
High Ammonium Addition Changes the Diversity and Structure of Bacterial Communities in Temperate Wetland Soils of Northeastern China
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Soil Sampling
2.3. Determination of Soil Physicochemical Properties
2.4. DNA Extraction
2.5. Bioinformatics
2.6. Statistical Analyses
3. Result
3.1. Changes of Soil Physicochemical Characteristics under Different NH4+ Addition
3.2. Sequencing Data Analysis and Distribution of Soil Bacterial Community
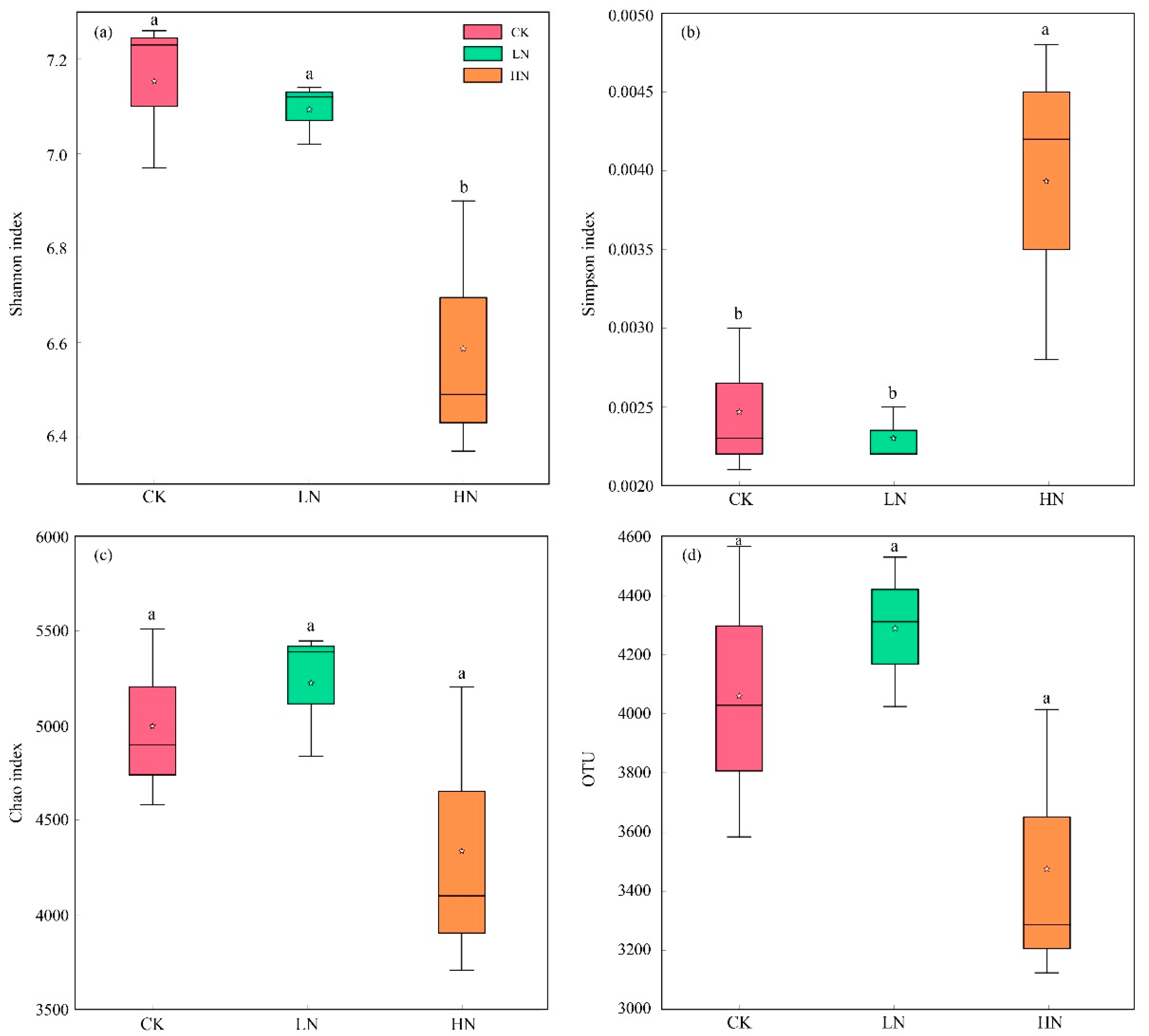
3.3. Increasing Ammonium Addition Influenced Bacterial α-Diversity
3.4. Soil Bacterial Community Composition Was Changed by Ammonium Addition
3.5. Indicator Analysis of Soil Bacterial Community Responding to Ammonium Addition
3.6. Ammonium Addition Changed the Bacterial Network Structure and Complexity
3.7. Relationships between Microbial Communities and Soil Properties
4. Discussion
4.1. Changes in Soil Bacterial Community Diversity and Structure
4.2. Effects of N Addition on the Soil Bacterial Community Composition
4.3. Changes in the Structure of Soil Bacterial Networks
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Galloway, J.N.; Dentener, F.J.; Capone, D.G.; Boyer, E.W.; Howarth, R.W.; Seitzinger, S.P.; Asner, G.P.; Cleveland, C.C.; Green, P.A.; Holland, E.A.; et al. Nitrogen cycles: Past, present, and future. Biogeochemistry 2004, 70, 153–226. [Google Scholar] [CrossRef]
- Hu, Y.; Chen, M.; Yang, Z.; Cong, M.; Zhu, X.; Jia, H. Soil microbial community response to nitrogen application on a swamp meadow in the arid region of central asia. Front. Microbiol. 2022, 12, 797306. [Google Scholar] [CrossRef] [PubMed]
- Zeng, J.; Liu, X.; Song, L.; Lin, X.; Zhang, H.; Shen, C.; Chu, H. Nitrogen fertilization directly affects soil bacterial diversity and indirectly affects bacterial community composition. Soil Biol. Biochem. 2016, 92, 41–49. [Google Scholar] [CrossRef]
- Knelman, J.E.; Nemergut, D.R. Changes in community assembly may shift the relationship between biodiversity and ecosystem function. Front. Microbiol. 2014, 5, 424. [Google Scholar] [CrossRef] [PubMed]
- Averill, C.; Waring, B. Nitrogen limitation of decomposition and decay: How can it occur? Glob. Chang. Biol. 2018, 24, 1417–1427. [Google Scholar] [CrossRef] [Green Version]
- Huang, L.B.; Bai, J.H.; Wen, X.J.; Zhang, G.L.; Zhang, C.D.; Cui, B.S.; Liu, X.H. Microbial resistance and resilience in response to environmental changes under the higher intensity of human activities than global average level. Glob. Chang. Biol. 2020, 26, 2377–2389. [Google Scholar] [CrossRef]
- Carreiro, M.M.; Sinsabaugh, R.L.; Repert, D.A.; Parkhurst, D.F. Microbial Enzyme Shifts Explain Litter Decay Responses to Simulated Nitrogen Deposition. Ecology 2000, 81, 2359. [Google Scholar] [CrossRef]
- Song, C.C.; Liu, D.Y.; Yang, G.S.; Song, Y.Y.; Mao, R. Effect of nitrogen addition on decomposition of Calamagrostis angustifolia litters from freshwater marshes of Northeast China. Ecol. Eng. 2011, 37, 1578–1582. [Google Scholar] [CrossRef]
- Song, C.C.; Wang, L.L.; Tian, H.Q.; Liu, D.Y.; Lu, C.Q.; Xu, X.F.; Zhang, L.H.; Yang, G.S.; Wan, Z.M. Effect of continued nitrogen enrichment on greenhouse gas emissions from a wetland ecosystem in the Sanjiang Plain, Northeast China: A 5-year nitrogen addition experiment. J. Geophys. Res. Biogeosci. 2013, 118, 741–751. [Google Scholar] [CrossRef]
- Bardgett, R.D.; Wardle, D.A. Aboveground-Belowground Linkages: Biotic interactions, Ecosystem Processes, and Global Change; Oxford University Press: New York, NY, USA, 2010. [Google Scholar]
- Peralta, R.M.; Ahn, C.; Gillevet, P.M. Characterization of soil bacterial community structure and physicochemical properties in created and natural wetlands. Sci. Total Environ. 2013, 443, 725–732. [Google Scholar] [CrossRef]
- Fierer, N.; Jackson, R.B. The diversity and biogeography of soil bacterial communities. Proc. Natl. Acad. Sci. USA 2006, 103, 626–631. [Google Scholar] [CrossRef] [PubMed]
- Li, B.B.; Roley, S.S.; Duncan, D.S.; Guo, J.; Quensen, J.F.; Yu, H.-Q.; Tiedje, J.M. Long-term excess nitrogen fertilizer increases sensitivity of soil microbial community to seasonal change revealed by ecological network and metagenome analyses. Soil Biol. Biochem. 2021, 160, 108349. [Google Scholar] [CrossRef]
- Xie, L.; Zhang, Q.; Cao, J.; Liu, X.; Xiong, D.; Kong, Q.; Yang, Y. Effects of Warming and Nitrogen Addition on the Soil Bacterial Community in a Subtropical Chinese Fir Plantation. Forests 2019, 10, 861. [Google Scholar] [CrossRef] [Green Version]
- Jia, M.; Gao, Z.; Gu, H.; Zhao, C.; Liu, M.; Liu, F.; Xie, L.; Wang, L.; Zhang, G.; Liu, Y.; et al. Effects of precipitation change and nitrogen addition on the composition, diversity, and molecular ecological network of soil bacterial communities in a desert steppe. PLoS ONE 2021, 16, e248194. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; Wang, T.; Shi, Z.; Chiariello, N.R.; Docherty, K.; Field, C.B.; Gutknecht, J.; Gao, Q.; Gu, Y.; Guo, X.; et al. Long-term nitrogen deposition enhances microbial capacities in soil carbon stabilization but reduces network complexity. Microbiome 2022, 10, 112. [Google Scholar] [CrossRef]
- Xu, L.; Zhang, B.; Peng, X.; Zhang, X.; Sun, B.; Sun, H.; Jiang, C.; Zhou, S.; Zeng, X.; Bai, Z.; et al. Dynamic variations of microbial community structure in Myriophyllum aquaticum constructed wetlands in response to different NH4+-N concentrations. Process Biochem. 2020, 93, 55–62. [Google Scholar] [CrossRef]
- Li, Y.; Tian, D.; Wang, J.; Niu, S.; Tian, J.; Ha, D.; Qu, Y.; Jing, G.; Kang, X.; Song, B. Differential mechanisms underlying responses of soil bacterial and fungal communities to nitrogen and phosphorus inputs in a subtropical forest. PeerJ 2019, 7, e7631. [Google Scholar] [CrossRef]
- Xu, R.H.; Tan, M.; Zhu, J.F.; Liu, Z.H. The response of soil microbial diversity to the difference of N deposition concentration in alpine wetland. J. Biol. 2021, 38, 75–81. [Google Scholar]
- Lu, G.; Xie, B.; Cagle, G.A.; Wang, X.; Han, G.; Wang, X.; Hou, A.; Guan, B. Effects of simulated nitrogen deposition on soil microbial community diversity in coastal wetland of the yellow river delta. Sci. Total Environ. 2021, 757, 143825. [Google Scholar] [CrossRef]
- Zhang, R.T.; Liu, Y.N.; Zhong, H.X.; Chen, X.W.; Sui, X. Effects of simulated nitrogen deposition on the soil microbial community diversity of a Deyeuxia angustifolia wetland in the Sanjiang plain, northeastern China. Ann. Microbiol. 2022, 72, 11. [Google Scholar] [CrossRef]
- Liu, X.J.; Zhang, Y.; Han, W.X.; Tang, A.H.; Shen, J.L.; Cui, Z.L.; Vitousek, P.; Erisman, J.W.; Goulding, K.; Christie, P.; et al. Enhanced nitrogen deposition over China. Nature 2013, 494, 459–462. [Google Scholar] [CrossRef] [PubMed]
- Fang, H.J.; Cheng, S.L.; Yu, G.R.; Zheng, J.J.; Zhang, P.L.; Xu, M.J.; Li, Y.N.; Yang, X.M. Responses of CO2 efflux from an alpine meadow soil on the Qinghai Tibetan Plateau to multi-form and low-level N addition. Plant Soil. 2012, 351, 177–190. [Google Scholar] [CrossRef]
- Fang, H.; Cheng, S.; Yu, G.; Cooch, J.; Wang, Y.; Xu, M.J.; Li, L.S.; Dang, X.S.; Li, Y.N. Low-level nitrogen deposition significantly inhibits methane uptake from an alpine meadow soil on the Qinghai-Tibetan Plateau. Geoderma 2014, 213, 444–452. [Google Scholar] [CrossRef]
- Kou, L.; Zhang, X.; Wang, H.; Yang, H.; Zhao, W.; Li, S. Nitrogen additions inhibit nitrification in acidic soils in a subtropical pine plantation: Effects of soil pH and compositional shifts in microbial groups. J. For. Res. 2018, 30, 669–678. [Google Scholar] [CrossRef]
- Matson, P.A.; McDowell, W.H.; Townsend, A.R.; Vitousek, P.M. The globalization of N deposition: Ecosystem consequences in tropical environments. Biogeochemistry 1999, 46, 67–83. [Google Scholar] [CrossRef]
- Recous, S.; Machet, J.M.; Mary, B. The partitioning of fertilizer-N between soil and crop-comparison of ammonium and nitrate applications. Plant Soil. 1992, 144, 101–111. [Google Scholar] [CrossRef]
- Zhang, Y.T.; Shen, H.; He, X.H.; Thomas, B.W.; Lupwayi, N.Z.; Hao, X.Y.; Thomas, M.C.; Shi, X.J. Fertilization shapes bacterial community structure by alteration of soil pH. Front. Microbiol. 2017, 8, 1325. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.; Dong, Y.; Sun, Q.; Jiao, R. Soil bacterial community response to short-term manipulation of the nitrogen deposition form and dose in a Chinese fir plantation in southern China. Water Air Soil Poll. 2016, 227, 447. [Google Scholar] [CrossRef]
- Geng, J.; Fang, H.; Cheng, S.; Pei, J. Effects of N deposition on the quality and quantity of soil organic matter in a boreal forest: Contrasting roles of ammonium and nitrate. Catena 2021, 198, 104996. [Google Scholar] [CrossRef]
- Weng, X.H.; Sui, X.; Liu, Y.N.; Yang, L.B.; Zhang, R. Effect of nitrogen addition on the carbon metabolism of soil microorganisms in a Calamagrostis angustifolia wetland of the Sanjiang Plain, northeastern China. Ann. Microbiol. 2022, 72, 18. [Google Scholar] [CrossRef]
- Lei, K.; Zhang, M.X. The wetland resources in China and the conservation advices. Wetl. Sci. 2005, 3, 81–86. [Google Scholar]
- He, D.M.; Ruan, H.H. Long term effect of land reclamation from lake on chemical composition of soil organic matter and its mineralization. PLoS ONE 2014, 9, e99251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Liu, J.; He, L. Responses of carbon dynamics to nitrogen deposition in typical freshwater wetland of Sanjiang Plain. J. Chem. 2014, 2014, 603948. [Google Scholar] [CrossRef] [Green Version]
- Wen, Z.; Xu, W.; Li, Q.; Han, M.J.; Tang, A.; Zhang, Y.; Luo, X.S.; Shen, J.L.; Wang, W.; Li, K.H.; et al. Changes of nitrogen deposition in China from 1980 to 2018. Environ. Int. 2020, 144, 106022. [Google Scholar] [CrossRef]
- Sui, X.; Zhang, R.T.; Liu, Y.N.; Xu, N.; Ni, H.W. Influence of simulation nitrogen deposition on soil microbial functional diversity of Calamagrostis angustifolia wetland in Sanjiang Plain. Acta. Agrestia Sin. 2016, 24, 1226–1233. [Google Scholar]
- Luan, Z.Q.; Deng, W.; Bai, J.H.; Zhu, B.G. Protection of Honghe national reserve wetland habitat. Res. Soil Water Conserv. 2003, 10, 154–157. [Google Scholar]
- Jones, D.L.; Willett, V.B. Experimental evaluation of methods to quantify dissolved organic nitrogen (DON) and dissolved organic carbon (DOC) in soil. Soil Biol. Biochem. 2005, 38, 991–999. [Google Scholar] [CrossRef]
- Lane, D. 16S/23S rRNA sequencing. In Nucleic Acid Techniques in Bacterial Systematics; Stackebrandt, E., Goodfellow, M., Eds.; John Wiley and Sons: New York, NY, USA, 1991. [Google Scholar]
- Parks, D.H.; Tyson, G.W.; Hugenholtz, P.; Beiko, R.G. STAMP: Statistical analysis of taxonomic and functional profiles. Bioinformatics 2014, 30, 3123–3124. [Google Scholar] [CrossRef] [Green Version]
- Harpole, W.S.; Tilman, D. Grassland species loss resulting from reduced niche dimension. Nature 2007, 446, 791–793. [Google Scholar] [CrossRef]
- Zhang, X.; Han, X. Nitrogen deposition alters soil chemical properties and bacterial communities in the inner mongolia grassland. J. Environ. Sci. 2012, 24, 1483–1491. [Google Scholar] [CrossRef]
- Weng, X.H.; Sui, X.; Li, M.S.; Zhang, R.; Yang, L.B. Effect of simulated nitrogen deposition on soil microbial carbon metabolism in Calamagrostis angustifolia wetland in Sanjiang Plain. Huan Jing Ke Xue 2022, 43, 4674–4683. [Google Scholar]
- Wang, C.; Liu, D.; Bai, E. Decreasing soil microbial diversity is associated with decreasing microbial biomass under nitrogen addition. Soil Biol. Biochem. 2018, 120, 126–133. [Google Scholar] [CrossRef]
- Li, Z.M.; Shen, J.P.; Zhang, L.M.; Liu, G.P.; Bai, P.M.; He, J.Z. Effect of stimulated nitrogen deposition on the bacterial community structure of semiarid temperate grassland. Environ. Sci. 2018, 39, 5665–5671. [Google Scholar]
- Frey, B.; Carnol, M.; Dharmarajah, A.; Brunner, I.; Schleppi, P. Only minor changes in the soil microbiome of a sub-alpine forest after 20 years of moderately increased nitrogen loads. Front. For. Glob. Chang. 2020, 3, 77. [Google Scholar] [CrossRef]
- Deng, J.J.; Zhou, Y.B.; Yin, Y.; Wei, Y.W.; Qin, S.J.; Zhu, W.X. Soil bacterial community structure characteristics in coniferous forests of Montane Regions of eastern Liaoning Province, China. Acta. Ecol. Sin. 2019, 39, 997–1008. [Google Scholar]
- Sui, X.; Zhang, R.; Frey, B.; Yang, L.; Liu, Y.; Ni, H.; Li, M.H. Soil physicochemical properties drive the variation in soil microbial communities along a forest successional series in a degraded wetland in northeastern China. Ecol. Evol. 2021, 11, 2194–2208. [Google Scholar] [CrossRef]
- Sha, M.; Xu, J.; Zheng, Z.; Fa, K. Enhanced atmospheric nitrogen deposition triggered little change in soil microbial diversity and structure in a desert ecosystem. Glob. Ecol. Conserv. 2021, 31, e01879. [Google Scholar] [CrossRef]
- Gavrichkova, O.; Kuzyakov, Y. Ammonium versus nitrate nutrition of Zea mays and Lupinus albus: Effect on root-derived CO2 efflux. Soil Biol. Biochem. 2008, 40, 2835–2842. [Google Scholar] [CrossRef]
- Thirukkumaran, C.M.; Parkinson, D. Microbial respiration, biomass, metabolic quotient and litter decomposition in a lodgepole pine forest floor amended with nitrogen and phosphorous fertilizers. Soil Biol. Biochem. 2000, 32, 59–66. [Google Scholar] [CrossRef]
- Tischner, R. Nitrate uptake and reduction in higher and lower plants. Plant Cell Environ. 2000, 23, 1005–1024. [Google Scholar] [CrossRef]
- Wang, C.Y.; Lv, Y.N.; Liu, X.Y.; Wang, L. Ecological effects of atmospheric nitrogen deposition on soil enzyme activity. J. For. Res. 2013, 24, 109–114. [Google Scholar] [CrossRef]
- Xie, F.; Ma, A.; Zhou, H.; Liang, Y.; Yin, J.; Ma, K.; Zhuang, X.L.; Zhuang, G.Q. Revealing fungal communities in alpine wetlands through species diversity, functional diversity and ecological network diversity. Microorganisms. 2020, 8, 632. [Google Scholar] [CrossRef] [PubMed]
- Fierer, N.; Bradford, M.A.; Jackson, R.B. Toward an ecological classification of soil bacteria. Ecology. 2007, 88, 1354–1364. [Google Scholar] [CrossRef]
- Fierer, N.; Lauber, C.L.; Ramirez, K.S.; Zaneveld, J.; Bradford, M.A.; Knight, R. Comparative metagenomic, phylogenetic and physiological analyses of soil microbial communities across nitrogen gradients. ISME J. 2012, 6, 1007–1017. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dai, Z.; Su, W.; Chen, H.; Barberan, A.; Zhao, H.; Yu, M.; Yu, L.; Brookes, P.C.; Schadt, C.W.; Chang, S.X.; et al. Long-term nitrogen fertilization decreases bacterial diversity and favors the growth of actinobacteria and proteobacteria in agro-ecosystems across the globe. Glob. Chang. Biol. 2018, 24, 3452–3461. [Google Scholar] [CrossRef] [PubMed]
- Yao, M.J.; Rui, J.P.; Li, J.B.; Dai, Y.M.; Bai, Y.F.; Heděnec, P.; Wang, J.M.; Zhang, S.H.; Pei, K.Q.; Liu, C.; et al. Rate-specific responses of prokaryotic diversity and structure to nitrogen deposition in the Leymus chinensis steppe. Soil Biol. Biochem. 2014, 79, 81–90. [Google Scholar] [CrossRef]
- Lauber, C.L.; Hamady, M.; Knight, R.; Fierer, N. Pyrosequencing-based assessment of soil pH as a predictor of soil bacterial community structure at the continental scale. Appl. Environ. Microb. 2009, 75, 5111–5120. [Google Scholar] [CrossRef] [Green Version]
- Lauber, C.L.; Strickland, M.S.; Bradford, M.A.; Fierer, N. The influence of soil properties on the structure of bacterial and fungal communities across land-use types. Soil Biol. Biochem. 2008, 40, 2407–2415. [Google Scholar] [CrossRef]
- Wei, Z.W.; Li, Y.Y.; Jiang, W.; Liao, X.R. Diversity of Acidobacteria in rhizosphere soils of common trees in Wuxi. Chin. J. Ecol. 2018, 37, 2649–2656. [Google Scholar]
- Rousk, J.; Bååth, E.; Brookes, P.C.; Lauber, C.L.; Lozupone, C.; Caporaso, J.G.; Knight, R.; Fierer, N. Soil bacterial and fungal communities across a pH gradient in an arable soil. ISME J. 2010, 4, 1340–1350. [Google Scholar] [CrossRef]
- Ramirez, K.S.; Lauber, C.L.; Knight, R.; Bradford, M.A.; Fierer, N. Consistent effects of nitrogen fertilization on soil bacterial communities in contrasting systems. Ecology 2010, 91, 3463–3470. [Google Scholar] [CrossRef]
- Zhang, C.; Zhang, X.Y.; Zou, H.T.; Kou, L.; Yang, Y.; Wen, X.F.; Li, S.G.; Wang, H.M.; Sun, X.M. Contrasting effects of ammonium and nitrate additions on the biomass of soil microbial communities and enzyme activities in subtropical China. Biogeosciences 2017, 14, 4815–4827. [Google Scholar] [CrossRef] [Green Version]
- Breuker, A.; Stadler, S.; Schippers, A. Microbial community analysis of deeply buried marine sediments of the New Jersey shallow shelf (IODP Expedition 313). FEMS Microbiol. Ecol. 2013, 85, 578–592. [Google Scholar] [CrossRef] [Green Version]
- Lovisa, B.R.; Philip, H.; Tyson, G.W.; Blackall, L.L. Filamentous Chloroflexi (green non-sulfur bacteria) are abundant in wastewater treatment processes with biological nutrient removal. Microbiology 2002, 148, 2309. [Google Scholar]
- Degens, B.P.; Schipper, L.A.; Sparling, G.P.; Vojvodic-Vukovic, M. Decreases in organic C reserves in soils can reduce the catabolic diversity of soil microbial communities. Soil. Biol. Biochem. 2000, 32, 189–196. [Google Scholar] [CrossRef]
- Zhang, N.; Wan, S.; Guo, J.; Han, G.; Gutknecht, J.; Schmid, B.; Yu, L.; Liu, W.; Bi, J.; Wang, Z.; et al. Precipitation modifies the effects of warming and nitrogen addition on soil microbial communities in northern Chinese grasslands. Soil Biol. Biochem. 2015, 89, 12–23. [Google Scholar] [CrossRef] [Green Version]
- Keeler, B.L.; Hobbie, S.E.; Kellogg, L.E. Effects of long-term nitrogen addition on microbial enzyme activity in eight forested and grassland sites: Implications for litter and soil organic matter decomposition. Ecosystems 2009, 12, 1–15. [Google Scholar] [CrossRef]
- Liu, W.X.; Zhang, Z.; Wan, S.Q. Predominant role of water in regulating soil and microbial respiration and their responses to climate change in a semiarid grassland. Glob. Chang. Biol. 2009, 15, 184–195. [Google Scholar] [CrossRef]
- Zak, D.R.; Kling, G.W. Microbial community composition and function across an arctic tundra landscape. Ecology 2006, 87, 1659–1670. [Google Scholar] [CrossRef] [PubMed]
- Anderson, C.R.; Hamonts, K.; Clough, T.J.; Condron, L.M. Biochar does not affect soil N-transformations or microbial community structure under ruminant urine patches but does alter relative proportions of nitrogen cycling bacteria. Agric. Ecosyst. Environ. 2014, 191, 63–72. [Google Scholar] [CrossRef]
- Du, S.Y.; Yu, M.; Liu, F.H.; Xiao, L.L.; Zhang, H.X.; Tao, J.; Gu, W.; Gu, J.Y.; Chen, X. Effect of facility management regimes on soil bacterial diversity and community structure. Chin. J. Eco-Agric. 2017, 25, 1615–1625. [Google Scholar]
- Fei, Y.C.; Ye, Y.Q.; Zheng, H.; Lu, J.; You, Y.F.; Huang, Y.; Chen, A.L.; Cao, G.Q. Effect of C/N ratio under the control of exogenous nitrogen on the bacterial community structure in litters of Cunninghamia lanceolate plantation under different undergrowth vegetation management measures. Acta Ecol. Sin. 2021, 41, 2011–2023. [Google Scholar]
- Yao, F.; Yang, S.; Wang, Z.; Wang, X.; Ye, J.; Wang, X.; DeBruyn, J.M.; Feng, X.; Jiang, Y.; Li, H. Microbial taxa distribution is associated with ecological trophic cascades along an elevation gradient. Front. Microbiol. 2017, 8, 2071. [Google Scholar] [CrossRef] [Green Version]
- Xu, R.Y.; Zuo, M.X.; Yuan, Y.L.; Sun, J.; Lu, Y.S.; Jie, K.Z.; Xu, P.Z. Effects of nitrogen fertilizer dosage optimization on nitrogen uptake content and utilization efficiency and microbial function genes of nitrogen cycle in sweet corn. J. South. Agr. 2020, 51, 2919–2926. [Google Scholar]
- Polcyn, W.; Luciński, R. Aerobic and anaerobic nitrate and nitrite reduction in free-living cells of Bradyrhizobium sp. (Lupinus). FEMS Microbiol. Lett. 2003, 226, 331–337. [Google Scholar] [CrossRef] [Green Version]
- Rivas, R.; Martens, M.; Lajudie, P.; De Willems, A. Multilocus sequence analysis of the genus Bradyrhizobium. Syst. Appl. Microbiol. 2009, 32, 101–110. [Google Scholar] [CrossRef]
- Chen, C.L.; Wu, M.N.; Wei, W.X. Effect of long-term application of nitrogen fertilizer on the diversity of nitrifying genes (amoA and hao) in Paddy soil. Environ. Sci. 2011, 32, 1489–1496. [Google Scholar]
- Khan, A.; Jiang, H.; Bu, J.; Adnan, M.; Gilanni, S.W.; Hussain, M.A.; Zhang, M.Q. Untangling the Rhizosphere Bacterial Community Composition and Response of Soil Physiochemical Properties to Different Nitrogen Applications in Sugarcane Field. Fron. Microbiol. 2022, 13, 856078. [Google Scholar] [CrossRef]
- Wan, Y.; Li, W.; Wang, J.; Shi, X. Bacterial diversity and community in response to long-term nitrogen fertilization gradient in citrus orchard soils. Diversity 2021, 13, 282. [Google Scholar] [CrossRef]
- Shen, C.; Ni, Y.; Liang, W.; Wang, J.; Chu, H. Distinct soil bacterial communities along a small-scale elevational gradient in alpine tundra. Front. Microbiol. 2015, 6, 582. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Dong, S.K.; Gao, Q.Z.; Liu, S.L.; Zhou, H.K.; Ganjurjav, H.; Wang, X.X. Climate change and human activities altered the diversity and composition of soil microbial community in alpine grasslands of the Qinghai-Tibetan Plateau. Sci. Total Environ. 2016, 562, 353–363. [Google Scholar] [CrossRef] [PubMed]
- Montoya, J.M.; Pimm, S.L.; Solé, R.V. Ecological networks and their fragility. Nature 2006, 442, 259–264. [Google Scholar] [CrossRef]
- Helbing, D. Globally networked risks and how to respond. Nature 2013, 497, 51–59. [Google Scholar] [CrossRef]
- Yuan, M.M.; Guo, X.; Wu, L.W.; Zhang, Y.; Xiao, N.J.; Ning, D.L.; Shi, Z.; Zhou, X.S.; Wu, L.Y.; Yang, Y.F. Climate warming enhances microbial network complexity and stability. Nat. Clim. Chang. 2021, 11, 343–348. [Google Scholar] [CrossRef]
- Han, X. Effects of Soil Nitrogen to Plant—Microbial on Nitrogen Competition in Temperate Forest; Beijing Forestry University: Beijing, China, 2015. [Google Scholar]
- Coyte, K.Z.; Schluter, J.; Foster, K.R. The ecology of the microbiome: Networks, competition, and stability. Science 2015, 350, 663–666. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.; Gao, Y.; Jia, X.; Wang, M.; Ding, J.; Cheng, L.; Bao, F.; Wu, B. Network analysis reveals the strengthening of microbial interaction in biological soil crust development in the Mu Us Sandy Land, northwestern China. Soil Biol. Biochem. 2020, 144, 107782. [Google Scholar] [CrossRef]
- Yu, Z.; Hu, X.; Wei, D.; Liu, J.; Zhou, B.; Jin, J.; Liu, X.; Wang, G. Long-term inorganic fertilizer use influences bacterial communities in mollisols of northeast China based on high-throughput sequencing and network analyses. Arch. Agron. Soil Sci. 2019, 65, 1331–1340. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatments | SMC | pH | NO3 (mg/kg) | NH4+ (mg/kg) | DOC (mg/kg) | DON (mg/kg) | TN (%) | SOC (%) |
|---|---|---|---|---|---|---|---|---|
| CK | 0.43 ± 0.06 b | 4.41 ± 0.43 a | 23.83 ± 6.34 b | 21.15 ± 3.45 a | 3897.88 ± 625.37 b | 299.74 ± 130.71 b | 0.57 ± 0.23 b | 5.34 ± 2.23 a |
| LN | 0.48 ± 0.01 ab | 4.19 ± 0.06 ab | 23.27 ± 3.49 b | 5.62 ± 0.63 b | 2571.62 ± 294.54 c | 520.08 ± 166.01 ab | 0.68 ± 0.05 ab | 4.53 ± 0.42 a |
| HN | 0.58 ± 0.05 a | 4.02 ± 0.11 b | 40.60 ± 6.17 a | 4.72 ± 0.06 b | 5004.81 ± 331.11 a | 719.68 ± 156.87 a | 1.18 ± 0.29 a | 6.86 ± 2.00 a |
| Treatment | OTU Numbers | Total Links | Positive Link | Negative Link | Average Degree | Average Clustering Coefficient | Average Geodesic Distance | Network Density | Modularity (No. of Modules) |
|---|---|---|---|---|---|---|---|---|---|
| CK | 465 | 11,902 | 6637 | 5265 | 25.6 | 0.784 | 11.723 | 0.055 | 0.812 (11) |
| LN | 464 | 10,922 | 5809 | 5113 | 23.5 | 0.791 | 11.604 | 0.051 | 0.824 (10) |
| HN | 458 | 12,542 | 7329 | 5213 | 27.4 | 0.812 | 11.409 | 0.060 | 0.779 (9) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Weng, X.; Wang, M.; Sui, X.; Frey, B.; Liu, Y.; Zhang, R.; Ni, H.; Li, M. High Ammonium Addition Changes the Diversity and Structure of Bacterial Communities in Temperate Wetland Soils of Northeastern China. Microorganisms 2023, 11, 2033. https://doi.org/10.3390/microorganisms11082033
Weng X, Wang M, Sui X, Frey B, Liu Y, Zhang R, Ni H, Li M. High Ammonium Addition Changes the Diversity and Structure of Bacterial Communities in Temperate Wetland Soils of Northeastern China. Microorganisms. 2023; 11(8):2033. https://doi.org/10.3390/microorganisms11082033
Chicago/Turabian StyleWeng, Xiaohong, Mingyu Wang, Xin Sui, Beat Frey, Yingnan Liu, Rongtao Zhang, Hongwei Ni, and Maihe Li. 2023. "High Ammonium Addition Changes the Diversity and Structure of Bacterial Communities in Temperate Wetland Soils of Northeastern China" Microorganisms 11, no. 8: 2033. https://doi.org/10.3390/microorganisms11082033
APA StyleWeng, X., Wang, M., Sui, X., Frey, B., Liu, Y., Zhang, R., Ni, H., & Li, M. (2023). High Ammonium Addition Changes the Diversity and Structure of Bacterial Communities in Temperate Wetland Soils of Northeastern China. Microorganisms, 11(8), 2033. https://doi.org/10.3390/microorganisms11082033