
Molecular Characterization and Prevalence of Antimicrobial-Resistant Escherichia coli Isolates Derived from Clinical Specimens and Environmental Habitats

 and
and 
Abstract
:1. Introduction
2. Materials and Methods
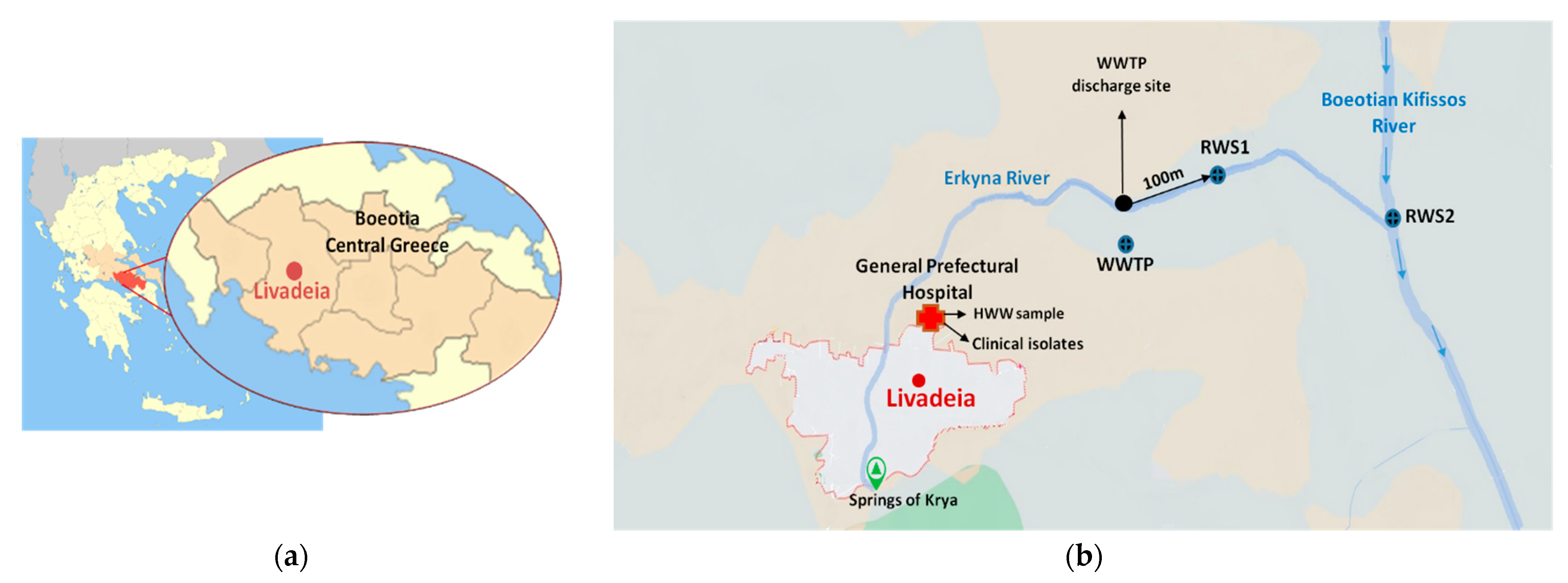
2.1. Sampling Locations and Collected Samples
2.2. E. coli Isolation and Identification
2.3. Antibiotic Susceptibility Testing
2.4. Isolation of Genomic DNA
2.5. PCR Amplification of Resistance Genes
2.6. Molecular Typing
2.6.1. Phylogrouping
2.6.2. Pulsed Field Gel Electrophoresis (PFGE)
2.7. Statistical Analysis
3. Results
3.1. E. coli Collection
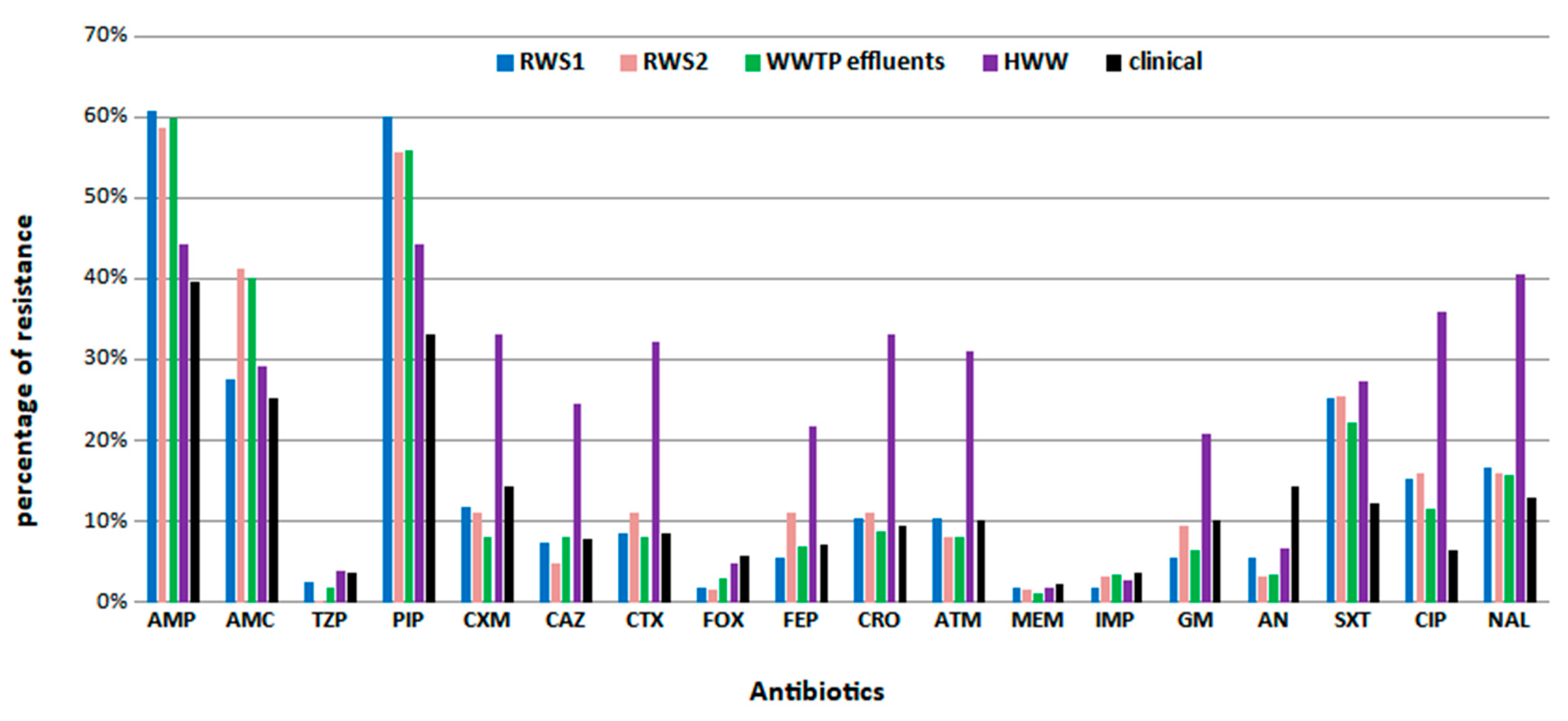
3.2. Antimicrobial Susceptibility Profiles and Assessment of Resistance Mechanisms
3.3. Resistance Genes Detection
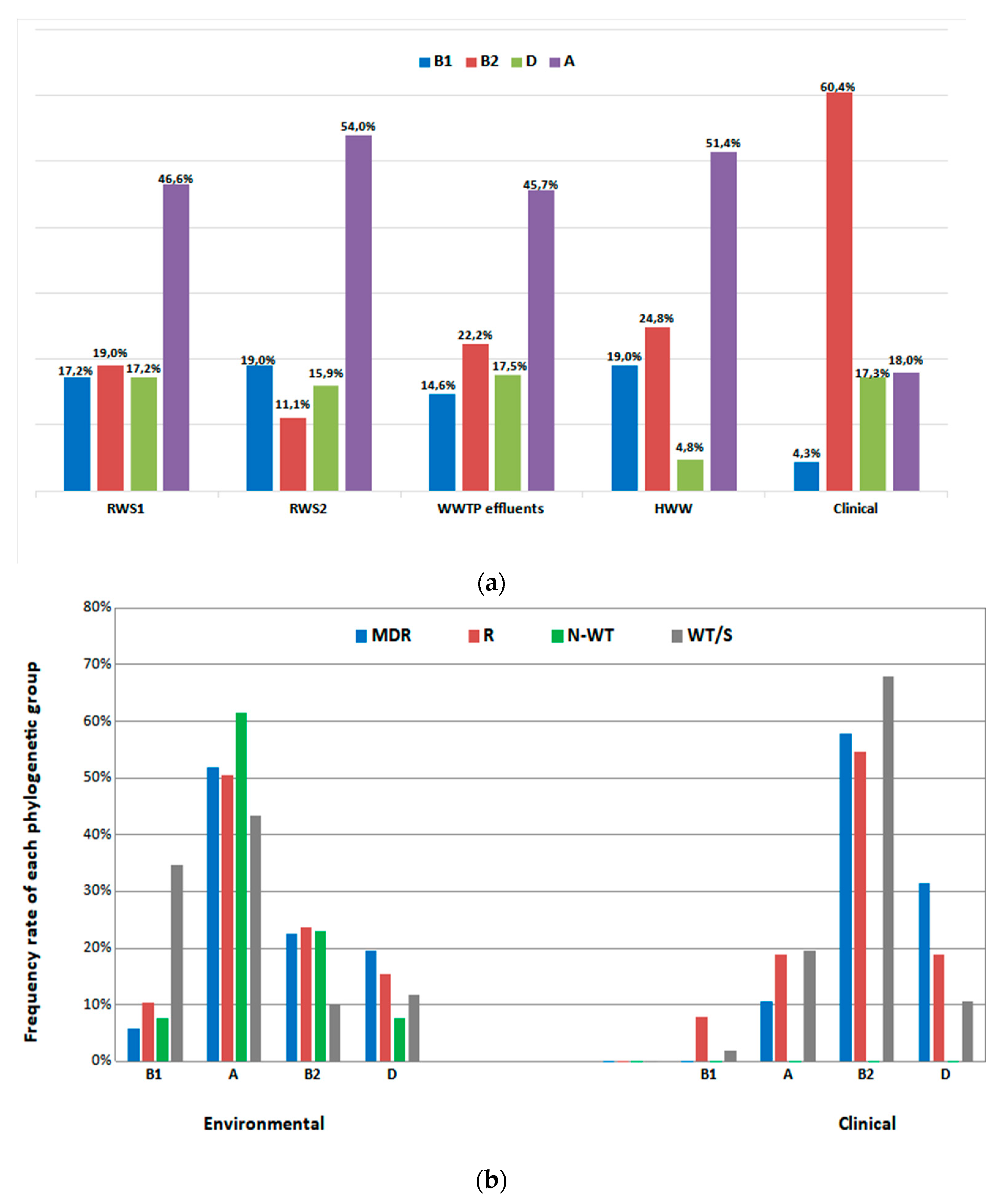
3.4. Molecular Typing Analysis
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kim, S.; Aga, D.S. Potential ecological and human health impacts of antibiotics and antibiotic-resistant bacteria from wastewater treatment plants. J. Toxicol. Environ. Health Part B Crit. Rev. 2007, 10, 559–573. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Antimicrobial Resistance: Global Report on Surveillance; WHO: Geneva, Switzerland, 2014. [Google Scholar]
- Zanotto, C.; Bissa, M.; Illiano, E.; Mezzanotte, V.; Marazzi, F.; Turolla, A.; Antonelli, M.; Morghen, C.D.G.; Radaelli, A. Identification of antibiotic-resistant Escherichia coli isolated from a municipal wastewater treatment plant. Chemosphere 2016, 164, 627–633. [Google Scholar] [CrossRef] [PubMed]
- Ben, W.; Wang, J.; Cao, R.; Yang, M.; Zhang, Y.; Qiang, Z. Distribution of antibiotic resistance in the effluents of ten municipal wastewater treatment plants in China and the effect of treatment processes. Chemosphere 2017, 172, 392–398. [Google Scholar] [CrossRef] [PubMed]
- Aydin, S.; Aydin, M.E.; Ulvi, A.; Kilic, H. Antibiotics in hospital effluents: Occurrence, contribution to urban wastewater, removal in a wastewater treatment plant, and environmental risk assessment. Environ. Sci. Pollut. Res. Int. 2019, 26, 544–558. [Google Scholar] [CrossRef]
- Cahill, N.; O’Connor, L.; Mahon, B.; Varley, Á.; McGrath, E.; Ryan, P.; Cormican, M.; Brehony, C.; Jolley, K.A.; Maiden, M.C.; et al. Hospital effluent: A reservoir for carbapenemase-producing Enterobacterales? Sci. Tota. Environ. 2019, 672, 618–624. [Google Scholar] [CrossRef]
- Amarasiri, M.; Sano, D.; Suzuki, S. Understanding human health risks caused by antibiotic resistant bacteria (ARB) and antibiotic resistance genes (ARG) in water environments: Current knowledge and questions to be answered. Crit. Rev. Environ. Sci. Technol. 2020, 50, 2016–2059. [Google Scholar] [CrossRef]
- Watkinson, A.J.; Micalizzi, G.B.; Graham, G.M.; Bates, J.B.; Costanzo, S.D. Antibiotic-resistant Escherichia coli in wastewaters, surface waters, and oysters from an urban riverine system. Appl. Environ. Microbiol. 2007, 73, 5667–5670. [Google Scholar] [CrossRef]
- Jang, J.; Hur, H.-G.; Sadowsky, M.; Byappanahalli, M.; Yan, T.; Ishii, S. Environmental Escherichia coli: Ecology and public health implications—A review. J. Appl. Microbiol. 2017, 123, 570–581. [Google Scholar] [CrossRef]
- Haberecht, H.B.; Nealon, N.J.; Gilliland, J.R.; Holder, A.V.; Runyan, C.; Oppel, R.C.; Ibrahim, H.M.; Mueller, L.; Schrupp, F.; Vilchez, S.; et al. Antimicrobial-Resistant Escherichia coli from Environmental Waters in Northern Colorado. J. Environ. Public Health 2019, 2019, 3862949. [Google Scholar] [CrossRef]
- Fewtrell, L.; Bartram, J. Water Quality: Guidelines, Standards and Health: Assessment of Risk and Risk Management for Water-Related Infectious Diseases/Edited by Lorna Fewtrell and Jamie Bartram; World Health Organization: Geneva, Switzerland, 2001; Available online: https://apps.who.int/iris/handle/10665/42442 (accessed on 11 June 2022).
- Zhang, C.; Xu, L.; Wang, X.; Zhuang, K.; Liu, Q. Effects of ultraviolet disinfection on antibiotic-resistant Escherichia coli from wastewater: Inactivation, antibiotic resistance profiles and antibiotic resistance genes. J. Appl. Microbiol. 2017, 123, 295–306. [Google Scholar] [CrossRef]
- Kaper, J.B.; Nataro, J.P.; Mobley, H.L.T. Pathogenic Escherichia coli. Nat. Rev. Microbiol. 2004, 2, 123–140. [Google Scholar] [CrossRef]
- Carducci, A.; Verani, M. Effects of bacterial, chemical, physical and meteorological variables on virus removal by a wastewater treatment plant. Food Environ. Virol. 2013, 5, 69–76. [Google Scholar] [CrossRef]
- Kokkinos, P.; Mandilara, G.; Nikolaidou, A.; Velegraki, A.; Theodoratos, P.; Kampa, D.; Blougoura, A.; Christopoulou, A.; Smeti, E.; Kamizoulis, G.; et al. Performance of three small-scale wastewater treatment plants. A challenge for possible re use. Environ. Sci. Pollut. Res. Int. 2015, 22, 17744–17752. [Google Scholar] [CrossRef]
- Hong, P.-Y.; Julian, T.R.; Pype, M.-L.; Jiang, S.C.; Nelson, K.L.; Graham, D.; Pruden, A.; Manaia, C.M. Reusing Treated Wastewater: Consideration of the Safety Aspects Associated with Antibiotic-Resistant Bacteria and Antibiotic Resistance Genes. Water 2018, 10, 244. [Google Scholar] [CrossRef]
- Carducci, A.; Donzelli, G.; Cioni, L.; Federigi, I.; Lombardi, R.; Verani, M. Quantitative Microbial Risk Assessment for Workers Exposed to Bioaerosol in Wastewater Treatment Plants Aimed at the Choice and Setup of Safety Measures. Int. J. Environ. Res. Public Health 2018, 15, 1490. [Google Scholar] [CrossRef]
- Hossain, S.; Ali, S.; Hossain, M.; Uddin, S.Z.; Moniruzzaman, M.; Islam, M.R.; Shohael, A.M.; Islam, S.; Ananya, T.H.; Rahman, M.; et al. ESBL Producing Escherichia coli in Faecal Sludge Treatment Plants: An Invisible Threat to Public Health in Rohingya Camps, Cox’s Bazar, Bangladesh. Front. Public Health 2021, 9, 783019. [Google Scholar] [CrossRef]
- Aslan, A.; Cole, Z.; Bhattacharya, A.; Oyibo, O. Presence of Antibiotic-Resistant Escherichia coli in Wastewater Treatment Plant Effluents Utilized as Water Reuse for Irrigation. Water 2018, 10, 805. [Google Scholar] [CrossRef]
- Kyriakou, G.; Nikolaou, A.; Chrysafopoulou, E. Interventions for the Qualitative and Quantitative Water Resources Management of the Kifissos River Basin; Technical Chamber of Greece Division of Eastern Sterea Ellada: Athens, Greece, 2009. [Google Scholar]
- ISO 9308.01-1: 2017; Water Quality—Enumeration of Escherichia coli and Coliform Bacteria—Part 1: Membrane Filtration Method for Waters with Low Bacterial Background Flora. International Organization for Standardization: Geneva, Switzerland, 2017.
- Bej, A.K.; McCarty, S.C.; Atlas, R.M. Detection of coliform bacteria and Escherichia coli by multiplex polymerase chain reaction: Comparison with defined substrate and plating methods for water quality monitoring. Appl. Environ. Microbiol. 1991, 57, 2429–2432. [Google Scholar] [CrossRef]
- World Health Organization. Guidance on Regulations for the Transport of Infectious Substances 2009–2010: Applicable as from 1 January 2009; World Health Organization: Geneva, Switzerland, 2009; Available online: https://apps.who.int/iris/handle/10665/69899 (accessed on 11 June 2022).
- European Committee on Antimicrobial Susceptibility Testing. EUCAST ECOFFs and Breakpoint Tables for Interpretation of MICs and Zone Diameters. Version 12.0. 2022. Available online: http://www.eucast.org (accessed on 10 June 2020).
- Magiorakos, A.-P.; Srinivasan, A.; Carey, R.B.; Carmeli, Y.; Falagas, M.E.; Giske, C.G.; Harbarth, S.; Hindler, J.F.; Kahlmeter, G.; Olsson-Liljequist, B.; et al. Multidrug-resistant, extensively drug-resistant and pandrug-resistant bacteria: An international expert proposal for interim standard definitions for acquired resistance. Clin. Microbiol. Infect. 2012, 18, 268–281. [Google Scholar] [CrossRef]
- Olga, P.; Apostolos, V.; Alexis, G.; George, V.; Athena, M. Antibiotic resistance profiles of Pseudomonas aeruginosa isolated from various Greek aquatic environments. FEMS Microbiol. Ecol. 2016, 92, fiw042. [Google Scholar] [CrossRef]
- European Committee on Antimicrobial Susceptibility Testing. EUCAST Guidelines for Detection of Resistance Mechanisms and Specific Resistances of Clinical and/or Epidemiological Importance. Version 2.0. 2017. Available online: https://www.eucast.org/fileadmin/src/media/PDFs/EUCAST_files/Resistance_mechanisms/EUCAST_detection_of_resistance_mechanisms_170711.pdf (accessed on 10 June 2020).
- Van Der Zwaluw, K.; De Haan, A.; Pluister, G.N.; Bootsma, H.J.; De Neeling, A.J.; Schouls, L.M. The Carbapenem inactivation method (CIM), a simple and low-cost alternative for the Carba Np test to assess phenotypic carbapenemase activity in gram-negative rods. PLoS ONE 2015, 10, e0123690. [Google Scholar] [CrossRef] [PubMed]
- Poirel, L.; Héritier, C.; Tolun, V.; Nordmann, P. Emergence of oxacillinase-mediated resistance to imipenem in Klebsiella pneumoniae. Antimicrob. Agents Chemother. 2004, 48, 15–22. [Google Scholar] [CrossRef] [PubMed]
- Woodford, N.; Fagan, E.J.; Ellington, M.J. Multiplex PCR for rapid detection of genes encoding CTX-M extended-spectrum (beta)-lactamases. J. Antimicrob. Chemother. 2006, 57, 154–155. [Google Scholar] [CrossRef] [PubMed]
- European Centre for Disease Prevention and Control. Laboratory Manual for Carbapenem and Colistin Resistance Detection and Characterisation for the Survey of Carbapenem- and/or Colistin-Resistant Enterobacteriaceae—Version 2.0; ECDC: Stockholm, Sweden, 2019. [Google Scholar]
- Pérez-Pérez, F.J.; Hanson, N.D. Detection of plasmid-mediated AmpC beta-lactamase genes in clinical isolates by using multiplex PCR. J. Clin. Microbiol. 2002, 40, 2153–2162. [Google Scholar] [CrossRef] [PubMed]
- Hannecart-Pokorni, E.; Depuydt, F.; de Wit, L.; van Bossuyt, E.; Content, J.; Vanhoof, R. Characterization of the 6’-N-aminoglycoside acetyltransferase gene aac(6′)-Im [corrected] associated with a sulI-type integron. Antimicrob. Agents Chemother. 1997, 41, 314–318. [Google Scholar] [CrossRef]
- Pappa, O.; Beloukas, A.; Vantarakis, A.; Mavridou, A.; Kefala, A.-M.; Galanis, A. Molecular Characterization and Phylogenetic Analysis of Pseudomonas aeruginosa Isolates Recovered from Greek Aquatic Habitats Implementing the Double-Locus Sequence Typing Scheme. Microb. Ecol. 2017, 74, 78–88. [Google Scholar] [CrossRef]
- Clermont, O.; Bonacorsi, S.; Bingen, E. Rapid and simple determination of the Escherichia coli phylogenetic group. Appl. Environ. Microbiol. 2000, 66, 4555–4558. [Google Scholar] [CrossRef]
- Standard Operating Procedure for PulseNet PFGE of Escherichia coli O157:H7, Escherichia coli Non O157 (STEC), Salmonella serotypes, Shigella sonnei and Shigella flexneri. April 2013. Available online: https://www.cdc.gov/pulsenet/pdf/ecoli-shigella-salmonella-pfge-protocol-508c.pdf (accessed on 10 June 2020).
- Tenover, F.C.; Arbeit, R.D.; Goering, R.V.; A Mickelsen, P.; E Murray, B.; Persing, D.H.; Swaminathan, B. Interpreting chromosomal DNA restriction patterns produced by pulsed-field gel electrophoresis: Criteria for bacterial strain typing. J. Clin. Microbiol. 1995, 33, 2233–2239. [Google Scholar] [CrossRef]
- Suzuki, Y.; Hashimoto, R.; Xie, H.; Nishimura, E.; Nishiyama, M.; Nukazawa, K.; Ishii, S. Growth and antibiotic resistance acquisition of Escherichia coli in a river that receives treated sewage effluent. Sci. Total Environ. 2019, 690, 696–704. [Google Scholar] [CrossRef]
- Reinthaler, F.; Posch, J.; Feierl, G.; Wüst, G.; Haas, D.; Ruckenbauer, G.; Mascher, F.; Marth, E. Antibiotic resistance of E. coli in sewage and sludge. Water Res. 2003, 37, 1685–1690. [Google Scholar] [CrossRef]
- Hassan, S.A.; Jamal, S.A.; Kamal, M. Occurrence of multidrug resistant and ESBL producing E. coli causing urinary tract infections. Aust. J. Basic Appl. Sci. 2011, 7, 39–43. [Google Scholar]
- Hu, Y.-Y.; Cai, J.-C.; Zhou, H.-W.; Chi, D.; Zhang, X.-F.; Chen, W.-L.; Zhang, R.; Chen, G.-X. Molecular typing of CTX-M-producing escherichia coli isolates from environmental water, swine feces, specimens from healthy humans, and human patients. Appl. Environ. Microbiol. 2013, 79, 5988–5996. [Google Scholar] [CrossRef]
- Saltoglu, N.; Karali, R.; Yemisen, M.; Ozaras, R.; Balkan, I.I.; Mete, B.; Tabak, F.; Mert, A.; Hondur, N.; Ozturk, R. Comparison of community-onset healthcare-associated and hospital-acquired urinary infections caused by extended-spectrum beta-lactamase-producing Escherichia coli and antimicrobial activities. Int. J. Clin. Pract. 2015, 69, 766–770. [Google Scholar] [CrossRef]
- Stoll, C.; Sidhu, J.P.S.; Tiehm, A.; Toze, S. Prevalence of clinically relevant antibiotic resistance genes in surface water samples collected from Germany and Australia. Environ. Sci. Technol. 2012, 46, 9716–9726. [Google Scholar] [CrossRef]
- Liu, H.; Zhou, H.; Li, Q.; Peng, Q.; Zhao, Q.; Wang, J.; Liu, X. Molecular characteristics of extended-spectrum β-lactamase-producing Escherichia coli isolated from the rivers and lakes in Northwest China. BMC Microbiol. 2018, 18, 125. [Google Scholar] [CrossRef]
- Lee, J.H.; Subhadra, B.; Son, Y.; Kim, D.H.; Park, H.S.; Kim, J.M.; Koo, S.H.; Oh, M.H.; Kim, H.; Choi, C.H. Phylogenetic group distributions, virulence factors and antimicrobial resistance properties of uropathogenic Escherichia coli strains isolated from patients with urinary tract infections in South Korea. Lett. Appl. Microbiol. 2016, 62, 84–90. [Google Scholar] [CrossRef]
- Iranpour, D.; Hassanpour, M.; Ansari, H.; Tajbakhsh, S.; Khamisipour, G.; Najafi, A. Phylogenetic groups of escherichia coli strains from patients with urinary tract infection in Iran based on the new Clermont phylotyping method. BioMed Res. Int. 2015, 2015, 846219. [Google Scholar] [CrossRef]
- Dadi, B.R.; Abebe, T.; Zhang, L.; Mihret, A.; Abebe, W.; Amogne, W. Distribution of virulence genes and phylogenetics of uropathogenic Escherichia coli among urinary tract infection patients in Addis Ababa, Ethiopia. BMC Infect. Dis. 2020, 20, 108. [Google Scholar] [CrossRef]
- Spurbeck, R.R.; Dinh, P.C., Jr.; Walk, S.T.; Stapleton, A.E.; Hooton, T.M.; Nolan, L.K.; Kim, K.S.; Johnson, J.R.; Mobley, H.L.T. Escherichia coli isolates that carry vat, fyuA, chuA, and yfcV efficiently colonize the urinary tract. Infect. Immun. 2012, 80, 4115–4122. [Google Scholar] [CrossRef]
- Stoppe, N.D.C.; Silva, J.S.; Carlos, C.; Sato, M.I.Z.; Saraiva, A.M.; Ottoboni, L.M.M.; Torres, T.T. Worldwide Phylogenetic Group Patterns of Escherichia coli from Commensal Human and Wastewater Treatment Plant Isolates. Front. Microbiol. 2017, 8, 2512. [Google Scholar] [CrossRef]
- Nagy, G.; Dobrindt, U.; Kupfer, M.; Emody, L.; Karch, H.; Hacker, J. Expression of hemin receptor molecule ChuA is influenced by RfaH in uropathogenic Escherichia coli strain 536. Infect. Immun. 2001, 69, 1924–1928. [Google Scholar] [CrossRef] [PubMed]
- Mbanga, J.; Abia, A.L.K.; Amoako, D.G.; Essack, S.Y. Quantitative microbial risk assessment for waterborne pathogens in a wastewater treatment plant and its receiving surface water body. BMC Microbiol. 2020, 20, 346. [Google Scholar] [CrossRef] [PubMed]
- Sano, D.; Haas, C.N.; Rose, J.B. A QMRA Framework for Sanitation Treatment Decisions. In Water and Sanitation for the 21st Century: Health and Microbiological Aspects of Excreta and Wastewater Management (Global Water Pathogen Project), Part 1: The Health Hazards of Excreta: Theory and Control; Rose, J.B., Jiménez-Cisneros, B., Eds.; Michigan State University: East Lansing, MI, USA, 2019. [Google Scholar]
- Singh, G.; Vajpayee, P.; Rani, N.; Amoah, I.D.; Stenström, T.A.; Shanker, R. Exploring the potential reservoirs of non specific TEM beta lactamase (bla TEM) gene in the Indo-Gangetic region: A risk assessment approach to predict health hazards. J. Hazard. Mater. 2016, 314, 121–128. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Environmental Isolates | WT | N-WT | R | MDR |
|---|---|---|---|---|
| HWW (n = 105) | 54.3% (57/105) | 3.8% (4/105) | 7.6% (8/105) | 34.3% (36/105) |
| WWTP (n = 171) | 36.8% (63/171) | 3.5% (6/171) | 43.3% (74/171) | 16.4% (28/171) |
| RWS1 (n = 163) | 36.8% (60/163) | 1.2% (2/163) | 45.4% (74/163) | 16.6% (27/163) |
| RWS2 (n = 63) | 36.5% (23/63) | 3.1% (2/63) | 42.9% (27/63) | 17.5% (11/63) |
| Clinical Isolates | S | R | MDR | |
| urine (n = 104) | 41.3% (43/104) | 45.2% (47/104) | 13.5% (14/104) | |
| blood (n = 30) | 43.3% (13/30) | 43.3% (13/30) | 13.4% (4/30) | |
| tissue (n = 5) | - | 80% (4/5) | 20% (1/5) | |
| Environmental Isolates (Source) | Clinical Isolates | |||
|---|---|---|---|---|
| MRP1: Related to ESBL production and resistance to penicillin/inhibitor combinations | PEN/PEN–inhibitor/ESCs + SXT | 1 (WWTP) | - | |
| PEN/PEN-inhibitor/ESCs + QNs | 1 (HWW) | - | ||
| PEN/PEN-inhibitor/ESCs/ATM | 2 (1 RWS1, 1 RWS2) | - | ||
| PEN/PEN-inhibitor/ESCs/ATM + QNs | 9 (3 HWW, 3 WWTP, 3 RWS1) | - | ||
| PEN/PEN-inhibitor/ESCs/ATM + AMG | 2 (1 WWTP, 1 RWS2) | - | ||
| PEN/PEN-inhibitor/ESCs/ATM + SXT | - | 1 | ||
| PEN/PEN-inhibitor/ESCs/ATM + SXT + QNs | 2 (1 HWW + 1 RWS2) | 2 | ||
| PEN/PEN-inhibitor/ESCs/ATM + AMG + QNs | 2 (HWW) | 3 | ||
| PEN/PEN-inhibitor/ESCs/ATM + AMG + SXT + QNs | 11 (HWW) | 1 | ||
| Total MRP1: 37 | 30 | 7 | ||
| MRP2: Related to ESBL production | PEN/ESCs/ATM + QNs | 8 (4 HWW, 3 RWS1, 1 RWS2) | - | |
| PEN/ESCs/ATM + AMG | 3 (2 RWS1, 1 RWS2) | - | ||
| PEN/ESCs/ATM + SXT | 7 (6 WWTP, 1 RWS1) | - | ||
| PEN/ESCs/ATM + SXT + QNs | 2 (WWTP) | 1 | ||
| PEN/ESCs/ATM + AMG + SXT + QNs | 8 (6 HWW, 2 RWS1) | - | ||
| PEN/ESCs/ATM + AMG + QNs | 4 (HWW) | - | ||
| PEN/ESCs + SXT + QNs | 1 (WWTP) | - | ||
| Total MRP2: 34 | 33 | 1 | ||
| MRP 3: Related to ESBL + carbapenemase production | PEN/PEN-inhibitor/ESCs/CARB/ATM + SXT | 1 (RWS1) | - | |
| Total MRP3: 1 | 1 | - | ||
| MRP 4: Related to ESBL + AmpC β-lactamases production | PEN/PEN-inhibitor/ESCs/FOX/ATM | 2 (1 HWW, 1 RWS1) | 1 | |
| PEN/PEN-inhibitor/ESCs/FOX/ATM + AMG + SXT | - | 1 | ||
| PEN/PEN-inhibitor/ESCs/FOX/ATM + SXT + QNs | - | 1 | ||
| PEN/PEN-inhibitor/ESCs/FOX + AMG + SXT + QNs | 2 (HWW) | - | ||
| Total MRP 4: 7 | 4 | 3 | ||
| MRP 5: Related to AmpC β-lactamases production | PEN/PEN-inhibitor/FOX + AMG + QNs | 1 (WWTP) | - | |
| PEN/PEN-inhibitor/FOX + QNs | 1 (WWTP) | - | ||
| PEN/PEN-inhibitor/NSCs/FOX + AMG | 2 (RWS1) | 2 | ||
| Total MRP5: 6 | 4 | 2 | ||
| MPR 6 | Susceptibility to cephalosporins Penicillinase production with resistance to other non-β-lactam antibiotics | PEN/PEN-inhibitor + SXT + QNs | 9 (2 WWTP, 4 RWS1, 3 RWS2) | 1 |
| Total MRP6: 10 | 9 | 1 | ||
| MRP 7 | PEN/PEN-inhibitor + AMG + SXT | 10 (6 WWTP, 4 RWS1) | 5 | |
| Total MRP7: 13 | 10 | 3 | ||
| MRP 8 | PEN/PEN-inhibitor + AMG + SXT + QNs | 7 (1 HWW, 1 WWTP, 2 RWS1, 3 RWS2) | 2 | |
| Total MRP8: 9 | 7 | 2 | ||
| MRP 9 | PEN/PEN-inhibitor + AMG + QNs | 1 (WWTP) | - | |
| Total MRP9: 1 | 1 | - | ||
| MRP10 | PEN + AMG + SXT + QNs | 3 (2 WWTP, 1 RWS1) | - | |
| Total MRP10: 3 | 3 | - | ||
| β-Lactamase Genes | Clinical Isolates | Environmental Isolates | Total | ||||
|---|---|---|---|---|---|---|---|
| HWW | WWTP Effluents | RWS1 | RWS2 | ||||
| ESBL genes | blaCTX-M-group-1-type | 10 | 22 | 10 | 8 | 2 | 52 |
| blaCTX-M-group-9-type | 1 | 3 | 3 | 7 | |||
| blaSHV | 1 | 12 | 2 | 3 | 17 | ||
| blaTEM | 4 | 3 | 2 | 3 | 12 | ||
| Carbapenemase genes | blaOXA-48-type | 1 | 1 | ||||
| AmpC-type genes | blaCMY-2-type | 1 | 1 | ||||
| blaFOX-type | 1 | 1 | |||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dioli, C.; Pappa, O.; Siatravani, E.; Bratakou, S.; Tatsiopoulos, A.; Giakkoupi, P.; Miriagou, V.; Beloukas, A. Molecular Characterization and Prevalence of Antimicrobial-Resistant Escherichia coli Isolates Derived from Clinical Specimens and Environmental Habitats. Microorganisms 2023, 11, 1399. https://doi.org/10.3390/microorganisms11061399
Dioli C, Pappa O, Siatravani E, Bratakou S, Tatsiopoulos A, Giakkoupi P, Miriagou V, Beloukas A. Molecular Characterization and Prevalence of Antimicrobial-Resistant Escherichia coli Isolates Derived from Clinical Specimens and Environmental Habitats. Microorganisms. 2023; 11(6):1399. https://doi.org/10.3390/microorganisms11061399
Chicago/Turabian StyleDioli, Chrysoula, Olga Pappa, Eirini Siatravani, Spyridoula Bratakou, Apostolos Tatsiopoulos, Panagiota Giakkoupi, Vivi Miriagou, and Apostolos Beloukas. 2023. "Molecular Characterization and Prevalence of Antimicrobial-Resistant Escherichia coli Isolates Derived from Clinical Specimens and Environmental Habitats" Microorganisms 11, no. 6: 1399. https://doi.org/10.3390/microorganisms11061399
APA StyleDioli, C., Pappa, O., Siatravani, E., Bratakou, S., Tatsiopoulos, A., Giakkoupi, P., Miriagou, V., & Beloukas, A. (2023). Molecular Characterization and Prevalence of Antimicrobial-Resistant Escherichia coli Isolates Derived from Clinical Specimens and Environmental Habitats. Microorganisms, 11(6), 1399. https://doi.org/10.3390/microorganisms11061399