
Interaction of Naturally Occurring Phytoplankton with the Biogeochemical Cycling of Mercury in Aquatic Environments and Its Effects on Global Hg Pollution and Public Health

 ,
,  , , and
, , and 
Abstract
:1. Introduction
2. Mercury Forms in the Environment
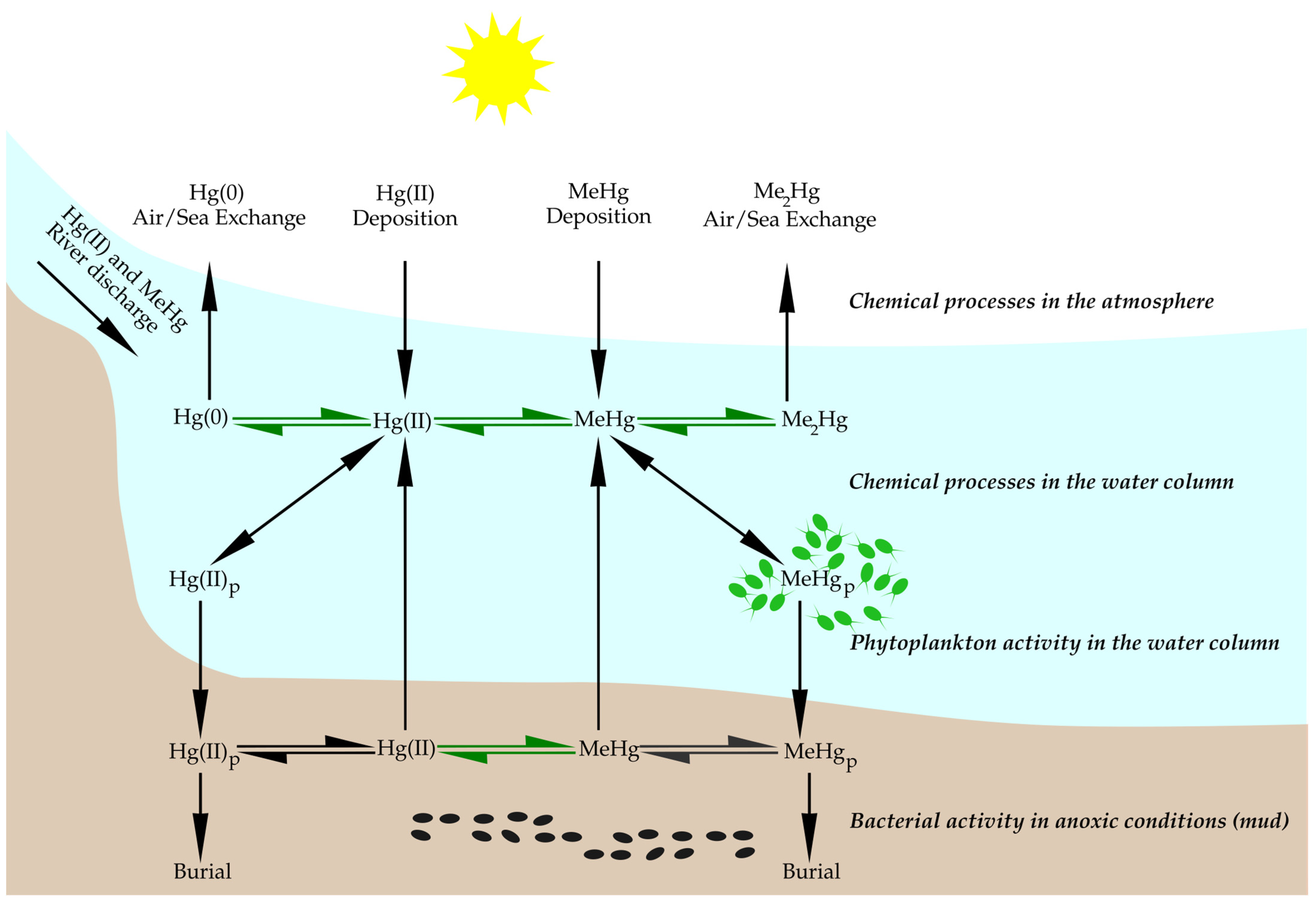
3. Biogeochemical Cycling of Mercury and Methylmercury
4. Impact of Anthropogenic Climate Change on Biogeochemical Cycling of Mercury
5. Mercury Bioaccumulation in Aquatic Food Chains
6. Effects of Mercury Exposure on Phytoplankton
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Bravo, A.G.; Cosio, C. Biotic formation of methylmercury: A bio-physico-chemical conundrum. Limnol. Oceanogr. 2019, 65, 1010–1027. [Google Scholar] [CrossRef]
- Karimi, R.; Fitzgerald, T.P.; Fisher, N.S. A Quantitative Synthesis of Mercury in Commercial Seafood and Implications for Exposure in the United States. Environ. Health Perspect. 2012, 120, 1512–1519. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Monastero, R.N.; Karimi, R.; Nyland, J.F.; Harrington, J.; Levine, K.; Meliker, J.R. Mercury exposure, serum antinuclear antibodies, and serum cytokine levels in the Long Island Study of Seafood Consumption: A cross-sectional study in NY, USA. Environ. Res. 2017, 156, 334–340. [Google Scholar] [CrossRef] [PubMed]
- Nyholt, K.; Jardine, T.D.; Villamarín, F.; Jacobi, C.M.; Hawes, J.E.; Campos-Silva, J.V.; Srayko, S.; Magnusson, W.E. High rates of mercury biomagnification in fish from Amazonian floodplain-lake food webs. Sci. Total Environ. 2022, 833, 155161. [Google Scholar] [CrossRef] [PubMed]
- Seco, J.; Aparício, S.; Brierley, A.S.; Bustamante, P.; Ceia, F.R.; Coelho, J.P.; Philips, R.A.; Saunders, R.A.; Fielding, S.; Gregory, S.; et al. Mercury biomagnification in a Southern Ocean food web. Environ. Pollut. 2021, 275, 116620. [Google Scholar] [CrossRef]
- Azevedo, L.S.; Pestana, I.A.; Almeida, M.G.; Nery, A.F.D.C.; Bastos, W.R.; Souza, C.M.M. Mercury biomagnification in an ichthyic food chain of an amazon floodplain lake (Puruzinho Lake): Influence of seasonality and food chain modeling. Ecotoxicol. Environ. Saf. 2021, 207, 111249. [Google Scholar] [CrossRef] [PubMed]
- Cossart, T.; Garcia-Calleja, J.; Santos, J.P.; Kalahroodi, E.L.; Worms, I.A.M.; Pedrero, Z.; Amouroux, D.; Slaveykova, V.I. Role of phytoplankton in aquatic mercury speciation and transformations. Environ. Chem. 2022, 19, 104–115. [Google Scholar] [CrossRef]
- Wu, P.; Zakem, E.J.; Dutkiewicz, S.; Zhang, Y. Biomagnification of Methylmercury in a Marine Plankton Ecosystem. Environ. Sci. Technol. 2020, 54, 5446–5455. [Google Scholar] [CrossRef] [PubMed]
- Beauvais-Flück, R.; Slaveykova, V.I.; Cosio, C. Molecular Effects of Inorganic and Methyl Mercury in Aquatic Primary Producers: Comparing Impact to A Macrophyte and A Green Microalga in Controlled Conditions. Geosciences 2018, 8, 393. [Google Scholar] [CrossRef] [Green Version]
- Beauvais-Flück, R.; Slaveykova, V.I.; Cosio, C. Cellular toxicity pathways of inorganic and methyl mercury in the green microalga Chlamydomonas reinhardtii. Sci. Rep. 2017, 7, 8034. [Google Scholar] [CrossRef]
- Santos, J.P.; Mehmeti, L.; Slaveykova, V.I. Simple Acid Digestion Procedure for the Determination of Total Mercury in Plankton by Cold Vapor Atomic Fluorescence Spectroscopy. Methods Protoc. 2022, 5, 29. [Google Scholar] [CrossRef]
- Mason, R.P.; Choi, A.L.; Fitzgerald, W.F.; Hammerschmidt, C.R.; Lamborg, C.H.; Soerensen, A.L.; Sunderland, E.M. Mercury biogeochemical cycling in the ocean and policy implications. Environ. Res. 2012, 119, 101–117. [Google Scholar] [CrossRef] [Green Version]
- Pinto, E.P.; Paredes, E.; Bellas, J. Influence of microplastics on the toxicity of chlorpyrifos and mercury on the marine microalgae Rhodomonas lens. Sci. Total Environ. 2023, 857, 159605. [Google Scholar] [CrossRef]
- Winder, M.; Sommer, U. Phytoplankton response to a changing climate. Hydrobiologia 2012, 698, 5–16. [Google Scholar] [CrossRef]
- B-Béres, V.; Stenger-Kovács, C.; Buczkó, K.; Padisák, J.; Selmeczy, G.B.; Lengyel, E.; Tapolczai, K. Ecosystem services provided by freshwater and marine diatoms. Hydrobiologia 2023, 850, 2707–2733. [Google Scholar] [CrossRef]
- Boening, D.W. Ecological effects, transport, and fate of mercury: A general review. Chemosphere 2000, 40, 1335–1351. [Google Scholar] [CrossRef]
- Thera, J.C.; Kidd, K.A.; Stewart, A.R.; Bertolo, R.F.; O’Driscoll, N.J. Using tissue cysteine to predict the trophic transfer of methylmercury and selenium in lake food webs. Environ. Pollut. 2022, 311, 119936. [Google Scholar] [CrossRef]
- Wiklund, J.A.; Kirk, J.L.; Muir, D.C.; Evans, M.; Yang, F.; Keating, J.; Parsons, M.T. Anthropogenic mercury deposition in Flin Flon Manitoba and the Experimental Lakes Area Ontario (Canada): A multi-lake sediment core reconstruction. Sci. Total Environ. 2017, 586, 685–695. [Google Scholar] [CrossRef]
- Ullrich, S.M.; Tanton, T.W.; Abdrashitova, S.A. Mercury in the Aquatic Environment: A Review of Factors Affecting Methylation. Crit. Rev. Environ. Sci. Technol. 2001, 31, 241–293. [Google Scholar] [CrossRef]
- Eklöf, K.; Bishop, K.; Bertilsson, S.; Björn, E.; Buck, M.; Skyllberg, U.; Osman, O.A.; Kronberg, R.-M.; Bravo, A.G. Formation of mercury methylation hotspots as a consequence of forestry operations. Sci. Total Environ. 2018, 613–614, 1069–1078. [Google Scholar] [CrossRef]
- Gworek, B.; Dmuchowski, W.; Baczewska-Dąbrowska, A.H. Mercury in the terrestrial environment: A review. Environ. Sci. Eur. 2020, 32, 128. [Google Scholar] [CrossRef]
- Mahboob, S.; Al-Ghanim, K.A.; Al-Misned, F.; Shahid, T.; Sultana, S.; Sultan, T.; Hussain, B.; Ahmed, Z. Impact of Water Pollution on Trophic Transfer of Fatty Acids in Fish, Microalgae, and Zoobenthos in the Food Web of a Freshwater Ecosystem. Biomolecules 2019, 9, 231. [Google Scholar] [CrossRef] [Green Version]
- Lavoie, R.A.; Jardine, T.D.; Chumchal, M.M.; Kidd, K.A.; Campbell, L.M. Biomagnification of Mercury in Aquatic Food Webs: A Worldwide Meta-Analysis. Environ. Sci. Technol. 2013, 47, 13385–13394. [Google Scholar] [CrossRef]
- Marnane, I. Mercury, a Persistent Threat to the Environment and Health. European Environment Agency, 2018. Available online: https://www.eea.europa.eu/articles/mercury-a-persistent-threat-to (accessed on 25 June 2023).
- Li, J.; Chen, B.; Chen, G.; Wei, W.; Wang, X.; Ge, J.; Dong, K.; Xia, H.; Xia, X. Tracking mercury emission flows in the global supply chains: A multi-regional input-output analysis. J. Clean. Prod. 2017, 140, 1470–1492. [Google Scholar] [CrossRef]
- Zhang, H.; Zhou, Y.; Liu, T.; Tian, X.; Zhang, Y.; Wang, J.; Zhang, M.; Phoutthavong, T.; Liang, P. Mercury release behaviors of Guizhou bituminous coal during co-pyrolysis: Influence of Chlorella. J. Environ. Sci. 2022, 119, 23–32. [Google Scholar] [CrossRef]
- Dastoor, A.; Angot, H.; Bieser, J.; Christensen, J.H.; Douglas, T.A.; Heimbürger-Boavida, L.-E.; Jiskra, M.; Mason, R.P.; McLagan, D.S.; Obrist, D.; et al. Arctic mercury cycling. Nat. Rev. Earth Environ. 2022, 3, 270–286. [Google Scholar] [CrossRef]
- Guédron, S.; Tolu, J.; Brisset, E.; Sabatier, P.; Perrot, V.; Bouchet, S.; Develle, A.; Bindler, R.; Cossa, D.; Fritz, S.; et al. Late Holocene volcanic and anthropogenic mercury deposition in the western Central Andes (Lake Chungará, Chile). Sci. Total Environ. 2019, 662, 903–914. [Google Scholar] [CrossRef]
- Li, Y.; Li, D.; Song, B.; Li, Y. The potential of mercury methylation and demethylation by 15 species of marine microalgae. Water Res. 2022, 215, 118266. [Google Scholar] [CrossRef]
- Siedlewicz, G.; Korejwo, E.; Szubska, M.; Grabowski, M.; Kwasigroch, U.; Bełdowski, J. Presence of mercury and methylmercury in Baltic Sea sediments, collected in ammunition dumpsites. Mar. Environ. Res. 2020, 162, 105158. [Google Scholar] [CrossRef]
- Capo, E.; Feng, C.; Bravo, A.G.; Bertilsson, S.; Soerensen, A.L.; Pinhassi, J.; Buck, M.; Karlsson, C.; Hawkes, J.; Björn, E. Expression Levels of hgcAB Genes and Mercury Availability Jointly Explain Methylmercury Formation in Stratified Brackish Waters. Environ. Sci. Technol. 2022, 56, 13119–13130. [Google Scholar] [CrossRef]
- Capo, E.; Broman, E.; Bonaglia, S.; Bravo, A.G.; Bertilsson, S.; Soerensen, A.L.; Pinhassi, J.; Lundin, D.; Buck, M.; Hall, P.O.J.; et al. Oxygen-deficient water zones in the Baltic Sea promote uncharacterized Hg methylating microorganisms in underlying sediments. Limnol. Oceanogr. 2021, 67, 135–146. [Google Scholar] [CrossRef]
- Li, P.; Wang, R.; Kainz, M.J.; Yin, D. Algal Density Controls the Spatial Variations in Hg Bioconcentration and Bioaccumulation at the Base of the Pelagic Food Web of Lake Taihu, China. Environ. Sci. Technol. 2022, 56, 14528–14538. [Google Scholar] [CrossRef]
- Quiroga-Flores, R.; Guédron, S.; Achá, D. High methylmercury uptake by green algae in Lake Titicaca: Potential implications for remediation. Ecotoxicol. Environ. Saf. 2021, 207, 111256. [Google Scholar] [CrossRef]
- Ulus, Y.; Tsui, M.T.-K.; Sakar, A.; Nyarko, P.; Aitmbarek, N.B.; Ardón, M.; Chow, A.T. Declines of methylmercury along a salinity gradient in a low-lying coastal wetland ecosystem at South Carolina, USA. Chemosphere 2022, 308, 136310. [Google Scholar] [CrossRef]
- Lanza, W.G.; Achá, D.; Point, D.; Masbou, J.; Alanoca, L.; Amouroux, D.; Lazzaro, X. Association of a Specific Algal Group with Methylmercury Accumulation in Periphyton of a Tropical High-Altitude Andean Lake. Arch. Environ. Contam. Toxicol. 2017, 72, 1–10. [Google Scholar] [CrossRef]
- Soerensen, A.L.; Schartup, A.T.; Gustafsson, E.; Gustafsson, B.G.; Undeman, E.; Björn, E. Eutrophication Increases Phytoplankton Methylmercury Concentrations in a Coastal Sea—A Baltic Sea Case Study. Environ. Sci. Technol. 2016, 50, 11787–11796. [Google Scholar] [CrossRef] [Green Version]
- Krabbenhoft, D.P. (Ed.) Methylmercury Contamination of Aquatic Ecosystems: A Widespread Problem with Many Challenges for the Chemical Sciences. In Water and Sustainable Development: Opportunities for the Chemical Sciences: A Workshop Report to the Chemical Sciences Roundtable; National Academy of Sciences: Washington, DC, USA, 2004. [Google Scholar]
- Sams, C.E. Methylmercury Contamination: Impacts on Aquatic Systems and Terrestrial Species, and Insights for Abatement. In Advancing the Fundamental Sciences, Proceedings of the Forest Service National Earth Sciences Conference, San Diego, CA, USA, 18–22 October 2004; U.S. Department of Agriculture, Forest Service, Pacific Northwest Research Station: San Diego, CA, USA, 2007. [Google Scholar]
- Selin, N.E. Global Biogeochemical Cycling of Mercury: A Review. An. Rev. Environ. Resour. 2009, 34, 43–63. [Google Scholar] [CrossRef] [Green Version]
- Canário, J.; Santos-Echeandia, J.; Padeiro, A.; Amaro, E.; Strass, V.; Klaas, C.; Hoppema, M.; Ossebaar, S.; Koch, B.P.; Laglera, L.M. Mercury and methylmercury in the Atlantic sector of the Southern Ocean. Deep. Sea Res. Part II Top. Stud. Oceanogr. 2017, 138, 52–62. [Google Scholar] [CrossRef] [Green Version]
- Caballero-Gallardo, K.; Palomares-Bolaños, J.; Olivero-Verbel, J. Mercury Concentrations in Water, Sediments, Soil, and Fish Around Ancestral Afro-Descendant Territories Impacted by Gold Mining in the Cauca Department, Colombia. Water Air Soil Pollut. 2022, 233, 393. [Google Scholar] [CrossRef]
- Stoichev, T.; de Chanvalon, A.T.; Veloso, S.; Deborde, J.; Tessier, E.; Lanceleur, L.; Amouroux, D. Assessing and predicting the changes for inorganic mercury and methylmercury concentrations in surface waters of a tidal estuary (Adour Estuary, SW France). Mar. Pollut. Bull. 2023, 186, 114400. [Google Scholar] [CrossRef]
- Horvat, M.; Kotnik, J.; Logar, M.; Fajon, V.; Zvonarić, T.; Pirrone, N. Speciation of mercury in surface and deep-sea waters in the Mediterranean Sea. Atmos. Environ. 2003, 37, 93–108. [Google Scholar] [CrossRef]
- Roulet, M.; Lucotte, M.; Canuel, R.; Farella, N.; De Freitos Goch, Y.G.; Peleja, J.R.P.; Guimaraes, J.R.D.; Mergler, D.; De Amorim, M.M.R. Spatio-temporal geochemistry of mercury in waters of the Tapajós and Amazon rivers, Brazil. Limnol. Oceanogr. 2001, 46, 1141–1157. [Google Scholar] [CrossRef] [Green Version]
- Cui, X.; Lamborg, C.H.; Hammerschmidt, C.R.; Xiang, Y.; Lam, P.J. The Effect of Particle Composition and Concentration on the Partitioning Coefficient for Mercury in Three Ocean Basins. Front. Environ. Chem. 2021, 2, 660267. [Google Scholar] [CrossRef]
- Mason, R.; Rolfhus, K.; Fitzgerald, W. Mercury in the North Atlantic. Mar. Chem. 1998, 61, 37–53. [Google Scholar] [CrossRef]
- Lescord, G.L.; Emilson, E.J.S.; Johnston, T.A.; Branfireun, B.A.; Gunn, J.M. Optical Properties of Dissolved Organic Matter and Their Relation to Mercury Concentrations in Water and Biota Across a Remote Freshwater Drainage Basin. Environ. Sci. Technol. 2018, 52, 3344–3353. [Google Scholar] [CrossRef]
- Lamborg, C.H.; Fitzgerald, W.F.; Damman, A.W.H.; Benoit, J.M.; Balcom, P.H.; Engstrom, D.R. Modern and historic atmospheric mercury fluxes in both hemispheres: Global and regional mercury cycling implications. Glob. Biogeochem. Cycles 2002, 16, 51-1–51-11. [Google Scholar] [CrossRef]
- Soerensen, A.L.; Schartup, A.T.; Skrobonja, A.; Bouchet, S.; Amouroux, D.; Liem-Nguyen, V.; Björn, E. Deciphering the Role of Water Column Redoxclines on Methylmercury Cycling Using Speciation Modeling and Observations From the Baltic Sea. Glob. Biogeochem. Cycles 2018, 32, 1498–1513. [Google Scholar] [CrossRef]
- Soerensen, A.; Schartup, A.; Skrobonja, A.; Björn, E. Organic matter drives high interannual variability in methylmercury concentrations in a subarctic coastal sea. Environ. Pollut. 2017, 229, 531–538. [Google Scholar] [CrossRef]
- Rodríguez, J.; Andersson, A.; Björn, E.; Timonen, S.; Brugel, S.; Skrobonja, A.; Rowe, O. Inputs of Terrestrial Dissolved Organic Matter Enhance Bacterial Production and Methylmercury Formation in Oxic Coastal Water. Front. Microbiol. 2022, 13, 809166. [Google Scholar] [CrossRef]
- Morway, E.D.; Thodal, C.E.; Marvin-DiPasquale, M. Long-term trends of surface-water mercury and methylmercury concentrations downstream of historic mining within the Carson River watershed. Environ. Pollut. 2017, 229, 1006–1018. [Google Scholar] [CrossRef]
- Shadrin, N.; Stetsiuk, A.; Anufriieva, E. Differences in Mercury Concentrations in Water and Hydrobionts of the Crimean Saline Lakes: Does Only Salinity Matter. Water 2022, 14, 2613. [Google Scholar] [CrossRef]
- Kannan, K.; Smith, R.G.S., Jr.; Lee, R.F.; Windom, H.L.; Heitmuller, P.T.; Macauley, J.M.; Summers, J.K. Distribution of Total Mercury and Methyl Mercury in Water, Sediment, and Fish from South Florida Estuaries. Arch. Environ. Contam. Toxicol. 1998, 34, 109–118. [Google Scholar] [CrossRef]
- Mao, L.; Liu, X.; Wang, B.; Lin, C.; Xin, M.; Zhang, B.-T.; Wu, T.; He, M.; Ouyang, W. Occurrence and risk assessment of total mercury and methylmercury in surface seawater and sediments from the Jiaozhou Bay, Yellow Sea. Sci. Total Environ. 2020, 714, 136539. [Google Scholar] [CrossRef]
- Emmerton, C.A.; Cooke, C.A.; Wentworth, G.R.; Graydon, J.A.; Ryjkov, A.; Dastoor, A. Total Mercury and Methylmercury in Lake Water of Canada’s Oil Sands Region. Environ. Sci. Technol. 2018, 52, 10946–10955. [Google Scholar] [CrossRef] [Green Version]
- Achá, D.; Guédron, S.; Amouroux, D.; Point, D.; Lazzaro, X.; Fernandez, P.E.; Sarret, G. Algal Bloom Exacerbates Hydrogen Sulfide and Methylmercury Contamination in the Emblematic High-Altitude Lake Titicaca. Geosciences 2018, 8, 438. [Google Scholar] [CrossRef] [Green Version]
- Ramlal, P.S.; Bugenyi, F.W.; Kling, G.W.; Nriagu, J.O.; Rudd, J.W.; Campbell, L.M. Mercury Concentrations in Water, Sediment, and Biota from Lake Victoria, East Africa. J. Great Lakes Res. 2003, 29, 283–291. [Google Scholar] [CrossRef]
- Cinnirella, S.; Bruno, D.E.; Pirrone, N.; Horvat, M.; Živković, I.; Evers, D.C.; Johnson, S.; Sunderland, E.M. Mercury concentrations in biota in the Mediterranean Sea, a compilation of 40 years of surveys. Sci. Data 2019, 6, 205. [Google Scholar] [CrossRef] [Green Version]
- Cossa, D.; Martin, J.-M.; Takayanagi, K.; Sanjuan, J. The distribution and cycling of mercury species in the western Mediterranean. Deep. Sea Res. Part II Top. Stud. Oceanogr. 1997, 44, 721–740. [Google Scholar] [CrossRef]
- Guédron, S.; Tisserand, D.; Garambois, S.; Spadini, L.; Molton, F.; Bounvilay, B.; Charlet, L.; Polya, D.A. Baseline investigation of (methyl)mercury in waters, soils, sediments and key foodstuffs in the Lower Mekong Basin: The rapidly developing city of Vientiane (Lao PDR). J. Geochem. Explor. 2014, 143, 96–102. [Google Scholar] [CrossRef]
- Bravo, A.G.; Le Faucheur, S.; Monperrus, M.; Amouroux, D.; Slaveykova, V.I. Species-specific isotope tracers to study the accumulation and biotransformation of mixtures of inorganic and methyl mercury by the microalga Chlamydomonas reinhardtii. Environ. Pollut. 2014, 192, 212–215. [Google Scholar] [CrossRef]
- Laurier, F.; Mason, R.; Gill, G.; Whalin, L. Mercury distributions in the North Pacific Ocean—20 years of observations. Mar. Chem. 2004, 90, 3–19. [Google Scholar] [CrossRef]
- Fu, X.; Feng, X.; Zhang, G.; Xu, W.; Li, X.; Yao, H.; Liang, P.; Li, J.; Sommar, J.; Yin, R.; et al. Mercury in the marine boundary layer and seawater of the South China Sea: Concentrations, sea/air flux, and implication for land outflow. J. Geophys. Res. Atmos. 2010, 115. [Google Scholar] [CrossRef] [Green Version]
- Sinclair, K.A.; Xie, Q.; Mitchell, C.P. Methylmercury in water, sediment, and invertebrates in created wetlands of Rouge Park, Toronto, Canada. Environ. Pollut. 2012, 171, 207–215. [Google Scholar] [CrossRef]
- Ci, Z.; Zhang, X.; Wang, Z.; Niu, Z. Phase speciation of mercury (Hg) in coastal water of the Yellow Sea, China. Mar. Chem. 2011, 126, 250–255. [Google Scholar] [CrossRef]
- Skrobonja, A.; Gojkovic, Z.; Soerensen, A.L.; Westlund, P.-O.; Funk, C.; Björn, E. Uptake Kinetics of Methylmercury in a Freshwater Alga Exposed to Methylmercury Complexes with Environmentally Relevant Thiols. Environ. Sci. Technol. 2019, 53, 13757–13766. [Google Scholar] [CrossRef]
- Wu, P.; Kainz, M.J.; Bravo, A.G.; Åkerblom, S.; Sonesten, L.; Bishop, K. The importance of bioconcentration into the pelagic food web base for methylmercury biomagnification: A meta-analysis. Sci. Total Environ. 2019, 646, 357–367. [Google Scholar] [CrossRef]
- Folgar, S.; Torres, E.; Pérez-Rama, M.; Cid, A.; Herrero, C.; Abalde, J. Dunaliella salina as marine microalga highly tolerant to but a poor remover of cadmium. J. Hazard. Mater. 2009, 165, 486–493. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Xu, X.; Qu, B.; Liu, X.; Yi, W.; Zhang, H. Study on Adsorption Properties of Modified Corn Cob Activated Carbon for Mercury Ion. Energies 2021, 14, 4483. [Google Scholar] [CrossRef]
- Zhu, Q.-L.; Guo, S.-N.; Wen, F.; Zhang, X.-L.; Wang, C.-C.; Si, L.-F.; Zheng, J.-L.; Liu, J. Transcriptional and physiological responses of Dunaliella salina to cadmium reveals time-dependent turnover of ribosome, photosystem, and ROS-scavenging pathways. Aquat. Toxicol. 2019, 207, 153–162. [Google Scholar] [CrossRef]
- Beauvais-Flück, R.; Slaveykova, V.I.; Ulf, S.; Cosio, C. Towards early-warning gene signature of Chlamydomonas reinhardtii exposed to Hg-containing complex media. Aquat. Toxicol. 2019, 214, 105259. [Google Scholar] [CrossRef]
- Pickhardt, P.C.; Fisher, N.S. Accumulation of Inorganic and Methylmercury by Freshwater Phytoplankton in Two Contrasting Water Bodies. Environ. Sci. Technol. 2007, 41, 125–131. [Google Scholar] [CrossRef]
- Slaveykova, V.I.; Majumdar, S.; Regier, N.; Li, W.; Keller, A.A. Metabolomic Responses of Green Alga Chlamydomonas reinhardtii Exposed to Sublethal Concentrations of Inorganic and Methylmercury. Environ. Sci. Technol. 2021, 55, 3876–3887. [Google Scholar] [CrossRef]
- Wu, Y.; Wang, W.-X. Accumulation, subcellular distribution and toxicity of inorganic mercury and methylmercury in marine phytoplankton. Environ. Pollut. 2011, 159, 3097–3105. [Google Scholar] [CrossRef]
- Zhang, P.; Sun, J.; Chen, J.; Wei, J.; Zhao, W.; Liu, Q.; Sun, H. Effect of feeding selectivity on the transfer of methylmercury through experimental marine food chains. Mar. Environ. Res. 2013, 89, 39–44. [Google Scholar] [CrossRef]
- Tada, Y.; Marumoto, K. Uptake of methylmercury by marine microalgae and its bioaccumulation in them. J. Oceanogr. 2020, 76, 63–70. [Google Scholar] [CrossRef]
- Moye, H.A.; Miles, C.J.; Phlips, E.J.; Sargent, B.; Merritt, K.K. Kinetics and Uptake Mechanisms for Monomethylmercury between Freshwater Algae and Water. Environ. Sci. Technol. 2002, 36, 3550–3555. [Google Scholar] [CrossRef]
- Gorski, P.R.; Armstrong, D.E.; Hurley, J.P.; Shafer, M.M. Speciation of aqueous methylmercury influences uptake by a freshwater alga (Selenastrum capricornutum). Environ. Toxicol. Chem. 2006, 25, 534–540. [Google Scholar] [CrossRef]
- Bouchet, S.; Björn, E. Analytical developments for the determination of monomethylmercury complexes with low molecular mass thiols by reverse phase liquid chromatography hyphenated to inductively coupled plasma mass spectrometry. J. Chromatogr. A 2014, 1339, 50–58. [Google Scholar] [CrossRef]
- Garcia-Soto, C.; Cheng, L.; Caesar, L.; Schmidtko, S.; Jewett, E.B.; Cheripka, A.; Rigor, I.; Caballero, A.; Chiba, S.; Báez, J.C.; et al. An Overview of Ocean Climate Change Indicators: Sea Surface Temperature, Ocean Heat Content, Ocean pH, Dissolved Oxygen Concentration, Arctic Sea Ice Extent, Thickness and Volume, Sea Level and Strength of the AMOC (Atlantic Meridional Overturning Circulation). Front. Mar. Sci. 2021, 8, 642372. [Google Scholar] [CrossRef]
- Liang, X.; Zhu, N.; Johs, A.; Chen, H.; Pelletier, D.A.; Zhang, L.; Yin, X.; Gao, Y.; Zhao, J.; Gu, B. Mercury Reduction, Uptake, and Species Transformation by Freshwater Alga Chlorella vulgaris under Sunlit and Dark Conditions. Environ. Sci. Technol. 2022, 56, 4961–4969. [Google Scholar] [CrossRef]
- Liem-Nguyen, V.; Nguyen-Ngoc, H.-T.; Adediran, G.A.; Björn, E. Determination of picomolar levels of methylmercury complexes with low molecular mass thiols by liquid chromatography tandem mass spectrometry and preconcentration. Anal. Bioanal. Chem. 2020, 412, 1619–1628. [Google Scholar] [CrossRef] [Green Version]
- Minamata Convention on Mercury. UN Environment Programme. 2013. Available online: https://mercuryconvention.org/en (accessed on 25 June 2023).
- World Health Organisation. Mercury and Health. 2017. Available online: https://www.who.int/news-room/fact-sheets/detail/mercury-and-health (accessed on 25 June 2023).
- Directive 2008/105/EC of the European Parliament. EUR-Lex. 2008. Available online: https://eur-lex.europa.eu/eli/dir/2008/105/oj (accessed on 25 June 2023).
- Commission Regulation (EU) 2023/915 on Maximum Levels for Certain Contaminants in Food and Repealing Regulation (EC) No 1881/2006. EUR-Lex. 2023. Available online: https://eur-lex.europa.eu/eli/reg/2023/915/oj (accessed on 25 June 2023).
- Yin, X.; Wang, L.; Liang, X.; Zhang, L.; Zhao, J.; Gu, B. Contrary effects of phytoplankton Chlorella vulgaris and its exudates on mercury methylation by iron- and sulfate-reducing bacteria. J. Hazard. Mater. 2022, 433, 128835. [Google Scholar] [CrossRef]
- Cavicchioli, R.; Ripple, W.J.; Timmis, K.N.; Azam, F.; Bakken, L.R.; Baylis, M.; Behrenfeld, M.J.; Boetius, A.; Boyd, P.W.; Classen, A.T.; et al. Scientists’ warning to humanity: Microorganisms and climate change. Nat. Rev. Microbiol. 2019, 17, 569–586. [Google Scholar] [CrossRef] [Green Version]
- Ighalo, J.O.; Dulta, K.; Kurniawan, S.B.; Omoarukhe, F.O.; Ewuzie, U.; Eshiemogie, S.O.; Ojo, A.U.; Abdullah, S.R.S. Progress in Microalgae Application for CO2 Sequestration. Clean. Chem. Eng. 2022, 3, 100044. [Google Scholar] [CrossRef]
- Even, C.; Hadroug, D.; Boumlaik, Y.; Simon, G. Microalgae-based Bioenergy with Carbon Capture and Storage quantified as a Negative Emissions Technology. Energy Nexus 2022, 7, 100117. [Google Scholar] [CrossRef]
- Di Lena, G.; Casini, I.; Lucarini, M.; del Pulgar, J.S.; Aguzzi, A.; Caproni, R.; Gabrielli, P.; Lombardi-Boccia, G. Chemical characterization and nutritional evaluation of microalgal biomass from large-scale production: A comparative study of five species. Eur. Food Res. Technol. 2020, 246, 323–332. [Google Scholar] [CrossRef]
- Le Faucheur, S.; Campbell, P.G.; Fortin, C.; Slaveykova, V.I. Interactions between mercury and phytoplankton: Speciation, bioavailability, and internal handling. Environ. Toxicol. Chem. 2014, 33, 1211–1224. [Google Scholar] [CrossRef]
- Tang, W.; He, M.; Chen, B.; Ruan, G.; Xia, Y.; Xu, P.; Song, G.; Bi, Y.; Hu, B. Investigation of toxic effect of mercury on Microcystis aeruginosa: Correlation between intracellular mercury content at single cells level and algae physiological responses. Sci. Total Environ. 2023, 858, 159894. [Google Scholar] [CrossRef]
- Satoh, M.; Hirachi, Y.; Yoshioka, A.; Kobayashi, M.; Oyama, Y. Determination of cellular levels of nonprotein thiols in phytoplankton and their correlations with susceptibility to mercury. J. Phycol. 2002, 38, 983–990. [Google Scholar] [CrossRef]
- Quevedo-Ospina, C.; Arroyave, C.; Peñuela-Vásquez, M.; Villegas, A. Effect of mercury in the influx and efflux of nutrients in the microalga Desmodesmus armatus. Aquat. Toxicol. 2023, 258, 106496. [Google Scholar] [CrossRef]
- Dranguet, P.; Cosio, C.; Le Faucheur, S.; Beauvais-Flück, R.; Freiburghaus, A.; Worms, I.A.; Petit, B.; Civic, N.; Docquier, M.; Slaveykova, V.I. Transcriptomic approach for assessment of the impact on microalga and macrophyte of in-situ exposure in river sites contaminated by chlor-alkali plant effluents. Water Res. 2017, 121, 86–94. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Wang, W.-X. Thiol compounds induction kinetics in marine phytoplankton during and after mercury exposure. J. Hazard. Mater. 2012, 217–218, 271–278. [Google Scholar] [CrossRef]
- Chevrollier, L.-A.; Koski, M.; Søndergaard, J.; Trapp, S.; Aheto, D.W.; Darpaah, G.; Nielsen, T.G. Bioaccumulation of metals in the planktonic food web in the Gulf of Guinea. Mar. Pollut. Bull. 2022, 179, 113662. [Google Scholar] [CrossRef]
- Khatiwada, B.; Hasan, M.T.; Sun, A.; Kamath, K.S.; Mirzaei, M.; Sunna, A.; Nevalainen, H. Proteomic response of Euglena gracilis to heavy metal exposure—Identification of key proteins involved in heavy metal tolerance and accumulation. Algal Res. 2020, 45, 101764. [Google Scholar] [CrossRef]
- Gómez-Jacinto, V.; García-Barrera, T.; Gómez-Ariza, J.L.; Garbayo-Nores, I.; Vílchez-Lobato, C. Elucidation of the defence mechanism in microalgae Chlorella sorokiniana under mercury exposure. Identification of Hg–phytochelatins. Chem. Interact. 2015, 238, 82–90. [Google Scholar] [CrossRef]


{kind=link}
| Location | Total Hg | MeHg | References |
|---|---|---|---|
| - | pM or ng/L | pM, fM or ng/L | - |
| Adour Estuary (France) | 0.51–3.42 ng/L | 0.025–0.081 ng L | [43] |
| Adriatic Sea | 1.46 pM | 0.28 pM | [44] |
| Amazon River | 2.8 ng/L | - | [45] |
| Arctic Ocean (depth) | 0.5 pM | - | [46] |
| Arctic Ocean (surface) | 1.1 pM | - | [46] |
| Atlantic Ocean (Southern Polar Front) | 0.93 ± 0.69 ng/L | 0.26 ± 0.12 ng/L | [41] |
| Atlantic Ocean (north) | 2.4 pM | - | [47] |
| Attawapiskat Drainage Basin (Canada) | 0.32–7.4 ng/L | 0.004–0.09 ng/L | [48] |
| Average in oceans | 1.5 pM | - | [49] |
| Average in surface water of lakes and rivers | - | 0.003–1.03 ng/L | [34] |
| Baltic Sea (northen) | 1.0 ± 0.3 pM | 37 ± 15 fM; 21 ± 9 fM | [37,50] |
| Baltic Sea (southern) | 1.5 ± 0.7 pM | 23 ± 13 fM | [50] |
| Bothnian Bay (Baltic) | 1.24 ± 0.3 pM | 80 ± 25 fM | [51] |
| Bothnian Bay (Baltic) | 11.5 ± 1.66 pM | 116–236 fM | [52] |
| Bothnian Sea (Baltic) | 0.84 ± 0.24 pM | 21 ± 9 fM | [51] |
| Carson River (Nevada) | 29.1 ng/L | 1.21 ng/L | [53] |
| Cauca River basin (Columbia) | 650 ng/L | - | [42] |
| Crimean saline lakes | 129 ng/L | - | [54] |
| Florida Bay (discharging canals) | 3–7.4 ng/L | <0.03–52% of THg | [55] |
| Jiaozhou Bay (Yellow Sea) | 8.46–27.3 ng/L | 0.08–0.83 ng/L | [56] |
| Oil Sands Region Lakes (Canada) | 0.4–5.3 ng/L | 0.01–0.34 ng/L | [57] |
| Lake Titicaca (Bolivia) | - | 0.01–0.18 ng/L | [58] |
| Lake Victoria (Africa) | 3–15 ng/L | - | [59] |
| Mediterranean Sea | 1.0 pM; 2.5 pM | - | [60,61] |
| Mediterranean Sea | 1.46 ± 0.41 pM | 0.28 ± 0.05 pM | [44] |
| Mekong River | 1.3 ± 0.4 ng/L | 0.05 ± 0.03 ng/L | [62] |
| Olt River (Romania) | 8–88 ng/L | 0.7 ng/L | [63] |
| Pacific Ocean | 1.2 pM | - | [64] |
| Råne River estuary (Baltic) | 2.0–5.95 pM | 306 fM | [51] |
| South China Sea | 0.8–2.3 ng/L | 0,05–0.22 ng/L | [65] |
| Tapajos River (Brazil) | 1.8 ng/L | 1.46 ± 0.41 pM | [45] |
| In Wetlands of Rouge Park (Canada) | 1.45 ± 0.91 ng/L | 0.59 ± 0.45 ng/L | [66] |
| Yellow Sea | 6.7–27.5 pM | - | [67] |
| Microalgae or Cyanobacteria Strain | Experimental MeHg Conc. | Time of Exposure | MeHg Uptake | References |
|---|---|---|---|---|
| - | μg/L; ng/L; pM; nM | h | ng/gDW; µg/gDW; ag/µm3 Biomass; amol/Cell | - |
| Chlamydomonas reinhardtii | 97 ± 11 pM | 48 h | 1.4 ± 0.19 × 10−2 amol/cell | [63] |
| Chlamydomonas reinhardtii | 0.64−0.74 nM | 48 h | 17 ng/gDW | [74] |
| Chlamydomonas reinhardtii | 5 nM | 2 h | 2 ng/gDW | [75] |
| Chlamydomonas reinhardtii | 50 nM | 2 h | 18 ng/gDW | [75] |
| Chlorella autotrophica | 590 ng/L | 72 h | 132.7 µg/gDW | [76] |
| Chlorella sp. | 1 μg/L | 72 h | 27.91 µg/gDW | [77] |
| Cyanophyceae | 0.7 ng/L | 1 h | 0.588 ag/µm3 biomass | [78] |
| Isochrysis galbana | 590 ng/L | 72 h | 88.5 µg/gDW | [76] |
| Isochrysis galbana | 1 μg/L | 72 h | 40.03 µg/gDW | [77] |
| Natural consortium: (Oedogonium spp. Chlorella spp. Scenedesmus spp.) | 0.995 nM (200 ng/L) | 6 h | 340–400 ng/gDW | [34] |
| Nitzschia closterium | 1 μg/L | 72 h | 32.74 µg/gDW | [77] |
| Pelagophyceae | 0.7 ng/L | 1 h | 0.236 ag/µm3 biomass | [78] |
| Schizothrix calcicola | 1.9 nM | 0.083 h | 356 ± 22.1 ng/gDW | [79] |
| Selenastrum capricornutum | 1 pM (2 ng/L) | 48 h | 180.7 ng/gDW | [80] |
| Selenastrum capricornutum | 233 nM | 45 h | 0.294 ng/gDW | [81] |
| Synechococcus sp. | 0.7 ng/L | 1 h | 0.63 ag/µm3 biomass | [78] |
| Thalassiosira pseudonana | 3 nM (600 ng/L) | 72 h | 22.1 µg/gDW | [76] |
| Thalassiosira weissflogii | 1.9 nM | 0.083 h | 473 ± 30.5 ng/gDW | [79] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gojkovic, Z.; Simansky, S.; Sanabria, A.; Márová, I.; Garbayo, I.; Vílchez, C. Interaction of Naturally Occurring Phytoplankton with the Biogeochemical Cycling of Mercury in Aquatic Environments and Its Effects on Global Hg Pollution and Public Health. Microorganisms 2023, 11, 2034. https://doi.org/10.3390/microorganisms11082034
Gojkovic Z, Simansky S, Sanabria A, Márová I, Garbayo I, Vílchez C. Interaction of Naturally Occurring Phytoplankton with the Biogeochemical Cycling of Mercury in Aquatic Environments and Its Effects on Global Hg Pollution and Public Health. Microorganisms. 2023; 11(8):2034. https://doi.org/10.3390/microorganisms11082034
Chicago/Turabian StyleGojkovic, Zivan, Samuel Simansky, Alain Sanabria, Ivana Márová, Inés Garbayo, and Carlos Vílchez. 2023. "Interaction of Naturally Occurring Phytoplankton with the Biogeochemical Cycling of Mercury in Aquatic Environments and Its Effects on Global Hg Pollution and Public Health" Microorganisms 11, no. 8: 2034. https://doi.org/10.3390/microorganisms11082034
APA StyleGojkovic, Z., Simansky, S., Sanabria, A., Márová, I., Garbayo, I., & Vílchez, C. (2023). Interaction of Naturally Occurring Phytoplankton with the Biogeochemical Cycling of Mercury in Aquatic Environments and Its Effects on Global Hg Pollution and Public Health. Microorganisms, 11(8), 2034. https://doi.org/10.3390/microorganisms11082034