrossRef]
- Omae, I. Organotin antifouling paints and their alternatives. Appl. Organomet. Chem. 2003, 17, 81–105. [Google Scholar] [CrossRef]
- Albitar, H.; Dandan, K.; Ananiev, A.; Kalaykov, I. Underwater robotics: Surface cleaning technics, adhesion and locomotion systems. Int. J. Adv. Robot. Syst. 2016, 13, 7. [Google Scholar] [CrossRef]
- Salters, B.; Piola, R. UVC Light for Antifouling. Mar. Technol. Soc. J. 2017, 51, 59–70. [Google Scholar] [CrossRef]
- Swain, G. Redefining antifouling coatings. J. Prot. Coat. Linings 1999, 16, 26–35. [Google Scholar]
- Pistone, A.; Scolaro, C.; Visco, A. Mechanical properties of protective coatings against marine fouling: A review. Polymers 2021, 13, 173. [Google Scholar] [CrossRef] [PubMed]
- Swain, G.; Erdogan, C.; Foy, L.; Gardner, H.; Harper, M.; Hearin, J.; Hunsucker, K.; Hunsucker, J.T.; Lieberman, K.; Nanney, M.; et al. Proactive in-water ship hull grooming as a method to reduce the environmental footprint of ships. Front. Mar. Sci. 2022, 8, 2017. [Google Scholar] [CrossRef]
- Swain, G.W. The importance of ship hull coatings and maintenance as drivers for environmental sustainability. In Proceedings of RINA Conference Ship Design Operation for Environmental Sustainability; Royal Institute of Naval Architects: London, UK, 2010. [Google Scholar]
- Piola, R.; Salters, B.; Grandison, C.; Ciacic, M.; Hietbrink, R. Assessing the Use of Low Voltage UV-Light Emitting Miniature LEDs for Marine Biofouling Control; Defence Science and Technology Group Fishermans Bend Victoria Australia: Port Melbourne, VIC, Australia, 2016.
- Bak, J.; Ladefoged, S.D.; Tvede, M.; Begovic, T.; Gregersen, A. Dose requirements for UVC disinfection of catheter biofilms. Biofouling 2009, 25, 289–296. [Google Scholar] [CrossRef] [PubMed]
- Parkinson, A.; Barry, M.J.; Roddick, F.A.; Hobday, M.D. Preliminary toxicity assessment of water after treatment with UV-irradiation and UVC/H2O2. Water Res. 2001, 35, 3656–3664. [Google Scholar] [CrossRef]
- Tsolaki, E.; Diamadopoulos, E. Technologies for ballast water treatment: A review. J. Chem. Technol. Biotechnol. 2010, 85, 19–32. [Google Scholar] [CrossRef]
- Braga, C.; Hunsucker, K.; Erdogan, C.; Gardner, H.; Swain, G. The Use of a UVC Lamp Incorporated with an ROV to Prevent Biofouling: A Proof-of-Concept Study. Mar. Technol. Soc. J. 2020, 54, 76–83. [Google Scholar] [CrossRef]
- Lakshmi, E.; Priya, M.; Achari, V.S. An overview on the treatment of ballast water in ships. Ocean Coast. Manag. 2021, 199, 105296. [Google Scholar] [CrossRef]
- Parsek, M.R.; Fuqua, C. Biofilms 2003: Emerging themes and challenges in studies of surface-associated microbial life. J. Bacteriol. 2004, 186, 4427–4440. [Google Scholar] [CrossRef] [PubMed]
- Procópio, L. The role of biofilms in the corrosion of steel in marine environments. World J. Microbiol. Biotechnol. 2019, 35, 73. [Google Scholar] [CrossRef] [PubMed]
- Ryan, E.; Turkmen, S.; Benson, S. An Investigation into the application and practical use of (UV) ultraviolet light technology for marine antifouling. Ocean Eng. 2020, 216, 107690. [Google Scholar] [CrossRef]
- Hunsucker, K.Z.; Braga, C.; Gardner, H.; Jongerius, M.; Hietbrink, R.; Salters, B.; Swain, G. Using ultraviolet light for improved antifouling performance on ship hull coatings. Biofouling 2019, 35, 658–668. [Google Scholar] [CrossRef] [PubMed]
- Braga, C.; Hunsucker, K.; Gardner, H.; Swain, G. A novel design to investigate the impacts of UV exposure on marine biofouling. Appl. Ocean Res. 2020, 101, 102226. [Google Scholar] [CrossRef]
- Richard, K.N.; Hunsucker, K.Z.; Gardner, H.; Hickman, K.; Swain, G. The Application of UVC Used in Synergy with Surface Material to Prevent Marine Biofouling. J. Mar. Sci. Eng. 2021, 9, 662. [Google Scholar] [CrossRef]
- Gabrielson, J.; Hart, M.; Jarelöv, A.; Kühn, I.; McKenzie, D.; Möllby, R. Evaluation of redox indicators and the use of digital scanners and spectrophotometer for quantification of microbial growth in microplates. J. Microbiol. Methods 2002, 50, 63–73. [Google Scholar] [CrossRef]
- Jiang, W.; Akagi, T.; Suzuki, H.; Takimoto, A.; Nagai, H. A new diatom growth inhibition assay using the XTT colorimetric method. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2016, 185, 13–19. [Google Scholar] [CrossRef]
- Molino, P.J.; Wetherbee, R. The biology of biofouling diatoms and their role in the development of microbial slimes. Biofouling 2008, 24, 365–379. [Google Scholar] [CrossRef]
- Zargiel, K.A.; Coogan, J.S.; Swain, G.W. Diatom community structure on commercially available ship hull coatings. Biofouling 2011, 27, 955–965. [Google Scholar] [CrossRef]
- Lehtola, M.J.; Laxander, M.; Miettinen, I.T.; Hirvonen, A.; Vartiainen, T.; Martikainen, P.J. The effects of changing water flow velocity on the formation of biofilms and water quality in pilot distribution system consisting of copper or polyethylene pipes. Water Res. 2006, 40, 2151–2160. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Pichel, F.; Bebout, B.M. Penetration of ultraviolet radiation into shallow water sediments: High exposure for photosynthetic communities. Mar. Ecol. Prog. Ser. 1996, 131, 257–262. [Google Scholar] [CrossRef]
- Foster, G.; Riegl, B.M.; Foster, K.A.; Morris, L.J. Acoustic detection and mapping of muck deposits in the Indian River Lagoon, Florida. J. Coast. Res. 2018, 34, 856–863. [Google Scholar] [CrossRef]
- Williams, D.E.; Hallock, P. Bleaching in Amphistegina gibbosa d’Orbigny (Class Foraminifera): Observations from laboratory experiments using visible and ultraviolet light. Mar. Biol. 2004, 145, 641–649. [Google Scholar]
- Schmidt, E.C.; Nunes, B.G.; Maraschin, M.; Bouzon, Z.L. Effect of ultraviolet-B radiation on growth, photosynthetic pigments, and cell biology of Kappaphycus alvarezii (Rhodophyta, Gigartinales) macroalgae brown strain. Photosynthetica 2010, 48, 161–172. [Google Scholar] [CrossRef]
- Stuart, V.; Sathyendranath, S.; Platt, T.; Maass, H.; Irwin, B.D. Pigments and species composition of natural phytoplankton populations: Effect on the absorption spectra. J. Plankton Res. 1998, 20, 187–217. [Google Scholar] [CrossRef]
- Characklis, W.G.; Wilderer, P.A. Structure and function of biofilms. In Dahlem Workshop on Structure and Function of Biofilms (1988: Berlin, Germany); Wiley: Hoboken, NJ, USA, 1989. [Google Scholar]
- Stoodley, P.; Boyle, J.D.; DeBeer, D.; Lappin-Scott, H.M. Evolving perspectives of biofilm structure. Biofouling 1999, 14, 75–90. [Google Scholar] [CrossRef]
- Conn, S.A.; Bahena, M.; Davis, J.T.; Ragland, R.L.; Rauschenberg, C.D.; Smith, B.J. Characterisation of the diatom photophobic response to high irradiance. Diatom Res. 2004, 19, 167–179. [Google Scholar] [CrossRef]
- Cohn, S.A.; Spurck, T.P.; Pickett-Heaps, J.D. High energy irradiation at the leading tip of moving diatoms causes a rapid change of cell direction. Diatom Res. 1999, 14, 193–206. [Google Scholar] [CrossRef]
- Martínez, L.F.; Mahamud, M.M.; Lavín, A.G.; Bueno, J.L. The regrowth of phytoplankton cultures after UV disinfection. Mar. Pollut. Bull. 2013, 67, 152–157. [Google Scholar] [CrossRef]
- Martínez, L.F.; Mahamud, M.M.; Lavín, A.G.; Bueno, J.L. Evolution of phytoplankton cultures after ultraviolet light treatment. Mar. Pollut. Bull. 2012, 64, 556–562. [Google Scholar] [CrossRef]
- Schultz, M.P.; Bendick, J.A.; Holm, E.R.; Hertel, W.M. Economic impact of biofouling on a naval surface ship. Biofouling 2011, 27, 87–98. [Google Scholar] [CrossRef] [PubMed]
- Yule, A.B.; Crisp, D.J. Adhesion of cypris larvae of the barnacle, Balanus balanoides, to clean and arthropodin treated surfaces. J. Mar. Biol. Assoc. UK 1983, 63, 261–271. [Google Scholar] [CrossRef]
- Unabia, C.R.C.; Hadfield, M.G. Role of bacteria in larval settlement and metamorphosis of the polychaete Hydroides elegans. Mar. Biol. 1999, 133, 55–64. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| UV Exposure Time | |
|---|---|
| Control | 0 |
| 10 min | 1626.2 |
| 20 min | 3252.4 |
| 30 min | 4878.6 |
| 40 min | 6504.8 |
| 45 min | 7317.9 |
| 60 min | 9757.2 |
| Type | UV Exposure Time | Average XTT Absorbance | Standard Deviation |
|---|---|---|---|
| Field Biofilms | Control | 0.89 | 1.3 × |
| 10 min | 0.54 | 1.1 × | |
| 30 min | 0.91 | 5.5 × | |
| 45 min | 0.60 | 6.5 × | |
| 60 min | 0.42 | 8.1 × | |
| Cultured Biofilms | Control | 0.37 | 0.15 |
| 10 min | 0.04 | 0.03 | |
| 20 min | 0.07 | 0.01 | |
| 30 min | 0.08 | 0.03 | |
| 40 min | 0.06 | 0.03 | |
| 60 min | 0.04 | 0.05 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Braga, C.R.; Richard, K.N.; Gardner, H.; Swain, G.; Hunsucker, K.Z. Investigating the Impacts of UVC Radiation on Natural and Cultured Biofilms: An assessment of Cell Viability. Microorganisms 2023, 11, 1348. https://doi.org/10.3390/microorganisms11051348
Braga CR, Richard KN, Gardner H, Swain G, Hunsucker KZ. Investigating the Impacts of UVC Radiation on Natural and Cultured Biofilms: An assessment of Cell Viability. Microorganisms. 2023; 11(5):1348. https://doi.org/10.3390/microorganisms11051348
Chicago/Turabian StyleBraga, Cierra R., Kailey N. Richard, Harrison Gardner, Geoffrey Swain, and Kelli Z. Hunsucker. 2023. "Investigating the Impacts of UVC Radiation on Natural and Cultured Biofilms: An assessment of Cell Viability" Microorganisms 11, no. 5: 1348. https://doi.org/10.3390/microorganisms11051348
APA StyleBraga, C. R., Richard, K. N., Gardner, H., Swain, G., & Hunsucker, K. Z. (2023). Investigating the Impacts of UVC Radiation on Natural and Cultured Biofilms: An assessment of Cell Viability. Microorganisms, 11(5), 1348. https://doi.org/10.3390/microorganisms11051348