
Effect of Low Doses of Dexamethasone on Experimental Pulmonary Tuberculosis

 ,
, 
 ,
,  ,
,
Abstract
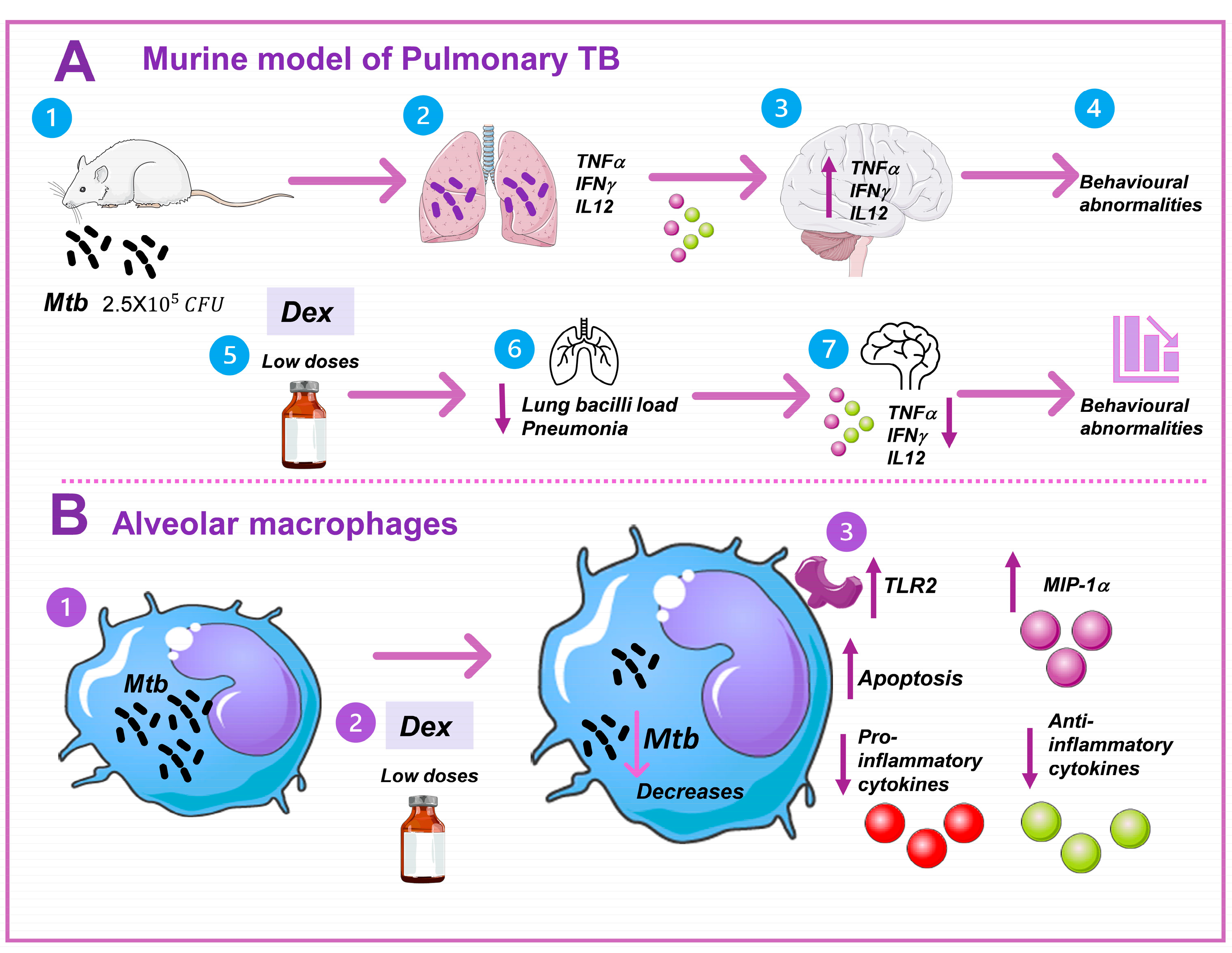
:1. Introduction
2. Materials and Methods
2.1. Reagents and Materials
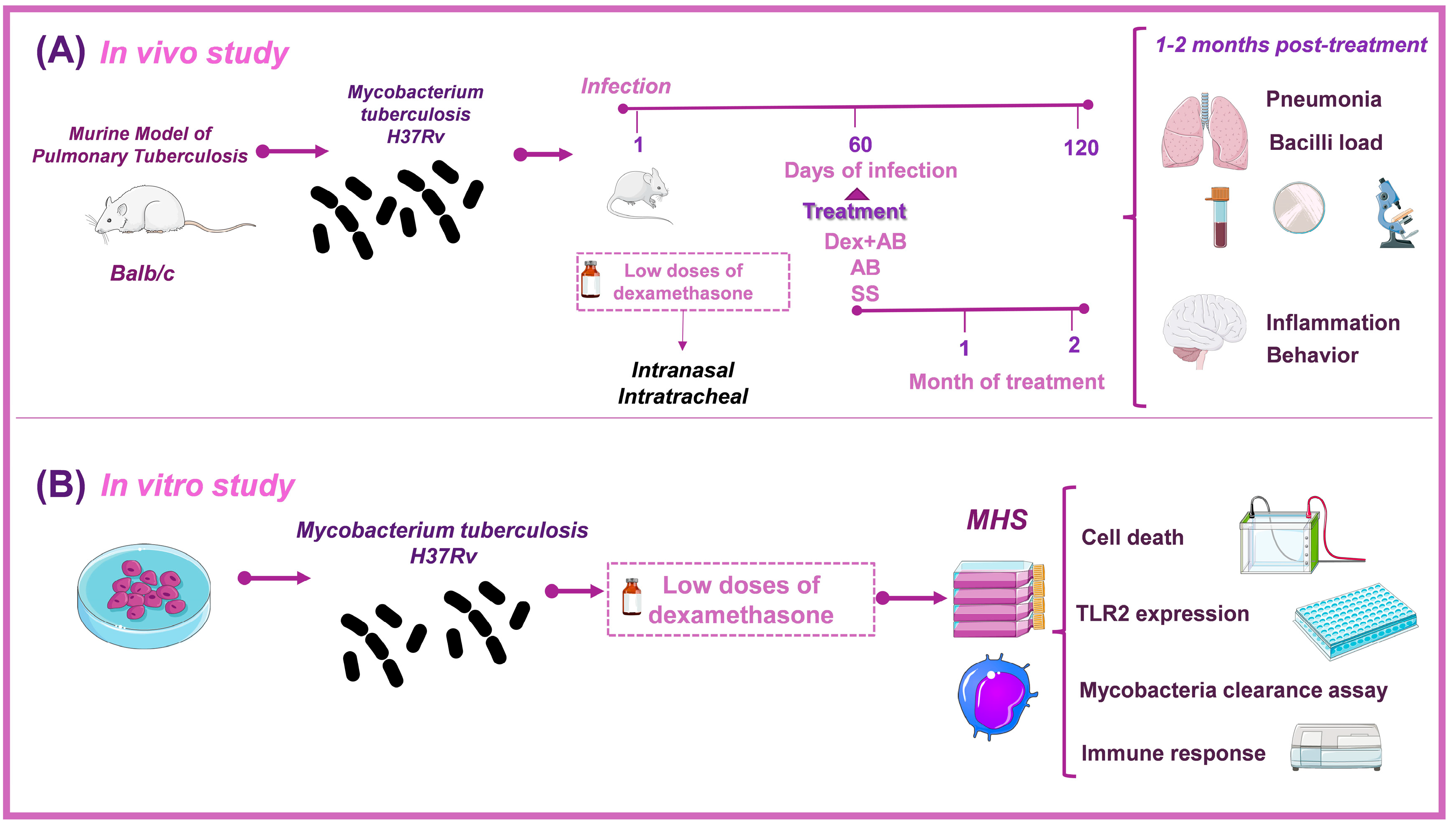
2.2. Experimental Design
2.3. Animals
2.4. Murine Model of Pulmonary Tuberculosis
2.5. Treatments
2.6. Colony-Forming Units (CFU) Methodology for Determining Brain and Pulmonary Bacillary Loads
2.7. Preparation of Tissue for Histology and Morphometry
2.8. RT-PCR Analysis of TNF and IL12 Expression in Several Brain Areas
2.9. Behavioral Studies
2.9.1. Sickness Behavior
2.9.2. Depression-like Behavior
2.9.3. Anxiety-like Behavior
2.9.4. Neurological Outcomes in Animals
2.9.5. Memory Impairment
2.10. Cell and Mycobacteria Culture
2.11. Infection of Macrophages and Bacillary Load Assay (CFU)
2.12. TLR-2 Expression of Macrophages by RT-PCR
2.13. Fragmentation Analysis of DNA
2.14. Evaluation of the Time-Dependent Viability of Cells
2.15. Quantification of Secreted Cytokines
2.16. Statistical Evaluation
3. Results
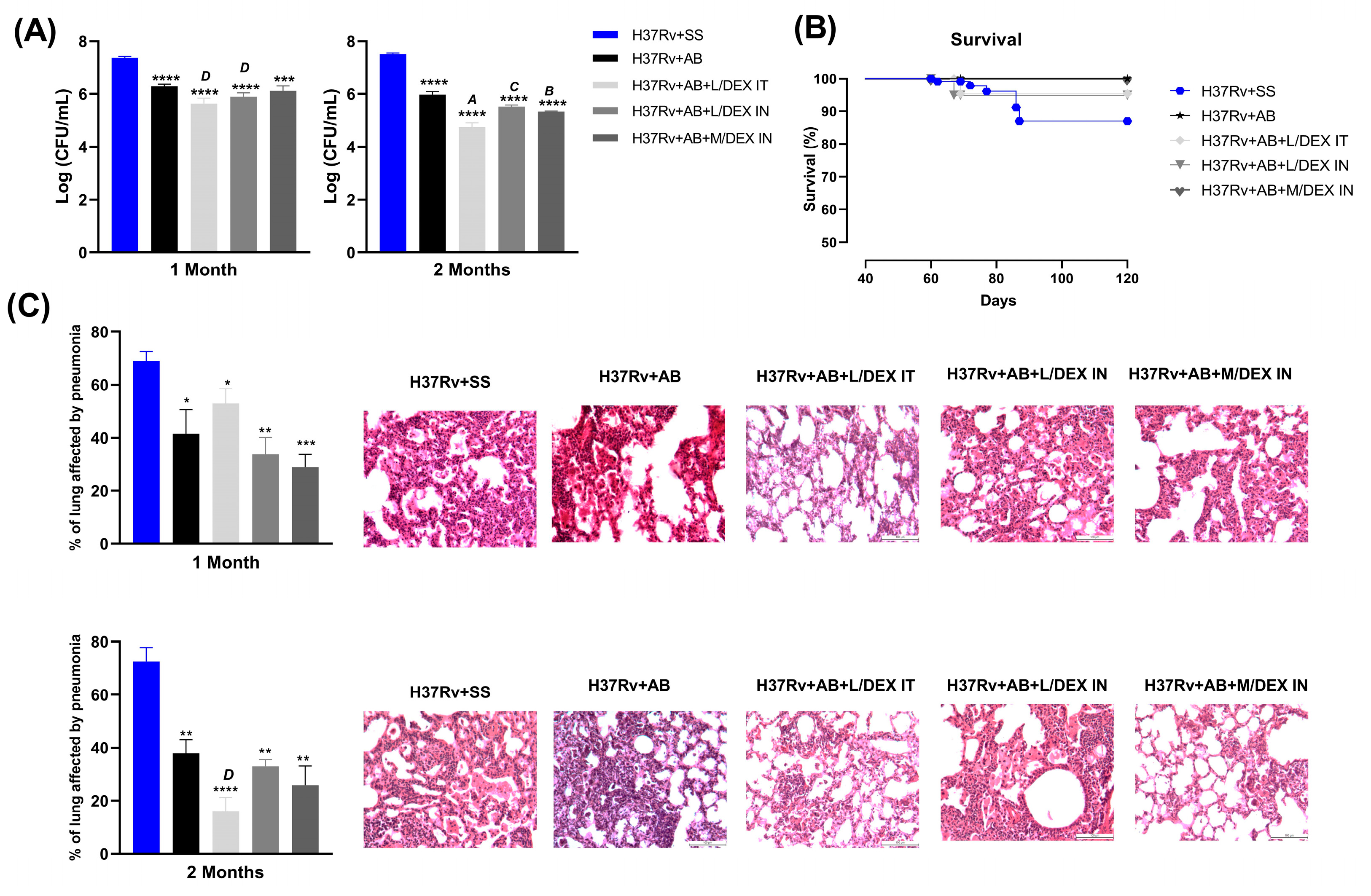
3.1. Low Doses of Dexamethasone in Combination with Antibiotics Decreased the Lung Bacilli Load and Tissue Damage (Pneumonia) of Mice Chronically Infected with Mtb H37Rv
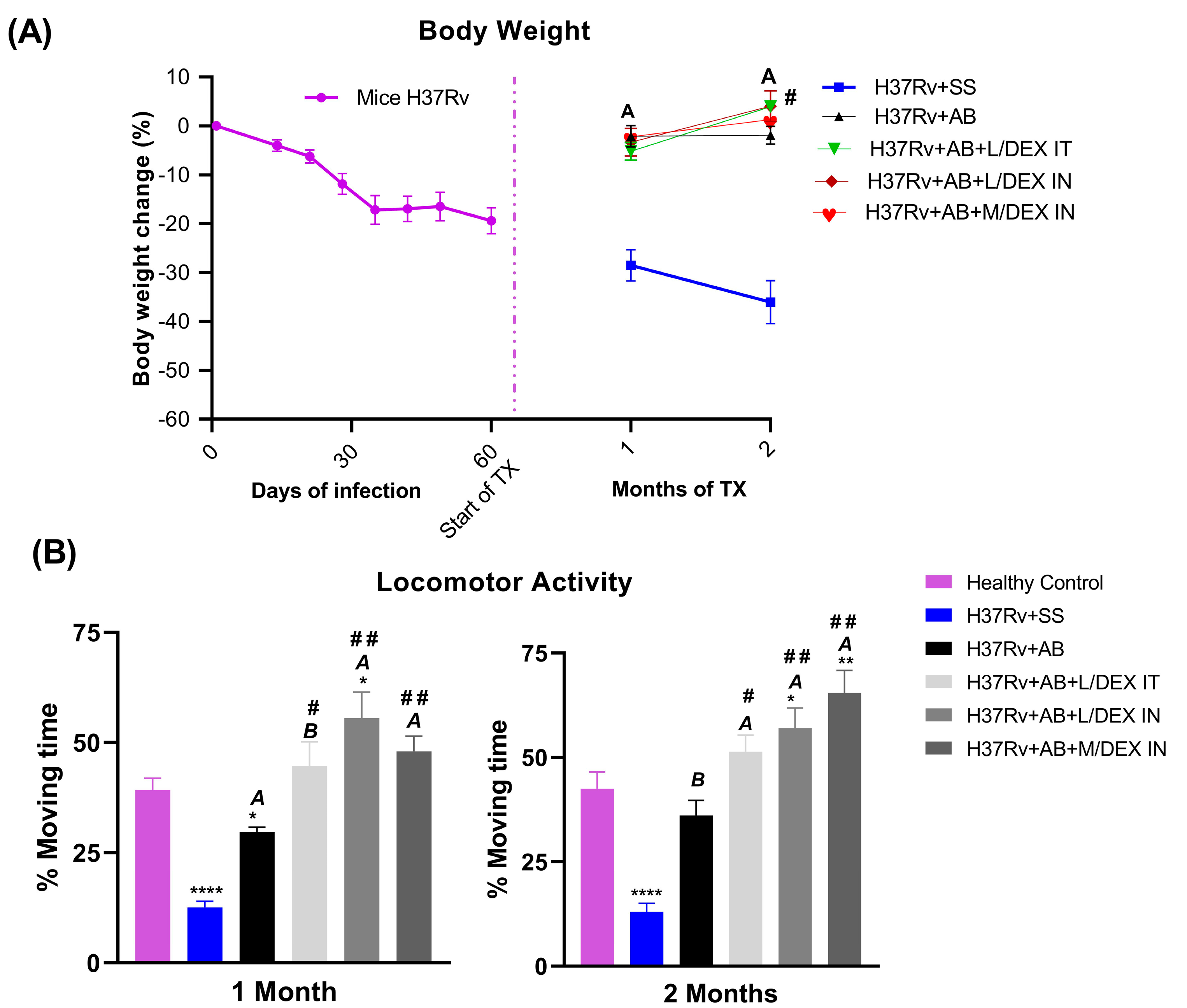
3.2. Low Doses of Dexamethasone in Combination with Antibiotics Decreased the Sickness Behavior of Mice Chronically Infected with Mtb H37Rv
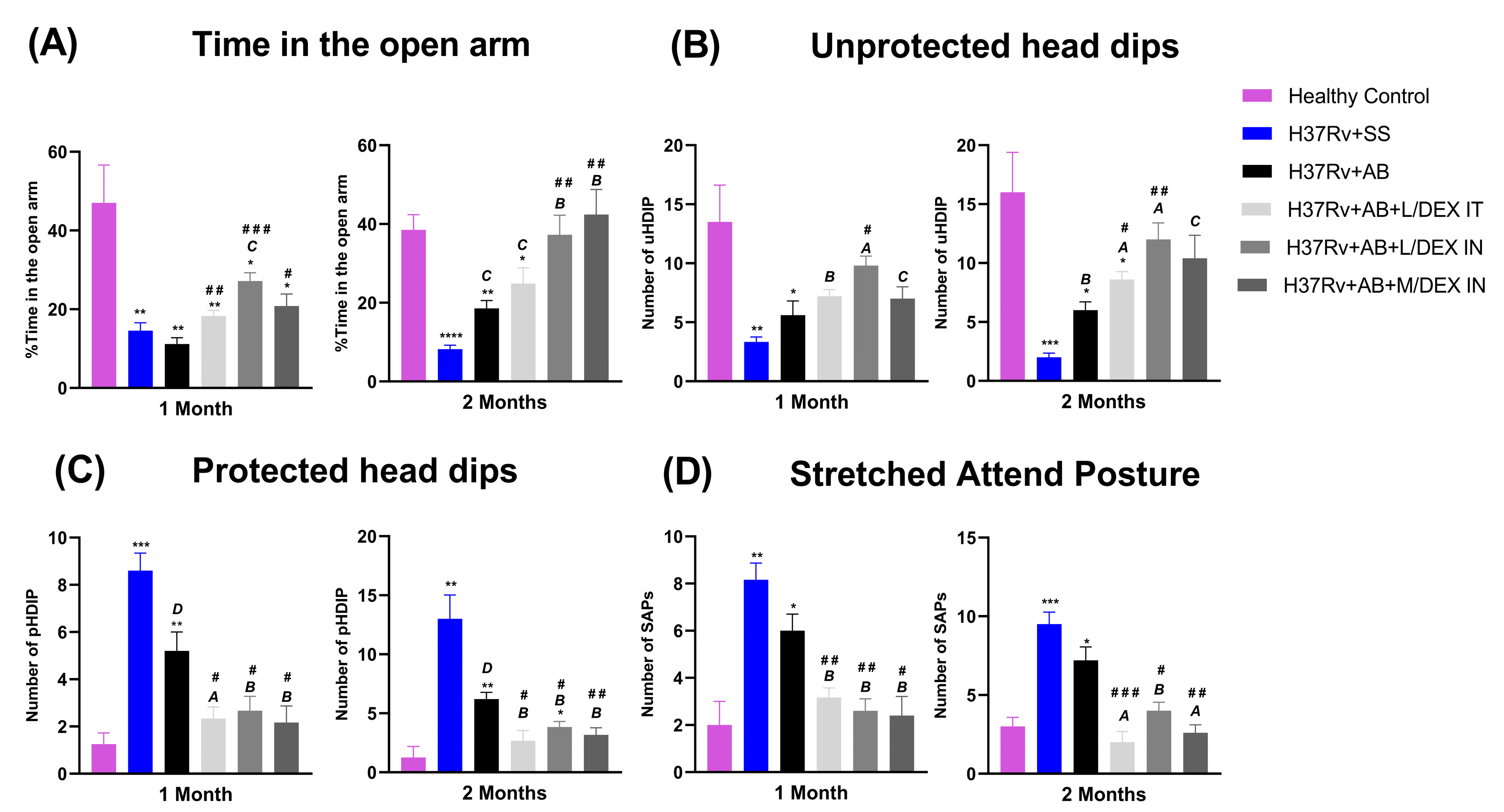
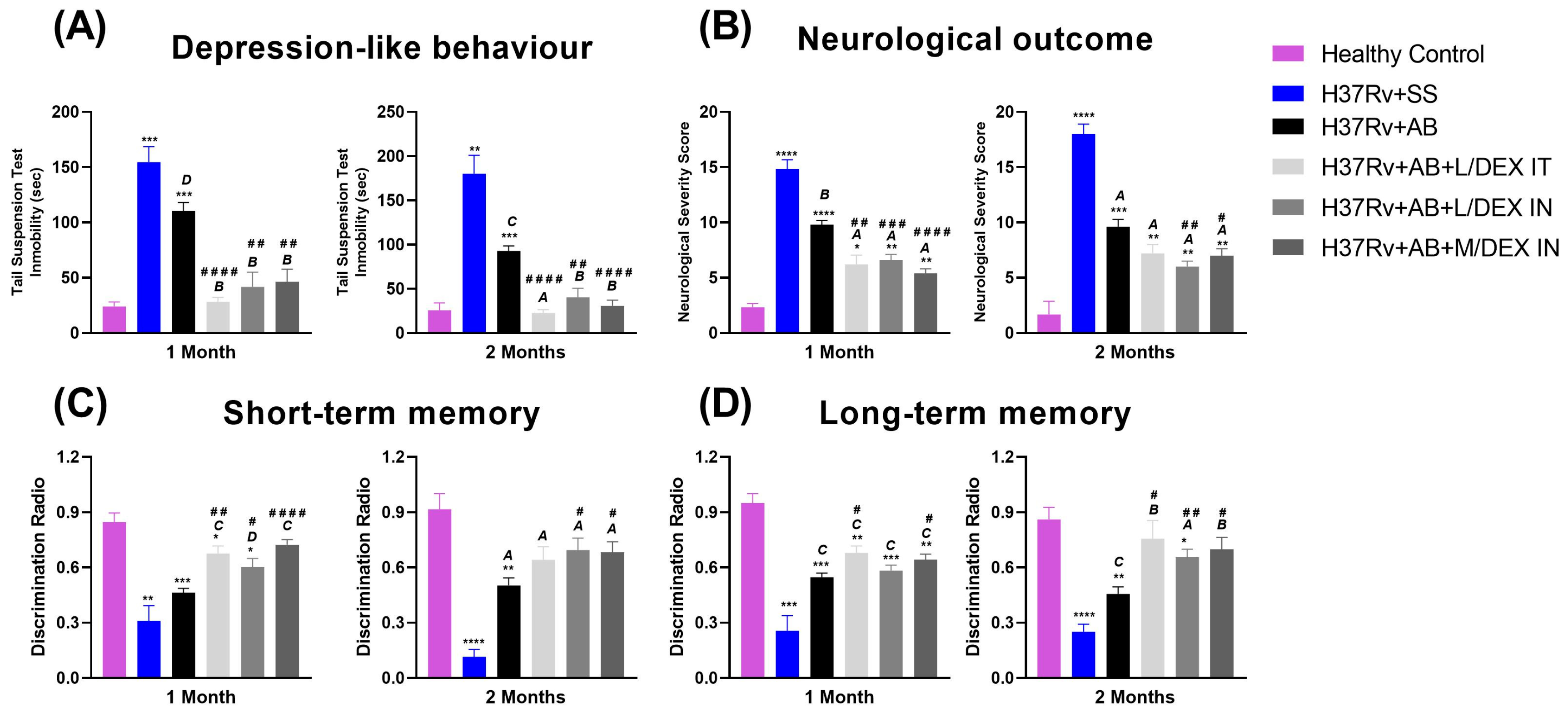
3.3. Low Doses of Dexamethasone in Combination with Antibiotics Decreased the Anxiety-like Behavior, Depression-like Behavior, and Neurological Damage, and Improved Memory of Mice Chronically Infected with Mtb H37Rv
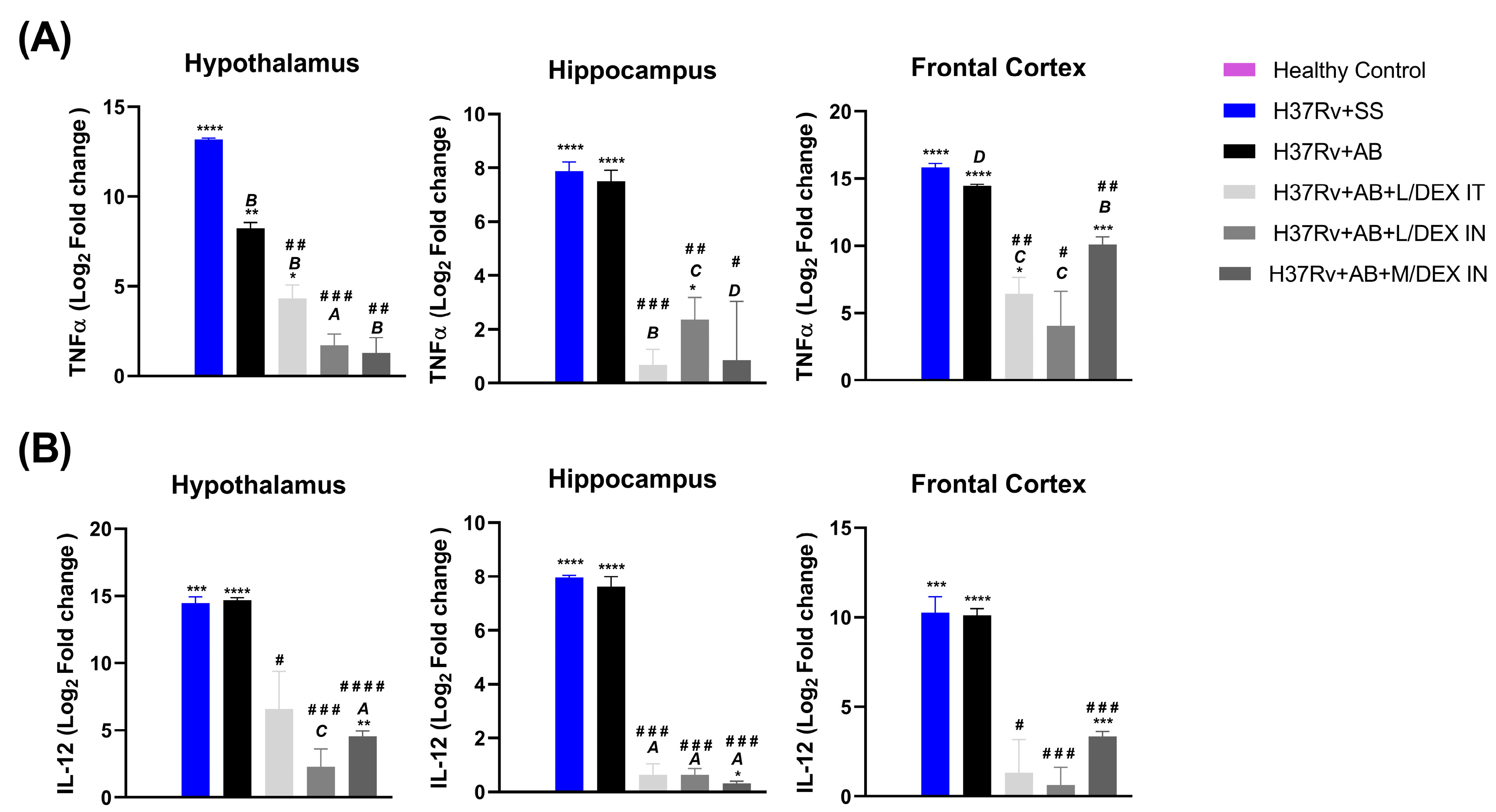
3.4. Low Doses of Dexamethasone in Combination with Antibiotics Decreased the Expression of IL-12 and TNF in Different Brain Areas of Mice Chronically Infected with Mtb H37Rv
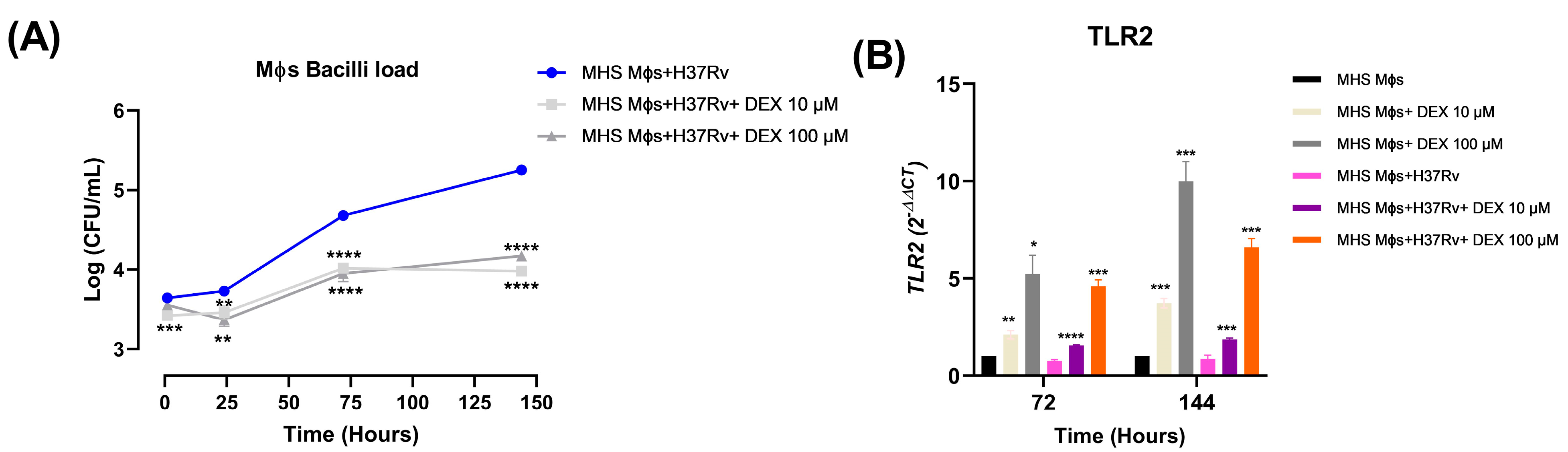
3.5. Low Doses of Dexamethasone Decreased the Bacilli Load and Induced TLR-2 Expression of the Alveolar Macrophages MHS Infected with Mtb H37Rv
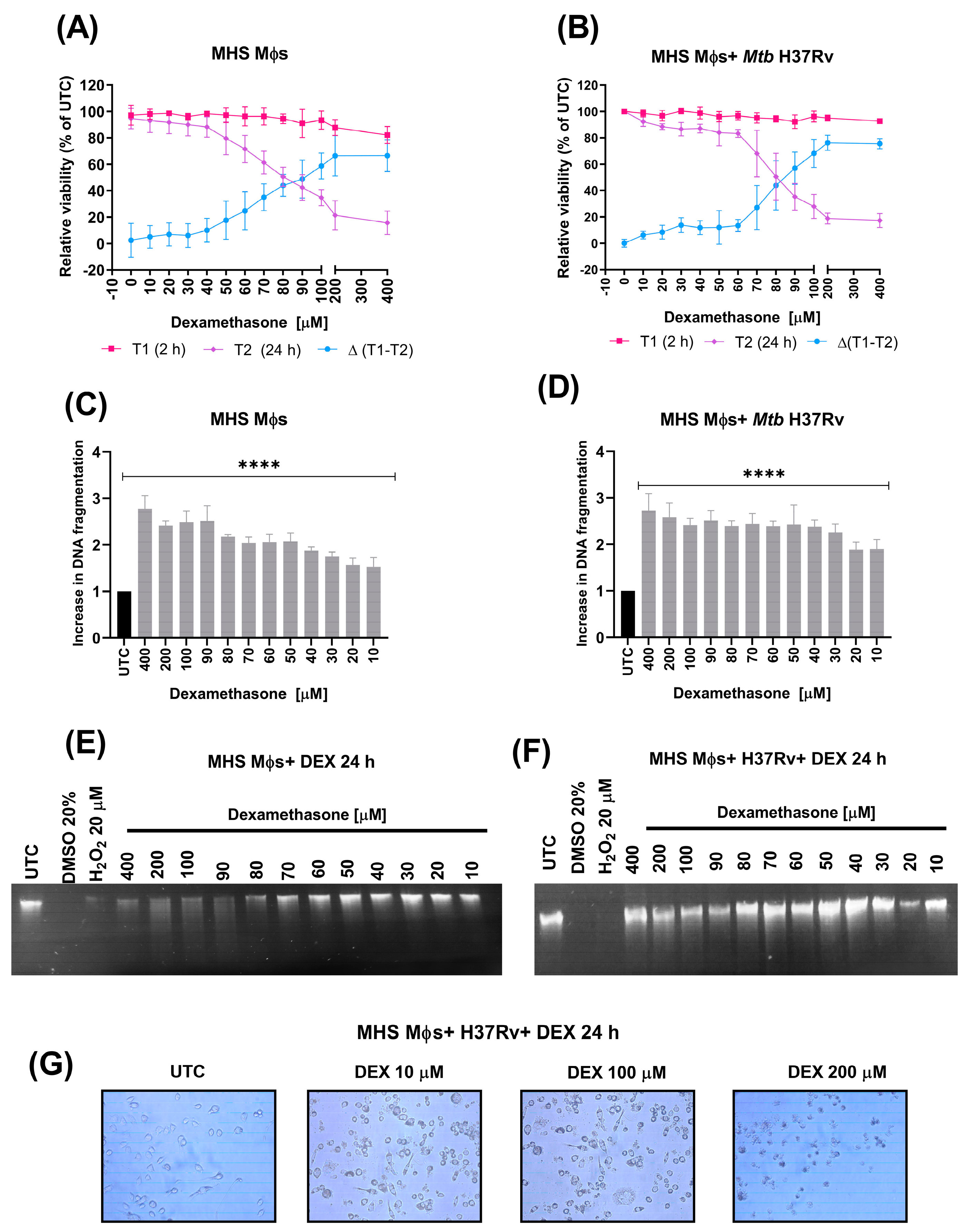
3.6. Dexamethasone Induces Apoptosis in Alveolar Macrophages Not Infected and Infected with Mtb H37Rv
3.7. Dexamethasone Reduces the Anti-Apoptotic Proteins FGFβ, VEGF, and IL6, Reduces Pro- and Anti-Inflammatory Cytokines, and Increases MIP-1α in Alveolar Macrophages Infected with Mtb H37Rv
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Koch, A.; Mizrahi, V. Mycobacterium tuberculosis. Trends Microbiol. 2018, 26, 555–556. [Google Scholar] [CrossRef] [PubMed]
- Migliori, G.B.; Ong, C.W.; Petrone, L.; D’Ambrosio, L.; Centis, R.; Goletti, D. The definition of tuberculosis infection based on the spectrum of tuberculosis disease. Breathe 2021, 17, 210079. [Google Scholar] [CrossRef] [PubMed]
- Global Tuberculosis Report 2022; World Health Organization: Geneva, Switzerland, 2022.
- Gagneux, S. Ecology and evolution of Mycobacterium tuberculosis. Nat. Rev. Microbiol. 2018, 16, 202–213. [Google Scholar] [CrossRef] [PubMed]
- Wallis, R.S. Reconsidering Adjuvant Immunotherapy for Tuberculosis. Clin. Infect. Dis. 2005, 41, 201–208. [Google Scholar] [CrossRef] [Green Version]
- Gopalaswamy, R.; Subbian, S. Corticosteroids for COVID-19 Therapy: Potential Implications on Tuberculosis. Int. J. Mol. Sci. 2021, 22, 3773. [Google Scholar] [CrossRef]
- Franco, L.M.; Gadkari, M.; Howe, K.N.; Sun, J.; Kardava, L.; Kumar, P.; Kumari, S.; Hu, Z.; Fraser, I.D.; Moir, S.; et al. Immune regulation by glucocorticoids can be linked to cell type–dependent transcriptional responses. J. Exp. Med. 2019, 216, 384–406. [Google Scholar] [CrossRef] [Green Version]
- Lannan, E.A.; Galliher-Beckley, A.J.; Scoltock, A.B.; Cidlowski, J.A. Proinflammatory Actions of Glucocorticoids: Glucocorticoids and TNFα Coregulate Gene Expression In Vitro and In Vivo. Endocrinology 2012, 153, 3701–3712. [Google Scholar] [CrossRef] [Green Version]
- Bansal, A.; Mostafa, M.M.; Kooi, C.; Sasse, S.K.; Michi, A.N.; Shah, S.V.; Leigh, R.; Gerber, A.N.; Newton, R. Interplay between nuclear factor-κB, p38 MAPK, and glucocorticoid receptor signaling synergistically induces functional TLR2 in lung epithelial cells. J. Biol. Chem. 2022, 298, 101747. [Google Scholar] [CrossRef]
- Cruz-Topete, D.; Cidlowski, J.A. One Hormone, Two Actions: Anti- and Pro-Inflammatory Effects of Glucocorticoids. Neuroimmunomodulation 2014, 22, 20–32. [Google Scholar] [CrossRef] [Green Version]
- Gräb, J.; Suárez, I.; van Gumpel, E.; Winter, S.; Schreiber, F.; Esser, A.; Hölscher, C.; Fritsch, M.; Herb, M.; Schramm, M.; et al. Corticosteroids inhibit Mycobacterium tuberculosis-induced necrotic host cell death by abrogating mitochondrial membrane permeability transition. Nat. Commun. 2019, 10, 688. [Google Scholar] [CrossRef]
- Lemos, C.X.; Anton, C.; Machado, F.D.; Bernardi, R.M.; Freitas, A.A.; Silva, D.R. Adjunctive corticosteroid therapy in patients with pulmonary tuberculosis. Rev. Assoc. Med. Bras. 2022, 68, 1199–1203. [Google Scholar] [CrossRef] [PubMed]
- Smego, R.A.; Ahmed, N. A systematic review of the adjunctive use of systemic corticosteroids for pulmonary tuberculosis. Int. J. Tuberc. Lung Dis. 2003, 7, 208–213. [Google Scholar] [PubMed]
- Mayanja-Kizza, H.; Jones-Lopez, E.; Okwera, A.; Wallis, R.S.; Ellner, J.J.; Mugerwa, R.D.; Whalen, C.C. Immunoadjuvant Prednisolone Therapy for HIV-Associated Tuberculosis: A Phase 2 Clinical Trial in Uganda. J. Infect. Dis. 2005, 191, 856–865. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meintjes, G.; Wilkinson, R.J.; Morroni, C.; Pepper, D.J.; Rebe, K.; Rangaka, M.X.; Oni, T.; Maartens, G. Randomized placebo-controlled trial of prednisone for paradoxical tuberculosis-associated immune reconstitution inflammatory syndrome. Aids 2010, 24, 2381–2390. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.Y.; Chang-Min, C.; Koh, Y.; Kim, W.-S.; Song, J.-W.; Oh, Y.-M.; Lee, S.-D.; Lee, S.W.; Lee, J.-S.; Lim, C.-M.; et al. Effects of Corticosteroids on Critically Ill Pulmonary Tuberculosis Patients with Acute Respiratory Failure: A Propensity Analysis of Mortality. Clin. Infect. Dis. 2016, 63, 1449–1455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, C.-H.; Kim, K.; Hyun, M.K.; Jang, E.J.; Lee, N.R.; Yim, J.-J. Use of inhaled corticosteroids and the risk of tuberculosis. Thorax 2013, 68, 1105–1113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nazir, A.; Masih, M.; Iqbal, M. Formulation, optimization, qualitative and quantitative analysis of new dosage form of corticosteroid. Futur. J. Pharm. Sci. 2021, 7, 208. [Google Scholar] [CrossRef]
- Giraldo-Velásquez, M.F.; Pérez-Osorio, I.N.; Espinosa-Cerón, A.; Bárcena, B.M.; Calderón-Gallegos, A.; Fragoso, G.; Torres-Ramos, M.; Páez-Martínez, N.; Sciutto, E. Intranasal Methylprednisolone Ameliorates Neuroinflammation Induced by Chronic Toluene Exposure. Pharmaceutics 2022, 14, 1195. [Google Scholar] [CrossRef]
- Meneses, G.; Gevorkian, G.; Florentino, A.; Bautista, M.A.; Espinosa, A.; Acero, G.; Díaz, G.; Fleury, A.; Pérez Osorio, I.N.; del Rey, A.; et al. Intranasal delivery of dexamethasone efficiently controls LPS-induced murine neuroinflammation. Clin. Exp. Immunol. 2017, 190, 304–314. [Google Scholar] [CrossRef] [Green Version]
- Lara-Espinosa, J.V.; Arce-Aceves, M.F.; Mata-Espinosa, D.; Barrios-Payán, J.; Marquina-Castillo, B.; Hernández-Pando, R. The Therapeutic Effect of Intranasal Administration of Dexamethasone in Neuroinflammation Induced by Experimental Pulmonary Tuberculosis. Int. J. Mol. Sci. 2021, 22, 5997. [Google Scholar] [CrossRef]
- Hernández-Pando, R.; Orozcoe, H.; Sampieri, A.; Pavón, L.; Velasquillo, C.; Larriva-Sahd, J.; Alcocer, J.M.; Madrid, M.V. Correlation between the kinetics of Th1, Th2 cells and pathology in a murine model of experimental pulmonary tuberculosis. Immunology 1996, 89, 26–33. [Google Scholar] [PubMed]
- Franklin, K.B.J.; Paxinos, G. Paxinos and Franklin’s the Mouse Brain in Stereotaxic Coordinates; Academic Press: Cambridge, MA, USA; Elsevier: Amsterdam, The Netherlands, 2013; Volume 246. [Google Scholar]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative CT method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef] [PubMed]
- Lara-Espinosa, J.V.; Santana-Martínez, R.A.; Maldonado, P.D.; Zetter, M.; Becerril-Villanueva, E.; Pérez-Sánchez, G.; Pavón, L.; Mata-Espinosa, D.; Barrios-Payán, J.; López-Torres, M.O.; et al. Experimental Pulmonary Tuberculosis in the Absence of Detectable Brain Infection Induces Neuroinflammation and Behavioural Abnormalities in Male BALB/c Mice. Int. J. Mol. Sci. 2020, 21, 9483. [Google Scholar] [CrossRef] [PubMed]
- Nestler, E.J.; Hyman, S.E. Animal models of neuropsychiatric disorders. Nat. Neurosci. 2010, 13, 1161–1169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gilhotra, R.; Goel, S.; Gilhotra, N. Behavioral and biochemical characterization of elevated “I-maze” as animal model of anxiety. Beni-Suef Univ. J. Basic Appl. Sci. 2015, 4, 214–224. [Google Scholar] [CrossRef] [Green Version]
- Stahel, P.F.; Shohami, E.; Younis, F.M.; Kariya, K.; Otto, V.I.; Lenzlinger, P.M.; Grosjean, M.B.; Eugster, H.-P.; Trentz, O.; Kossmann, T.; et al. Experimental Closed Head Injury: Analysis of Neurological Outcome, Blood–Brain Barrier Dysfunction, Intracranial Neutrophil Infiltration, and Neuronal Cell Death in Mice Deficient in Genes for Pro-Inflammatory Cytokines. J. Cereb. Blood Flow Metab. 2000, 20, 369–380. [Google Scholar] [CrossRef] [Green Version]
- Ennaceur, A.; Neave, N.; Aggleton, J.P. Spontaneous object recognition and object location memory in rats: The effects of lesions in the cingulate cortices, the medial prefrontal cortex, the cingulum bundle and the fornix. Exp. Brain Res. 1997, 113, 509–519. [Google Scholar] [CrossRef]
- Helm, K.; Beyreis, M.; Mayr, C.; Ritter, M.; Jakab, M.; Kiesslich, T.; Plaetzer, K. In Vitro Cell Death Discrimination and Screening Method by Simple and Cost-Effective Viability Analysis. Cell. Physiol. Biochem. 2017, 41, 1011–1019. [Google Scholar] [CrossRef]
- Espinosa-Pereiro, J.; Sánchez-Montalvá, A.; Aznar, M.L.; Espiau, M. MDR Tuberculosis Treatment. Medicina 2022, 58, 188. [Google Scholar] [CrossRef]
- Lange, C.; Barry, C.E., III; Horsburgh, C.R., Jr. Treatments of Multidrug-Resistant Tuberculosis: Light at the End of the Tunnel. Am. J. Respir. Crit. Care Med. 2022, 205, 1142–1144. [Google Scholar] [CrossRef]
- Madamsetty, V.S.; Mohammadinejad, R.; Uzieliene, I.; Nabavi, N.; Dehshahri, A.; García-Couce, J.; Tavakol, S.; Moghassemi, S.; Dadashzadeh, A.; Makvandi, P.; et al. Dexamethasone: Insights into Pharmacological Aspects, Therapeutic Mechanisms, and Delivery Systems. ACS Biomater. Sci. Eng. 2022, 8, 1763–1790. [Google Scholar] [CrossRef] [PubMed]
- Young, C.; Walzl, G.; Du Plessis, N. Therapeutic host-directed strategies to improve outcome in tuberculosis. Mucosal Immunol. 2020, 13, 190–204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, L.; Zhou, M.; Ma, J.; Liu, X.; Chen, P.; Zhu, H.; Tang, Q.; Cheng, T.; Guan, Y.; Xia, N. Dexamethasone ameliorates severe pneumonia but slightly enhances viral replication in the lungs of SARS-CoV-2-infected Syrian hamsters. Cell. Mol. Immunol. 2022, 19, 290–292. [Google Scholar] [CrossRef] [PubMed]
- Kesavan, A.K.; Mendez, S.E.; Hatem, C.L.; Lopez-Molina, J.; Aird, K.; Pitt, M.L.M.; Dannenberg, A.M.; Manabe, Y.C. Effects of Dexamethasone and Transient Malnutrition on Rabbits Infected with Aerosolized Mycobacterium tuberculosis CDC1551. Infect. Immun. 2005, 73, 7056–7060. [Google Scholar] [CrossRef] [Green Version]
- DE Kaspar, H.M.; Ta, C.N.; Engelbert, M.; Mette, M.; Thiel, M.; Kampik, A. Effects of intravitreal corticosteroid in the treatment of Staphylococcus aureus-induced experimental endophthalmitis. Retina 2008, 28, 326–332. [Google Scholar] [CrossRef]
- Toossi, Z. The Inflammatory Response in Mycobacterium Tuberculosis Infection. In Inflammation; Górski, A., Krotkiewski, H., Zimecki, M., Eds.; Springer: Dordrecht, The Netherlands, 2001; pp. 139–151. [Google Scholar] [CrossRef]
- Huang, C.; Irwin, M.; Wong, G.T.C.; Chang, R.C.C. Evidence of the impact of systemic inflammation on neuroinflammation from a non-bacterial endotoxin animal model. J. Neuroinflamm. 2018, 15, 147. [Google Scholar] [CrossRef] [Green Version]
- Xin, Y.; Tian, M.; Deng, S.; Li, J.; Yang, M.; Gao, J.; Pei, X.; Wang, Y.; Tan, J.; Zhao, F.; et al. The Key Drivers of Brain Injury by Systemic Inflammatory Responses after Sepsis: Microglia and Neuroinflammation. Mol. Neurobiol. 2023, 60, 1369–1390. [Google Scholar] [CrossRef]
- Feiger, J.A.M.; Snyder, R.L.M.; Walsh, M.J.B.; Cissne, M.B.; Cwiek, A.B.; Al-Momani, S.I.M.; Chiou, K.S. The Role of Neuroinflammation in Neuropsychiatric Disorders Following Traumatic Brain Injury: A Systematic Review. J. Head Trauma Rehabil. 2022, 37, E370–E382. [Google Scholar] [CrossRef]
- Becerril-Villanueva, E.; Ponce-Regalado, M.D.; Pérez-Sánchez, G.; Salazar-Juárez, A.; Arreola, R.; Álvarez-Sánchez, M.E.; Juárez-Ortega, M.; Falfán-Valencia, R.; Hernández-Pando, R.; Morales-Montor, J.; et al. Chronic infection with Mycobacterium lepraemurium induces alterations in the hippocampus associated with memory loss. Sci. Rep. 2018, 8, 9063. [Google Scholar] [CrossRef] [Green Version]
- Ponce-Regalado, M.; Salazar-Juárez, A.; Rojas-Espinosa, O.; Contis-Montes de Oca, A.; Hurtado-Alvarado, G.; Arce-Paredes, P.; Pérez-Sánchez, G.; Pavón, L.; Girón-Pérez, M.; Hernández-Pando, R.; et al. Development of Anxiolytic and Depression-like Behavior in Mice Infected with Mycobacterium lepraemurium. Neuroscience 2022, 493, 15–30. [Google Scholar] [CrossRef]
- Babu, G.N.; Kumar, A.; Kalita, J.; Misra, U. Proinflammatory cytokine levels in the serum and cerebrospinal fluid of tuberculous meningitis patients. Neurosci. Lett. 2008, 436, 48–51. [Google Scholar] [CrossRef] [PubMed]
- Erdő, F.; Bors, L.A.; Farkas, D.; Bajza, Á.; Gizurarson, S. Evaluation of intranasal delivery route of drug administration for brain targeting. Brain Res. Bull. 2018, 143, 155–170. [Google Scholar] [CrossRef]
- Borkar, S.P.; Raizaday, A. Different Strategies for Nose-to-Brain Delivery of Small Molecules. In Nasal Drug Delivery; Pathak, Y.V., Yadav, H.K.S., Eds.; Springer: Cham, Switzerland, 2023. [Google Scholar] [CrossRef]
- Espinosa, A.; Meneses, G.; Chavarría, A.; Mancilla, R.; Pedraza-Chaverri, J.; Fleury, A.; Bárcena, B.; Pérez-Osorio, I.N.; Besedovsky, H.; Arauz, A.; et al. Intranasal Dexamethasone Reduces Mortality and Brain Damage in a Mouse Experimental Ischemic Stroke Model. Neurotherapeutics 2020, 17, 1907–1918. [Google Scholar] [CrossRef]
- Rassy, D.; Bárcena, B.; Pérez-Osorio, I.N.; Espinosa, A.; Navarrete-Peón, A.; Terrazas, L.I.; Meneses, G.; Besedovsky, H.O.; Fragoso, G.; Sciutto, E. Intranasal Methylprednisolone Effectively Reduces Neuroinflammation in Mice with Experimental Autoimmune Encephalitis. J. Neuropathol. Exp. Neurol. 2020, 79, 226–237. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Osorio, I.N.; Espinosa, A.; Velázquez, M.G.; Padilla, P.; Bárcena, B.; Fragoso, G.; Jung-Cook, H.; Besedovsky, H.; Meneses, G.; Conde, E.L.S. Nose-to-Brain Delivery of Dexamethasone: Biodistribution Studies in Mice. J. Pharmacol. Exp. Ther. 2021, 378, 244–250. [Google Scholar] [CrossRef] [PubMed]
- Cárdenas, G.; Bobes, R.J.; Fragoso, G.; Pérez-Osorio, N.I.; Hernández, M.; Espinosa, A.; Fleury, A.; Flores, J.; Laclette, J.P.; Sciutto, E.; et al. Pharmacokinetic Study of Intranasal Dexamethasone and Methylprednisolone Compared with Intravenous Administration: Two Open-Label, Single-Dose, Two-Period, Two-Sequence, Cross-Over Study in Healthy Volunteers. Pharmaceutics 2023, 15, 105. [Google Scholar] [CrossRef]
- Amponsah, S.K.; Adams, I. Drug Absorption via the Nasal Route: Opportunities and Challenges. In Nasal Drug Delivery; Pathak, Y.V., Yadav, H.K.S., Eds.; Springer: Cham, Switzerland, 2023. [Google Scholar] [CrossRef]
- Ferreira, T.P.T.; Lima, J.G.M.E.; Farias-Filho, F.A.; De Sá, Y.A.P.J.; De Arantes, A.C.S.; Guimarães, F.V.; Carvalho, V.D.F.; Hogaboam, C.; Wallace, J.; Martins, M.A.; et al. Intranasal Flunisolide Suppresses Pathological Alterations Caused by Silica Particles in the Lungs of Mice. Front. Endocrinol. 2020, 11, 1–12. [Google Scholar] [CrossRef]
- Porzionato, A.; Zaramella, P.; Dedja, A.; Guidolin, D.; Van Wemmel, K.; Macchi, V.; Jurga, M.; Perilongo, G.; De Caro, R.; Baraldi, E.; et al. Intratracheal administration of clinical-grade mesenchymal stem cell-derived extracellular vesicles reduces lung injury in a rat model of bronchopulmonary dysplasia. Am. J. Physiol. Lung Cell. Mol. Physiol. 2019, 316, L6–L19. [Google Scholar] [CrossRef]
- Behrens, R.; McGovern, A.E.; Farrell, M.J.; Moe, A.A.K.; Mazzone, S.B. Mini Review: Central Organization of Airway Afferent Nerve Circuits. Neurosci. Lett. 2021, 744, 135604. [Google Scholar] [CrossRef]
- Li, C.; Chen, W.; Lin, F.; Li, W.; Wang, P.; Liao, G.; Zhang, L. Functional Two-Way Crosstalk Between Brain and Lung: The Brain–Lung Axis. Cell. Mol. Neurobiol. 2023, 43, 991–1003. [Google Scholar] [CrossRef]
- Tükenmez, H.; Edström, I.; Kalsum, S.; Braian, C.; Ummanni, R.; Fick, S.B.; Sundin, C.; Lerm, M.; Elofsson, M.; Larsson, C. Corticosteroids protect infected cells against mycobacterial killing in vitro. Biochem. Biophys. Res. Commun. 2019, 511, 117–121. [Google Scholar] [CrossRef] [PubMed]
- Oliviera Nascimento, L.; Massari, P.; Wetzler, L.M. The Role of TLR2 in Infection and Immunity. Front. Immunol. 2012, 3, 79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Homma, T.; Kato, A.; Hashimoto, N.; Batchelor, J.; Yoshikawa, M.; Imai, S.; Wakiguchi, H.; Saito, H.; Matsumoto, K. Corticosteroid and Cytokines Synergistically Enhance Toll-Like Receptor 2 Expression in Respiratory Epithelial Cells. Am. J. Respir. Cell Mol. Biol. 2004, 31, 463–469. [Google Scholar] [CrossRef] [Green Version]
- Imasato, A.; Desbois-Mouthon, C.; Han, J.; Kai, H.; Cato, A.C.B.; Akira, S.; Li, J.-D. Inhibition of p38 MAPK by Glucocorticoids via Induction of MAPK Phosphatase-1 Enhances Nontypeable Haemophilus influenzae-induced Expression of Toll-like Receptor 2. J. Biol. Chem. 2002, 277, 47444–47450. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoppstädter, J.; Dembek, A.; Linnenberger, R.; Dahlem, C.; Barghash, A.; Fecher-Trost, C.; Fuhrmann, G.; Koch, M.; Kraegeloh, A.; Huwer, H.; et al. Toll-Like Receptor 2 Release by Macrophages: An Anti-inflammatory Program Induced by Glucocorticoids and Lipopolysaccharide. Front. Immunol. 2019, 10, 1634. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, P.; Aulakh, G.K. Modulation of low-dose ozone and LPS exposed acute mouse lung inflammation by IF1 mediated ATP hydrolysis inhibitor, BTB06584. Front. Immunol. 2023, 14, 1126574. [Google Scholar] [CrossRef]
- Bhavsar, I.; Miller, C.S.; Al-Sabbagh, M. Macrophage Inflammatory Protein-1 Alpha (MIP-1 alpha)/CCL3: As a Biomarker. Gen. Methods Biomark. Res. Appl. 2015, 223–249. [Google Scholar] [CrossRef]
- Zeng, S.; Qiao, H.; Lv, X.-W.; Fan, D.; Liu, T.; Xie, D. High-dose dexamethasone induced LPS-stimulated rat alveolar macrophages apoptosis. Drug Des. Dev. Ther. 2017, 11, 3097–3104. [Google Scholar] [CrossRef] [Green Version]
- Kovalovich, K.; Li, W.; DeAngelis, R.; Greenbaum, L.E.; Ciliberto, G.; Taub, R. Interleukin-6 Protects against Fas-mediated Death by Establishing a Critical Level of Anti-apoptotic Hepatic Proteins FLIP, Bcl-2, and Bcl-xL. J. Biol. Chem. 2001, 276, 26605–26613. [Google Scholar] [CrossRef] [Green Version]
- Katoh, M.; Nakagama, H. FGF Receptors: Cancer Biology and Therapeutics. Med. Res. Rev. 2014, 34, 280–300. [Google Scholar] [CrossRef]
- Malemud, C.J. Growth hormone, VEGF and FGF: Involvement in rheumatoid arthritis. Clin. Chim. Acta 2007, 375, 10–19. [Google Scholar] [CrossRef] [PubMed]
- Lam, A.; Prabhu, R.; Gross, C.M.; Riesenberg, L.A.; Singh, V.; Aggarwal, S. Role of apoptosis and autophagy in tuberculosis. Am. J. Physiol.-Lung Cell. Mol. Physiol. 2017, 313, L218–L229. [Google Scholar] [CrossRef] [PubMed] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward | Reverse |
|---|---|---|
| GAPDH | 5′-CATTGTGGAAGGGCTATGA-3′ | 5′-GGAAGGCCATGCCAGTGAGC-3′ |
| TLR-2 | 5′-TGCTTTCCTGCTGGAGATTT-3′ | 5′-TGTAACGCAACAGCTTCAGG-3′ |
| TNF | 5′-GCCGAGAAAGGCTGCTTG-3′ | 5′-TGTGGCTTCGACCTCTACCTC-3′ |
| IL12 | 5′-GGATGGAAGAGTCCCCCAAA-3′ | 5′-GCTCTGCGGGCATTTAACAT-3′ |
| Protein (pg/mL) | Mϕs | Mϕs + H37Rv | Mϕs + H37Rv + DEX 10 μM | Mϕs + H37Rv + DEX 100 μM |
|---|---|---|---|---|
| Mitogenic activity | ||||
| FGF-β | 82.29 ± 3.225 | 97.99 ± 2.751 ** | 72.26 ± 7.10 C | 64.24 ± 2.232 **/A |
| IL-6 | 241.6 ± 54.59 | 628.74 ± 53.37 *** | 132.5 ± 18.46 A | 101.40 ± 16.04 */A |
| VEGF | 1203 ± 215.5 | 2761 ± 584.7 * | 23.06 ± 6.645 ***/B | 31.68 ± 12.20 ***/B |
| Anti-inflammatory | ||||
| IL-10 | 31.19 ± 6.051 | 54.16 ± 3.566 ** | 35.24 ± 6.283 D | 20.10 ± 3.164 A |
| IL-13 | 18.00 ± 2.843 | 26.13 ± 2.45 | 4.709 ± 0.5931 ***/A | 5.753 ± 1.085 **/A |
| IL-5 | 13.96 ± 0.9218 | 14.53 ± 1.057 | 8.042 ± 1.809 */D | 8.377 ± 0.7788 **/B |
| Proinflammatory | ||||
| IL-1β | 84.58 ± 5.483 | 105.2 ± 3.227 ** | 80.93 ± 5.02 C | 71.04 ± 4.384 A |
| IL1-α | 45.25 ± 8.864 | 114.3 ± 11.05 ** | 23.64 ± 7.257 A | 10.04 ± 3.411 **/A |
| IL-12 | 17.83 ± 2.464 | 28.01 ± 3.055 * | 11.75 ± 2.72 C | 9.51 ± 3.018 C |
| IFN-γ | 9.482 ± 1.926 | 15.22 ± 1.042 * | 5.40 ± 2.435 C | 1.837 ± 0.6193 **/A |
| TNF | 88.32 ± 11.34 | 130.5 ± 9.615 * | 81.81 ± 11.83 C | 60.49 ± 17.24 C |
| GM-CSF | 16.05 ± 2.223 | 73.44 ± 24.63 | 26.85 ± 7.354 | 7.700 ± 1.974 */D |
| MIP-1α | 20.86 ± 2.641 | 5.685 ± 1.644 *** | 16338 ± 25.40 ****/A | 17726 ± 18.75 ****/A |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lara-Espinosa, J.V.; Arce-Aceves, M.F.; Barrios-Payán, J.; Mata-Espinosa, D.; Lozano-Ordaz, V.; Becerril-Villanueva, E.; Ponce-Regalado, M.D.; Hernández-Pando, R. Effect of Low Doses of Dexamethasone on Experimental Pulmonary Tuberculosis. Microorganisms 2023, 11, 1554. https://doi.org/10.3390/microorganisms11061554
Lara-Espinosa JV, Arce-Aceves MF, Barrios-Payán J, Mata-Espinosa D, Lozano-Ordaz V, Becerril-Villanueva E, Ponce-Regalado MD, Hernández-Pando R. Effect of Low Doses of Dexamethasone on Experimental Pulmonary Tuberculosis. Microorganisms. 2023; 11(6):1554. https://doi.org/10.3390/microorganisms11061554
Chicago/Turabian StyleLara-Espinosa, Jacqueline V., María Fernanda Arce-Aceves, Jorge Barrios-Payán, Dulce Mata-Espinosa, Vasti Lozano-Ordaz, Enrique Becerril-Villanueva, María Dolores Ponce-Regalado, and Rogelio Hernández-Pando. 2023. "Effect of Low Doses of Dexamethasone on Experimental Pulmonary Tuberculosis" Microorganisms 11, no. 6: 1554. https://doi.org/10.3390/microorganisms11061554
APA StyleLara-Espinosa, J. V., Arce-Aceves, M. F., Barrios-Payán, J., Mata-Espinosa, D., Lozano-Ordaz, V., Becerril-Villanueva, E., Ponce-Regalado, M. D., & Hernández-Pando, R. (2023). Effect of Low Doses of Dexamethasone on Experimental Pulmonary Tuberculosis. Microorganisms, 11(6), 1554. https://doi.org/10.3390/microorganisms11061554