
Mucus-Trap-Assisted Feeding Is a Common Strategy of the Small Mixoplanktonic Prorocentrum pervagatum and P. cordatum (Prorocentrales, Dinophyceae)


Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains and Growth Conditions
2.2. Microscopy
2.3. Experiments
2.4. Modelling
3. Results
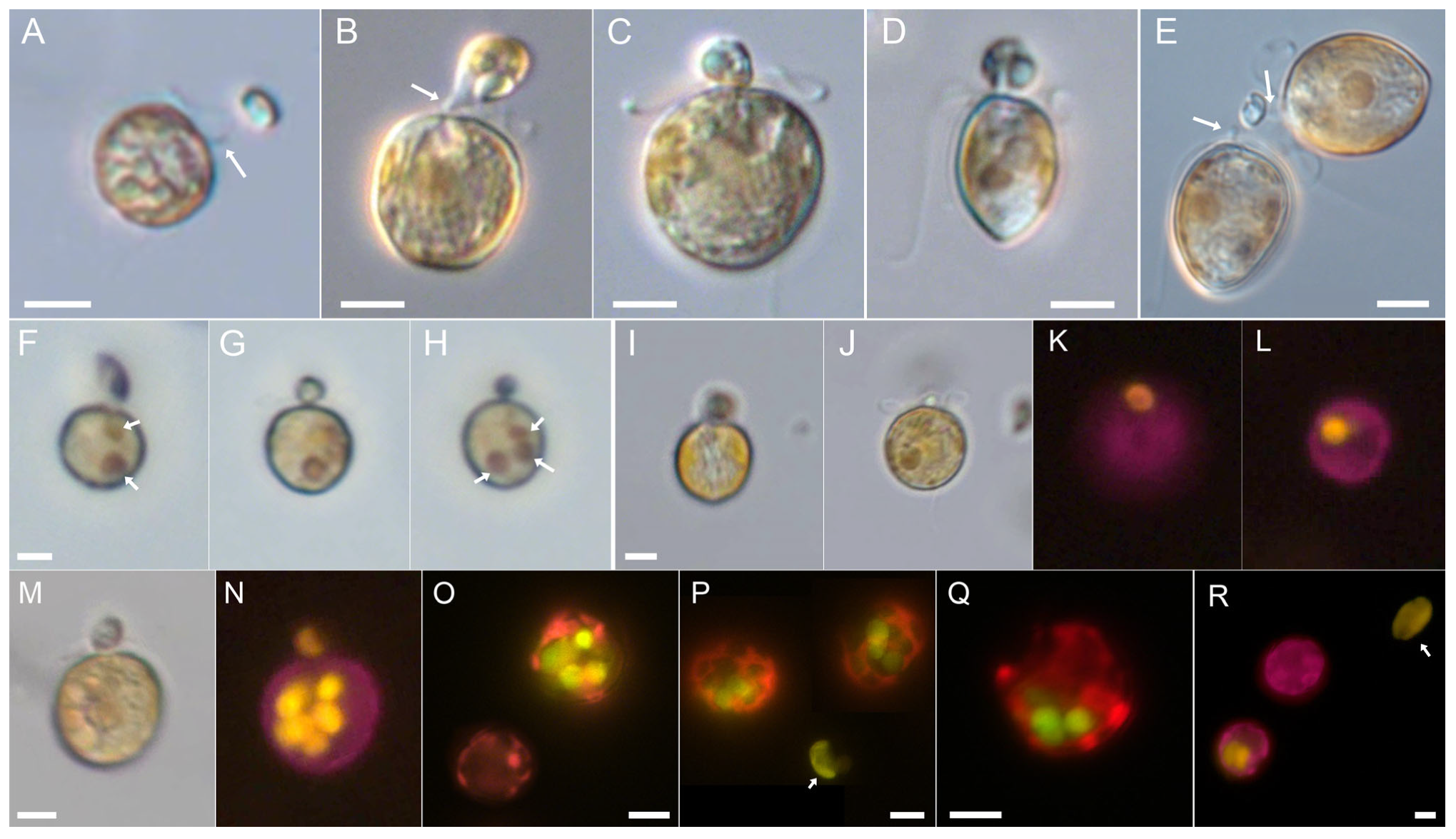
3.1. Prorocentrum pervagatum
3.2. Interactions between P. pervagatum and the Cryptophyte Prey Teleaulax amphioxeia
3.3. Prorocentrum cordatum
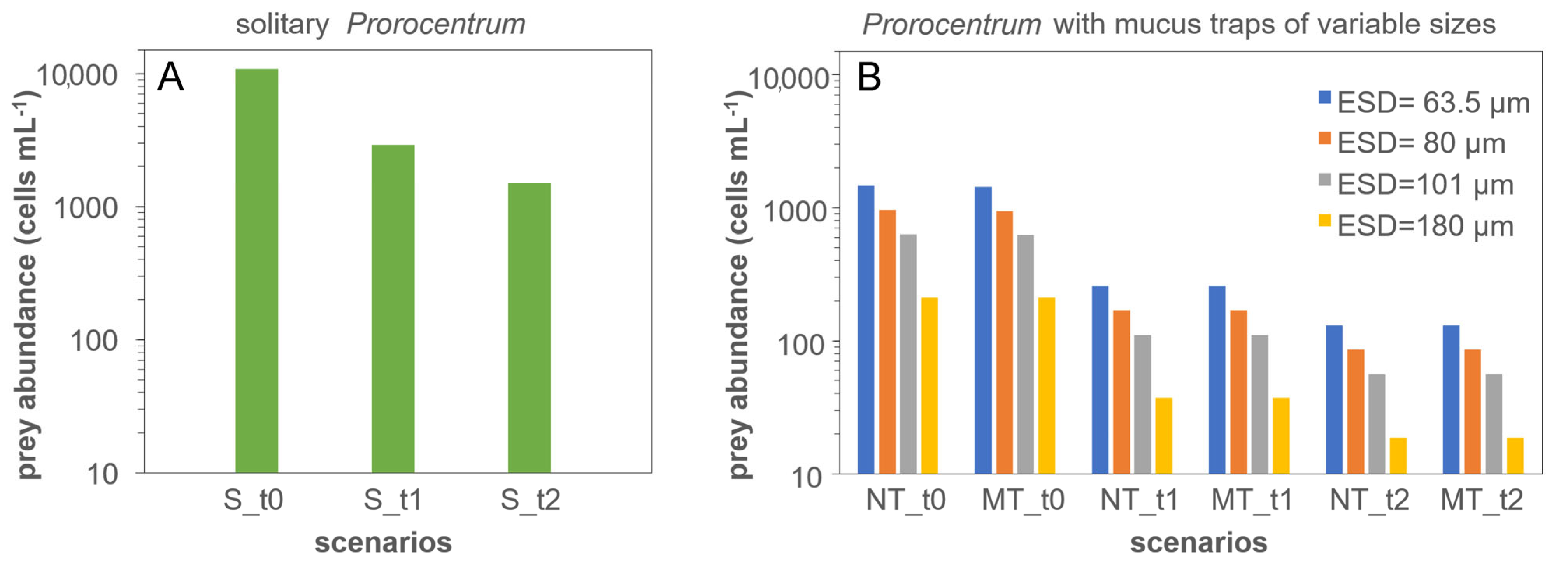
3.4. Modelling
4. Discussion
4.1. Production and Deployment of Mucus Traps
4.2. Phagotrophic Feeding Mechanism in Prorocentrum
4.3. Advantages for Phagotrophy through Trap Deployment
4.4. Mucus Structures Constructed by Dinoflagellates
4.5. Is Mucus-Trap-Assisted Phagomixotrophy More Widespread in Prorocentrum?
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dodge, J.D. The Prorocentrales (Dinophyceae). II Revision of the taxonomy within the genus Prorocentrum. Bot. J. Linn. Soc. 1975, 71, 103–125. [Google Scholar] [CrossRef]
- Hoppenrath, M.; Chomérat, N.; Horiguchi, T.; Schweikert, M.; Nagahama, Y.; Murray, S. Taxonomy and phylogeny of the benthic Prorocentrum species (Dinophyceae)—A proposal and review. Harmful Algae 2013, 27, 1–28. [Google Scholar] [CrossRef]
- Sohn, M.H.; Lim, S.; Seo, K.W.; Lee, S.J. Effect of ambient medium viscosity on the motility and flagella motion of Prorocentrum minimum (Dinophyceae). J. Plankton Res. 2013, 35, 1294–1304. [Google Scholar] [CrossRef]
- Mitra, A.; Caron, D.A.; Faure, E.; Flynn, K.J.; Goncales Leles, S.; Hansen, P.J.; McManus, G.B.; Not, F.; do Rosario Gomez, H.; Santoferrara, L.F.; et al. The mixoplankton database (MDB): Diversity of photo-phago-trophic plankton in form, function, and distribution across the global ocean. J. Eukaryot. Microbiol. 2023, e12972. [Google Scholar] [CrossRef]
- Hansen, P.J.; Calado, A.J. Phagotrophic mechanisms and prey selection in free-living dinoflagellates. J. Eukaryot. Microbiol. 1999, 46, 382–389. [Google Scholar] [CrossRef]
- Schnepf, E.; Elbrächter, M. Nutritional Strategies in Dinoflagellates. A Review with Emphasis on Cell Biological Aspects. Eur. J. Protistol. 1992, 28, 3–24. [Google Scholar] [CrossRef]
- Jeong, H.J.; Yoo, Y.D.; Park, J.Y.; Song, J.Y.; Kim, S.T.; Lee, S.H.; Kim, K.Y.; Yih, W.H. Feeding by phototrophic red-tide dinoflagellates: Five species newly revealed and six species previously known to be mixotrophic. Aquat. Microb. Ecol. 2005, 40, 133–150. [Google Scholar] [CrossRef] [Green Version]
- Johnson, M.D. Inducible mixotrophy in the dinoflagellate Prorocentrum minimum. J. Eukaryot. Microbiol. 2015, 62, 431–443. [Google Scholar] [CrossRef]
- Larsson, M.E.; Bramucci, A.R.; Collins, S.; Hallegraeff, G.; Kahlke, T.; Raina, J.B.; Seymour, J.R.; Doblin, M.A. Mucospheres produced by a mixotrophic protist impact ocean carbon cycling. Nat. Commun. 2022, 13, 1301. [Google Scholar] [CrossRef]
- Tillmann, U.; Gottschling, M.; Wietkamp, S.; Hoppenrath, M. Morphological and phylogenetic characterisation of Prorocentrum spinulentum sp. nov. (Prorocentrales, Dinophyceae), a small spiny species from the North Atlantic. Microorganisms 2023, 11, 271. [Google Scholar] [CrossRef]
- Gómez, F.; Zhang, H.; Roselli, L.; Lin, S. Detection of Prorocentrum shikokuense in the Mediterranean Sea and evidence that P. dentatum, P. obtusidens and P. shikokuense are three different species (Prorocentrales, Dinophyceae). Acta Protozool. 2022, 60, 47–59. [Google Scholar] [CrossRef]
- Tillmann, U.; Wietkamp, S.; Gottschling, M.; Hoppenrath, M. Prorocentrum pervagatum sp. nov. (Prorocentrales, Dinophyceae): A new, small, planktonic species with a global distribution. Phycol. Res. 2023, 71, 56–71. [Google Scholar] [CrossRef]
- Gómez, F.; Gourvil, P.; Li, T.; Huang, Y.; Zhang, H.; Courcot, L.; Artigas, L.F.; Onis, E.S.; Gutiérrez-Rodriguez, A.; Lin, S. Molecular phylogeny of the spiny-surfaced species of the dinoflagellate Prorocentrum with the description of P. thermophilum sp. nov. and P. criophilum sp. nov. (Prorocentrales, Dinophyceae). J. Phycol. 2023, 59, 70–86. [Google Scholar] [CrossRef]
- Keller, M.D.; Selvin, R.C.; Claus, W.; Guillard, R.R.L. Media for the culture of oceanic ultraphytoplankton. J. Phycol. 1987, 23, 633–638. [Google Scholar] [CrossRef]
- Frost, B.W. Effects of size and concentration of food particles on the feeding behavior of the marine planktonic copepod Calanus pacificus. Limnol. Oceanogr. 1972, 17, 805–815. [Google Scholar] [CrossRef] [Green Version]
- Heinbokel, J.F. Studies on the functional role of tintinnids in the southern California Bight. I. Grazing and growth rates in laboratory cultures. Mar. Biol. 1978, 47, 177–189. [Google Scholar] [CrossRef]
- Flynn, K.; Mitra, A. Why plankton modelers should reconsider using rectangular hyperbolic (Michaelis-Menten, Monod) descriptions of predator-prey interactions. Front. Mar. Sci. 2016, 3, 165. [Google Scholar] [CrossRef] [Green Version]
- Blossom, H.; Daugbjerg, N.; Hansen, P.J. Toxic mucus traps: A novel mechanism that mediates prey uptake in the mixotrophic dinoflagellate Alexandrium pseudogonyaulax. Harmful Algae 2012, 17, 40–53. [Google Scholar] [CrossRef]
- Ojamäe, K.; Hansen, P.J.; Lips, I. Mass entrapment and lysis of Mesodinium rubrum cells in mucus threads observed in cultures with Dinophysis. Harmful Algae 2016, 55, 77–84. [Google Scholar] [CrossRef]
- Giussani, V.; Sbrana, F.; Asnaghi, V.; Vassalli, M.; Faimali, M.; Casabianca, S.; Penna, A.; Ciminiello, P.; Dell’Aversano, C.; Tartaglione, L.; et al. Active role of the mucilage in the toxicity mechanism of the harmful benthic dinoflagellate Ostreopsis cf. ovata. Harmful Algae 2015, 44, 46–53. [Google Scholar] [CrossRef]
- Li, A.; Stoecker, D.K.; Coats, D.W.; Adam, E.J. Ingestion of fluorescently labeled and phycoerythrin-containing prey by mixotrophic dinoflagellates. Aquat. Microb. Ecol. 1996, 10, 139–147. [Google Scholar] [CrossRef]
- Stoecker, D.K.; Li, A.; Coats, D.W.; Gustafson, D.E.; Nannen, M.K. Mixotrophy in the dinoflagellate Prorocentrum minimum. Mar. Ecol. Prog. Ser. 1997, 152, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Wikfors, G.H.; Fernandez, E. Induced bacteriovory in a declining culture of the mixotrophic dinoflagellate Prorocentrum minimum (Pavillard) Schiller. Int. J. Ecol. 2013, 2013, 234372. [Google Scholar] [CrossRef] [Green Version]
- Berdieva, M.; Fel, A.; Kalinina, V.; Skarlato, S.O.; Matantseva, O. Induced phagotrophy in the mixotrophic dinoflagellate Prorocentrum cordatum: Exploring the role of cytoskeleton in prey ingestion. Protistology 2020, 14, 178–185. [Google Scholar] [CrossRef]
- Matantseva, O.; Berdieva, M.; Kalinina, V.; Pozdnyakov, I.; Pechkovskaya, S.; Skarlato, S. Stressor-induced ecdysis and thecate cyst formation in the armoured dinoflagellate Prorocentrum cordatum. Sci. Rep. 2020, 10, 18322. [Google Scholar] [CrossRef] [PubMed]
- Berdieva, M.; Kalinina, V.; Lomert, E.; Knyazev, N.; Skarlato, S. Life cycle stages and evidence for sexual reproduction in the marine dinoflagellate Prorocentrum minimum (Dinophyceae, Prorocentrales). J. Phycol. 2020, 56, 941–952. [Google Scholar] [CrossRef]
- Skarlato, S.; Telesh, S.O.; Matantseva, O.; Pozdnyakov, I.; Berdieva, M.; Schubert, H.; Filatova, N.A.; Knyazev, N.A.; Pechkovskaya, S.A. Studies of bloom-forming dinoflagellates Prorocentrum minimum in fluctuating environment: Contribution to aquatic ecology, cell biology and invasion theory. Protistology 2018, 12, 113–157. [Google Scholar] [CrossRef]
- Kalinina, V.; Matantseva, O.; Berdieva, M.; Skarlato, S. Trophic strategies in dinoflagellates: How nutrients pass through the amphiesma. Protistology 2018, 12, 3–11. [Google Scholar] [CrossRef]
- Berdieva, M.; Pozdnyakov, I.; Matantseva, O.; Knyazev, N.; Skarlato, S. Actin as a cytoskeletal basis for cell architecture and a protein essential for ecdysis in Prorocentrum minimum (Dinophyceae, Prorocentrales). Phycol. Res. 2018, 66, 127–136. [Google Scholar] [CrossRef]
- Telesh, I.V.; Schubert, H.; Skarlato, S.O. Ecological niche partitioning of the invasive dinoflagellate Prorocentrum minimum and its negarive congeners in the Baltic Sea. Harmful Algae 2016, 59, 100–111. [Google Scholar] [CrossRef]
- Khanaychenko, A.N.; Telesh, I.V.; Skarlato, S.O. Bloom-forming potentially toxic dinoflagellates Prorocentrum cordatum in marine plankton food webs. Protistology 2019, 13, 95–125. [Google Scholar] [CrossRef]
- Schnepf, E.; Winter, S. A microtubular basket in the armoured dinoflagellate Prorocentrum micans (Dinophyceae). Arch. Protistenkd. 1990, 138, 89–91. [Google Scholar] [CrossRef]
- Malcolm, S. Aspects of the Biology and Ultrastructure of Prorocentrum spp. (Pyrrhophyta). Master’s Thesis, University of Melbourne, Melbourne, Australia, 1987. [Google Scholar]
- Faust, M.A. Micromorphology of a small dinoflagellate Prorocentrum mariae-lebouriae (Parke and Ballatine) comb. nov. J. Phycol. 1974, 10, 315–322. [Google Scholar] [CrossRef]
- Yang, H.; Hu, Z.; Tang, Y.Z. Plasticity and multiplicity of trophic modes in the dinoflagellate Karlodinium and their pertinence to population maintenance and bloom dynamics. Mar. Sci. Eng. 2021, 9, 51. [Google Scholar] [CrossRef]
- Pandeirada, M.S.; Craveiro, S.C.; Daugbjerg, N.; Moestrup, O.; Calado, A. Cell fine structure and phylogeny of Parvodinium: Towards an ultrastructural characterization of the Peridiniopsidaceae (Dinophyceae). Eur. J. Phycol. 2022, 58, 169–189. [Google Scholar] [CrossRef]
- Calado, A.; Craveiro, S.C.; Daugbjerg, N.; Moestrup, O. Ultrastructure and LSU rDNA-based phylogeny of Esoptrodinium gemma (Dinophyceae), with noted on feeding behaviour and the description of the flagellar base are of a planozygote. J. Phycol. 2006, 42, 434–452. [Google Scholar] [CrossRef]
- Tillmann, U.; Beran, A.; Gottschling, M.; Wietkamp, S.; Hoppenrath, M. Clarifying confusion—Prorocentrum triestinum J. Schiller and Prorocentrum redfieldii Bursa (Prorocentrales, Dinophyceae) are two different species. Eur. J. Phycol. 2022, 57, 207–226. [Google Scholar] [CrossRef]
- Mitra, A.; Flynn, K.J. Low rates of bacterivory enhances phototrophy and competitive advantage for mixoplankton growing in oligotrophic waters. Sci. Rep. 2023, 13, 6900. [Google Scholar] [CrossRef]
- Mitra, A.; Flynn, K.J.; Stoecker, D.K.; Raven, J.A. Trait trade-offs in phagotrophic microalgae: The mixoplankton conundrum. Eur. J. Phycol. 2023, 1–20. [Google Scholar] [CrossRef]
- Raven, J.A.; Beardall, J.; Flynn, K.J.; Maberly, S.C. Phagotrophy in the origins of photosynthesis in eukaryotes and as a complementary mode of nutrition in phototrophs: Relation to Darwin’s insectivorous plants. J. Exp. Bot. 2009, 60, 3975–3987. [Google Scholar] [CrossRef] [Green Version]
- Raven, J.A. Phagotrophy in phototrophs. Limnol. Oceanogr. 1997, 42, 198–205. [Google Scholar] [CrossRef]
- Flynn, K.J.; Hipkin, C.R. Interactions between iron, light, ammonium, and nitrate: Insights from the construction of a dynamic model of algal physiology. J. Phycol. 1999, 35, 1171–1190. [Google Scholar] [CrossRef]
- MacKenzie, L.; Sims, I.; Beuzenberg, V.; Gillespie, P. Mass accumulation of mucilage caused by dinoflagellate polysaccharide exudates in Tasman Bay, New Zealand. Harmful Algae 2002, 1, 69–83. [Google Scholar] [CrossRef]
- Honsell, G.; Cabrini, M.; Darin, M. Gonyaulax fragilis (Schütt) Kofoid: A dinoflagellate from gelatinous aggregates of the Northern Adriatic Sea. G. Bot. Ital. 1992, 126, 749–751. [Google Scholar] [CrossRef]
- Roux, P.; Siano, R.; Souchu, P.; Collin, K.; Schmitt, A.; Manach, S.; Retho, M.; Duplessix, O.P.; Marchand, L.; Colliec-Jouault, S.; et al. Spatio-temporal dynamics and biogeochemical properties of green seawater discolorations caused by the marine dinoflagellate Lepidodinium chlorophorum along southern Brittany coast. Estuar. Coast. Shelf Sci. 2022, 275, 107950. [Google Scholar] [CrossRef]
- Honsell, G.; Bonifacio, A.; De Bortoli, M.; Penna, A.; Battocchi, C.; Ciminiello, P.; Dell’Aversano, C.; Fattorusso, E.; Sosa, S.; Yasumoto, T.; et al. New insights on cytological and metabolic features of Ostreopsis cf. ovata Fukuyo (Dinophyceae): A multidisciplinary approach. PLoS ONE 2013, 8, e57291. [Google Scholar] [CrossRef] [PubMed]
- Taylor, J.F.R. The Biology of Dinoflagellates; Blackwell Scientific Publications: Oxford, UK, 1987; Volume 21, p. 785. [Google Scholar]
- Bouck, G.B.; Sweeney, B.M. The Fine Structure and Ontogeny of Trichocysts in Marine Dinoflagellates. Protoplasma 1966, 61, 205–223. [Google Scholar] [CrossRef] [PubMed]
- Tillmann, U.; Hoppenrath, M.; Gottschling, M. Reliable determination of Prorocentrum micans Ehrenb. (Prorocentrales, Dinophyceae) based on newly collected material from the type locality. Eur. J. Phycol. 2019, 54, 417–431. [Google Scholar] [CrossRef]
- Zhou, J.; Fritz, L. Ultrastructure of two toxic marine dinoflagellates, Prorocentrum lima and Prorocentrum maculosum. Phycologia 1993, 32, 444–450. [Google Scholar] [CrossRef]
- Hoppenrath, M.; Leander, B.S. Morphology and molecular phylogeny of a new marine sand-dwelling Prorocentrum species, P. tsawwassenense (Dinophyceae, Prorocentrales), from British Columbia, Canada. J. Phycol. 2008, 44, 451–455. [Google Scholar] [CrossRef]
- Dodge, J.D.; Bibby, B.T. The Prorocentrales (Dinophyceae) I. A comparative account of fine structure in the genera Prorocentrum and Exuviaella. Bot. J. Linn. Soc. 1973, 67, 175–187. [Google Scholar] [CrossRef]
- Dodge, J.D. The ultrastructure of the dinoflagellate pusule: A unique osmo.regulatory organelle. Protoplasma 1972, 75, 285–302. [Google Scholar] [CrossRef]
- Loeblich, A.R., III; Sherley, J.L.; Schmidt, R.J. The correct position of flagellar insertion in Prorocentrum and description of Prorocentrum rhathymum sp. nov. (Pyrrhophyta). J. Plankton Res. 1979, 1, 113–120. [Google Scholar] [CrossRef]
- Mohammad-Noor, N.; Moestrup, O.; Daugbjerg, N. Light, electron microscopy and DNA sequences of the dinoflagellate Prorocentrum concavum (syn. P. arabianum) with special emphasis on the periflagellar area. Phycologia 2007, 46, 549–564. [Google Scholar] [CrossRef]
- Murray, S.; Nagahama, Y.; Fukuyo, Y. Phylogenetic study of benthic, spine-bearing prorocentroids, including Prorocentrum fukuyoi sp. nov. Phycol. Res. 2007, 55, 91–102. [Google Scholar] [CrossRef]
- Faust, M.A.; Vandersea, M.W.; Kibler, S.; Tester, P.A.; Litaker, R.W. Prorcentrum levis, a new benthic species (Dinophyceae) from a mangrove island, Twin Case, Belize. J. Phycol. 2008, 44, 232–240. [Google Scholar] [CrossRef]
- Verma, A.; Kazandjan, A.; Sarowar, C.; Harwood, D.T.; Murray, J.S.; Pargmann, I.; Hoppenrath, M.; Murray, S.A. Morphology and phylogenetics of benthic Prorocentrum species (Dinophyceae) from tropical Northwestern Australia. Toxins 2021, 11, 571. [Google Scholar] [CrossRef] [Green Version]
- Cembella, A.D.; Durán-Riveroll, L.M.; Tarazona-Janampa, U.; Okolodkov, J.B.; Gracia-Sandoval, R.; Krock, B.; Hörstmann, C.; John, U. Phylogeography and diversity among populations of the toxigenic benthic dinoflagellate Prorocentrum from coastal reef systems in Mexico. Front. Mar. Sci. 2021, 8, 716669. [Google Scholar] [CrossRef]
- Hansen, P.J.; Tillmann, U. Mixotrophy in dinoflagellates: Prey selection, physiology and ecological importance. In Dinoflagellates: Classification, Evolution, Physiology, and Ecological Significance; Subba Rao, D.V., Ed.; Nova Science Publishers: New York, NY, USA, 2020; pp. 201–260. [Google Scholar]
- Kiorboe, T.; Titelman, J. Feeding, prey selection and prey encounter mechanisms in the heterotrophic dinoflagellate Noctiluca scintillans. J. Plankton Res. 1998, 20, 1615–1636. [Google Scholar] [CrossRef] [Green Version]
- Faust, M.A. Mixotrophy in tropical benthic dinoflagellates. In Harmful Algae, Proceedings of the VIII International Conference on Harmful Algae, Vigo, Spain, 25–29 June 1997; Reguera, B., Blanco, J., Fernandez, L., Wyatt, T., Eds.; Xunta de Galicia and Intergovernmental Oceanographic Commission of UNESCO: Paris, France, 1998; pp. 390–393. [Google Scholar]
- Faust, M.A.; Morton, S.L.; Quod, J.P. Further SEM studies of marine dinoflagellates: The genus Ostreopsis (Dinophyceae). J. Phycol. 1996, 32, 1053–1065. [Google Scholar] [CrossRef]
- Barone, R. Behavioral trait of Ostreopsis ovata (Dinophyceae) in Mediterranean rock pools: The spider’s strategy. Harmful Algae News 2007, 33, 1–3. [Google Scholar]
- Barone, R.; Prisinzano, A. Peculiarita comportamentale del dinoflagellato Ostreopsis ovata Fukuyo (Dinophyceae): La starategia del ragno. Nat. Sicil. 2006, 30, 401–418. [Google Scholar]
- Nagai, S.; Nitshitani, G.; Tomaru, Y.; Sakiyama, S.; Kamiyama, T. Predation by the toxic dinoflagellate Dinophysis fortii on the ciliate Myrionecta rubra and observation of sequestration of ciliate chloroplasts. J. Phycol. 2008, 44, 909–922. [Google Scholar] [CrossRef] [PubMed]
- Papiol, G.G.; Beuzenberg, V.; Selwood, A.I.; MacKenzie, L.; Packer, M.A. The use of a mucus trap by Dinophysis acuta for the capture of Mesodinium rubrum prey under culture conditions. Harmful Algae 2016, 58, 1–7. [Google Scholar] [CrossRef]
- Blossom, H.E.; Baedkel, T.D.; Tillmann, U.; Hansen, P.J. A search for mixotrophy and mucus trap production in Alexandrium spp. and the dynamics of mucus trap formation in Alexandrium pseudogonyaulax. Harmful Algae 2017, 64, 51–62. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Li, L.; Yu, L.; Yang, X.; Shi, X.; Wang, J.X.; Li, J.; Lin, S. Transcriptome profiling reveals verstile dissolved organic nitrogen utilization, mixotrophy, and N conservation in the dinoflagellate Prorocentrum shikokuense under N deficiency. Sci. Total Environ. 2021, 763, 143013. [Google Scholar] [CrossRef]
- Blossom, H.; Hansen, P.J. The loss of mixotrophy in Alexandrium pseudogonyaulax: Implications for trade-offs between toxicity, mucus trap production, and phagotrophy. Limnol. Oceanogr. 2021, 66, 528–542. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Strain | Reference | Isolator Date | Origin | Growth Conditions | ||||
|---|---|---|---|---|---|---|---|---|---|
| Temp | Sal | PFD | L:D | Medium | |||||
| P. pervagatum | PM-01 | [12] | U.Tillmann 2017 | Labrador Sea | 15 | 33 | 80 | 16:8 | 1/10 K |
| P. pervagatum | LP-D3 | [12] | U.Tillmann 2020 | North Sea, off Denmark | 20 | 33 | 80 | 16:8 | 1/10 K |
| P. pervagatum | LP-D10 | [12] | U.Tillmann 2020 | North Sea, off Denmark | 20 | 33 | 80 | 16:8 | 1/10 K |
| P. cordatum | BS 4-A5 | this study | U.Tillmann 2021 | southwestern Black Sea | 20 | 20 | 80 | 16:8 | 1/10 K |
| P. cordatum | BS 4-B5 | this study | U.Tillmann 2021 | southwestern Black Sea | 20 | 20 | 80 | 16:8 | 1/10 K |
| P. cordatum | BS 4-G2 | this study | U.Tillmann 2021 | southwestern Black Sea | 20 | 20 | 80 | 16:8 | 1/10 K |
| P. cordatum | DWER-PM23A8 | this study | M.Larsson 2021 | Wilson Inlet, Great Southern Region | 23 | 20 | 80 | 12:12 | K |
| P. cordatum | DWERP-M23H9 | this study | M.Larsson 2021 | Wilson Inlet, Great Southern Region | 23 | 20 | 80 | 12:12 | K |
| P. cordatum | DWER-PM19E6 | this study | M.Larsson 2021 | Murray River, Peel Region | 23 | 25 | 80 | 12:12 | K |
| P. cordatum | DWER-PM19F8 | this study | M.Larsson 2021 | Murray River, Peel Region | 23 | 25 | 80 | 12:12 | K |
| Teleaulax amphioxeia | k-1837 | NORCCA | n.a. | n.a. | 15/20 | 20/33 | 80 | 16:8 | 1/10 K |
| Rhodomonas salina | KAC-30 | KAC | n.a. | n.a. | 15/20 | 33 | 80 | 16:8 | 1/10 K |
| Rhodomomas salina | CS-24 | ANACC | n.a | n.a. | 23 | 20/25 | 80 | 12:12 | K |
| Parameter | Definition | Unit | Value |
|---|---|---|---|
| AE | assimilation efficiency of the predator | dimensionless | 0.8 a |
| Cappot | likelihood of a Prorocentrum cell (single or with mucus trap) capturing encountered prey | dimensionless | 0.2 a |
| Capmax | maximum prey cells that can be captured daily by a Prorocentrum cell (solitary or within a mucus trap) | prey predator−1 d−1 | 12 |
| ESDprey | equivalent spherical diameter of prey cell | µm | 8 * |
| equivalent spherical diameter of predator cell | µm | 13 * | |
| equivalent spherical diameter of trap | µm | 80–180 * | |
| swMot | Scalar for motility of predator calculated from swimming speeds | dimensionless | 0.7–1.3 * |
| µmax | maximum growth rate of Prorocentrum | gC (gC)−1 d−1 | 0.6 * |
| w | turbulence | m s−1 | 0–0.001 a |
| N | prey abundance | number m−3 | user selected |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tillmann, U.; Mitra, A.; Flynn, K.J.; Larsson, M.E. Mucus-Trap-Assisted Feeding Is a Common Strategy of the Small Mixoplanktonic Prorocentrum pervagatum and P. cordatum (Prorocentrales, Dinophyceae). Microorganisms 2023, 11, 1730. https://doi.org/10.3390/microorganisms11071730
Tillmann U, Mitra A, Flynn KJ, Larsson ME. Mucus-Trap-Assisted Feeding Is a Common Strategy of the Small Mixoplanktonic Prorocentrum pervagatum and P. cordatum (Prorocentrales, Dinophyceae). Microorganisms. 2023; 11(7):1730. https://doi.org/10.3390/microorganisms11071730
Chicago/Turabian StyleTillmann, Urban, Aditee Mitra, Kevin J. Flynn, and Michaela E. Larsson. 2023. "Mucus-Trap-Assisted Feeding Is a Common Strategy of the Small Mixoplanktonic Prorocentrum pervagatum and P. cordatum (Prorocentrales, Dinophyceae)" Microorganisms 11, no. 7: 1730. https://doi.org/10.3390/microorganisms11071730
APA StyleTillmann, U., Mitra, A., Flynn, K. J., & Larsson, M. E. (2023). Mucus-Trap-Assisted Feeding Is a Common Strategy of the Small Mixoplanktonic Prorocentrum pervagatum and P. cordatum (Prorocentrales, Dinophyceae). Microorganisms, 11(7), 1730. https://doi.org/10.3390/microorganisms11071730