
Vaginal Microbial Network Analysis Reveals Novel Taxa Relationships among Adolescent and Young Women with Incident Sexually Transmitted Infection Compared with Those Remaining Persistently Negative over a 30-Month Period
 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Design, Participants, and Sample Size
2.2. Detection of Bacterial vaginosis and Sexually Transmitted Infections
2.3. Data Collection
2.4. Characterization of Vaginal Microbiome
2.5. Construction of Analytic Data Set
2.6. Statistical Analysis
3. Results
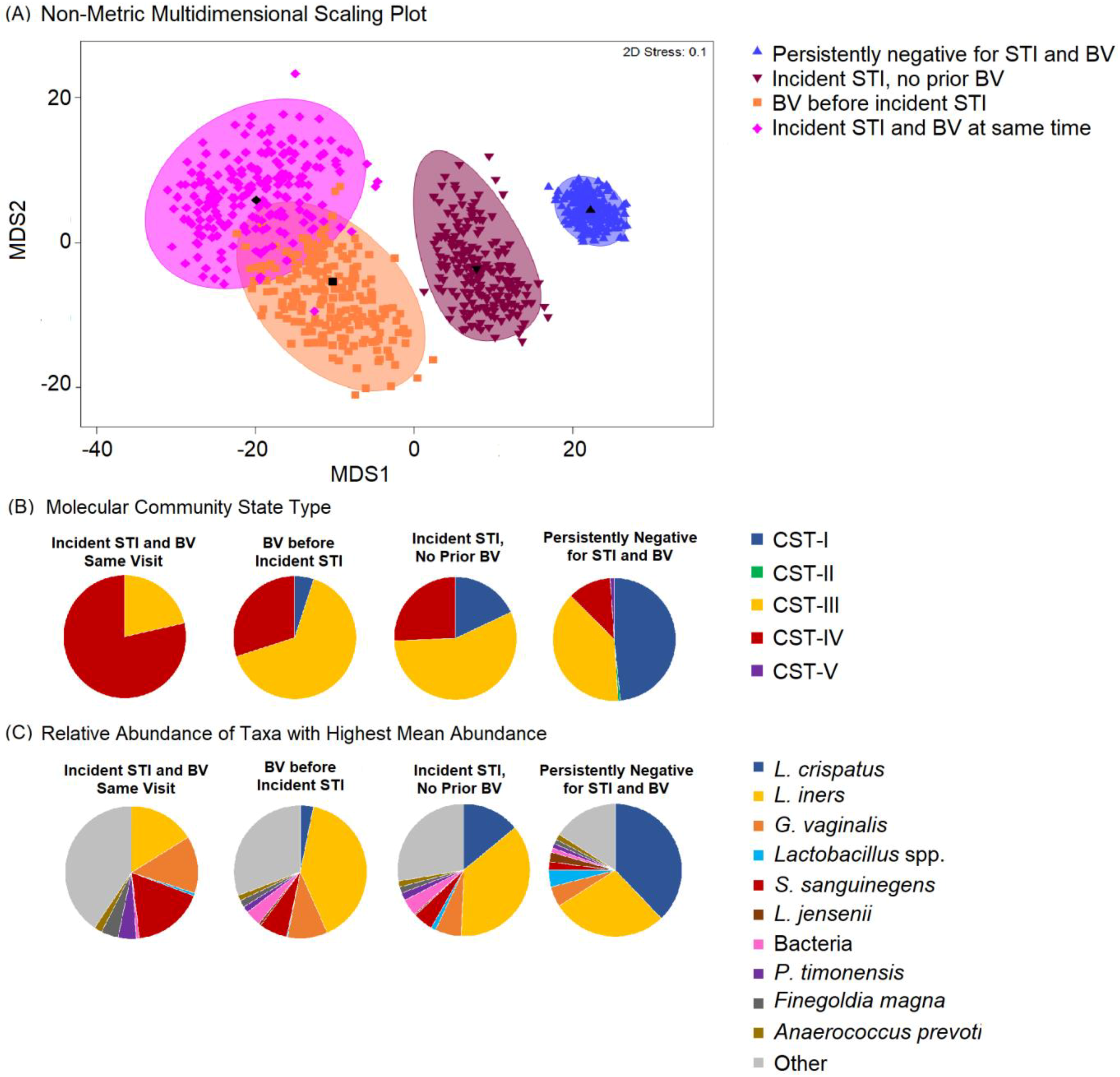
3.1. Characteristics of Study Sample and Microbiome Composition
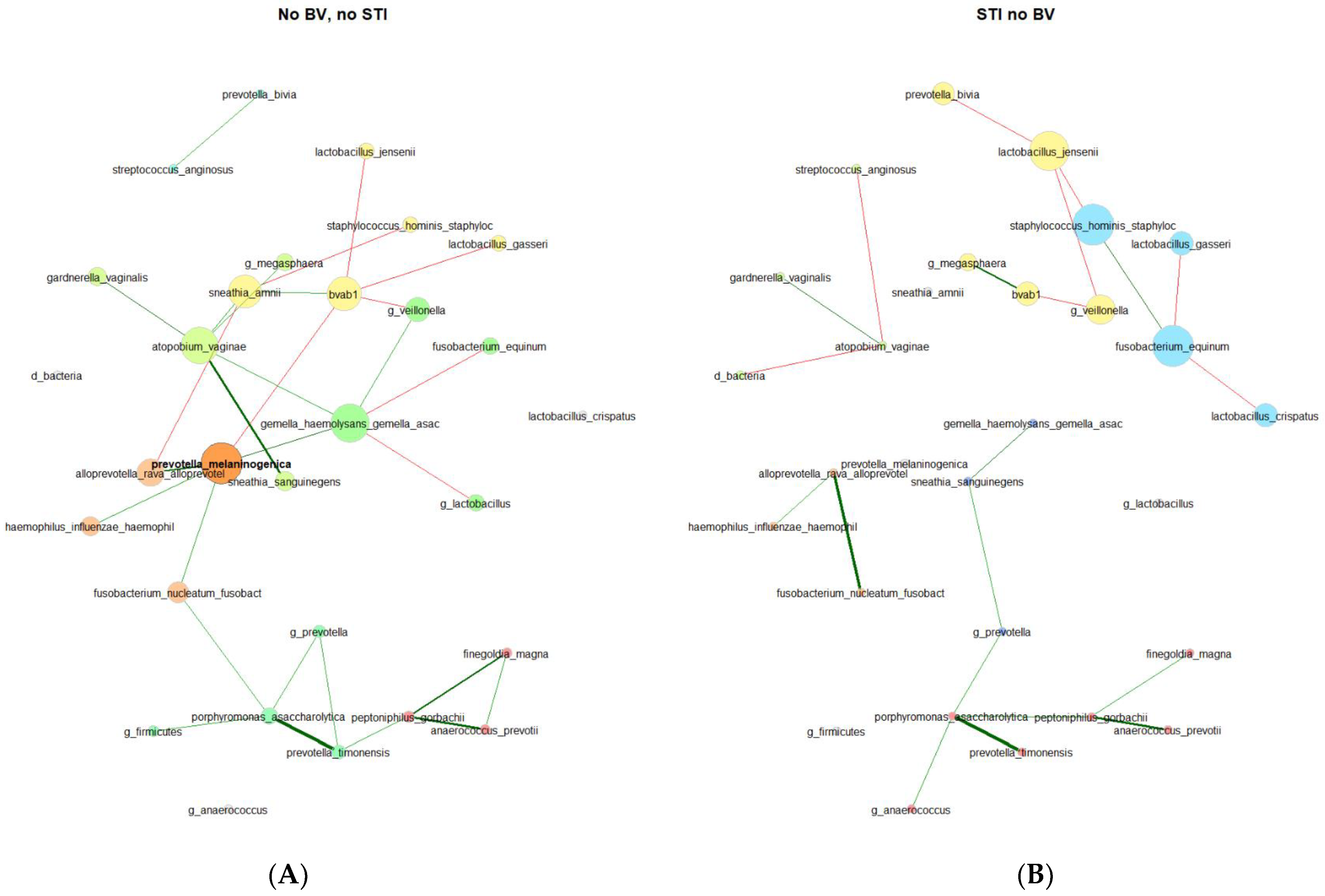
3.2. Results of Microbial Co-Occurrence Network Analysis: Differences in Network Properties and Centralities for Participants with Incident STI Compared to Persistently Negative Participants
3.3. Results of Participant-Level Network Analysis: Network Properties Differ by Sociodemographic, Behavioral, and VMB Composition
4. Discussion
Limitations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Peebles, K.; Velloza, J.; Balkus, J.E.; McClelland, R.S.; Barnabas, R.V. High global burden and costs of bacterial vaginosis: A systematic review and meta-analysis. Sex. Transm. Dis. 2019, 46, 304–311. [Google Scholar] [CrossRef]
- Atashili, J.; Poole, C.; Ndumbe, P.M.; Adimora, A.A.; Smith, J.S. Bacterial vaginosis and HIV acquisition: A meta-analysis of published studies. AIDS 2008, 22, 1493–1501. [Google Scholar] [CrossRef] [Green Version]
- Leitich, H.; Kiss, H. Asymptomatic bacterial vaginosis and intermediate flora as risk factors for adverse pregnancy outcome. Clin. Obstet. Gynaecol. 2007, 21, 375–390. [Google Scholar] [CrossRef] [PubMed]
- Mohanty, T.; Doke, P.P.; Khuroo, S.R. Effect of bacterial vaginosis on preterm birth: A meta-analysis. Arch. Gynecol. Obstet. 2022. Ahead of print. [Google Scholar] [CrossRef] [PubMed]
- Torrone, E.A.; Morrison, C.S.; Chen, P.-L.; Kwok, C.; Francis, S.C.; Hayes, R.J.; Looker, K.J.; McCormack, S.; McGrath, N.; van de Wijgert, J.H.; et al. Prevalence of sexually transmitted infections and bacterial vaginosis among women in sub-Saharan Africa: An individual participant data meta-analysis of 18 HIV prevention studies. PLoS Med. 2018, 15, e1002511. [Google Scholar] [CrossRef] [Green Version]
- Allsworth, J.E.; Peipert, J.F. Severity of bacterial vaginosis and the risk of sexually transmitted infection. Am. J. Obstet. Gynecol. 2011, 205, 113.e1–113.e6. [Google Scholar] [CrossRef] [Green Version]
- Brotman, R.M.; Klebanoff, M.A.; Nansel, T.R.; Yu, K.F.; Andrews, W.W.; Zhang, J.; Schwebke, J.R. Bacterial vaginosis assessed by Gram stain and diminished colonization resistance to incident gonococcal, chlamydia, and trichomonal genital infection. J. Infect. Dis. 2010, 202, 1907–1915. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Sexually Transmitted Infections among Adolescents. The Need for Adequate Health Services. Technical Document. 2005. Available online: https://www.who.int/publications/i/item/9241562889 (accessed on 13 February 2023).
- McKinnon, L.R.; Achilles, S.L.; Bradshaw, C.S.; Burgener, A.; Crucitti, T.; Fredricks, D.N.; Jaspan, H.B.; Kaul, R.; Kaushic, C.; Klatt, N.; et al. The evolving facets of Bacterial vaginosis: Implications for HIV transmission. AIDS Res. Hum. Retroviruses 2019, 35, 219–228. [Google Scholar] [CrossRef] [Green Version]
- Ravel, J.; Brotman, R.M. Translating the vaginal microbiome: Gaps and challenges. Genome Med. 2016, 8, 35. [Google Scholar] [CrossRef] [Green Version]
- Gosmann, C.; Anahtar, M.N.; Handley, S.A.; Farcasanu, M.; Abu-Ali, G.; Bowman, B.A.; Padavattan, N.; Desai, C.; Droit, L.; Moodley, A.; et al. Lactobacillus-deficient cervicovaginal bacterial communities are associated with increased HIV acquisition in young South African women. Immunity 2017, 46, 29–37. [Google Scholar] [CrossRef] [Green Version]
- Mehta, S.D.; Zulaika, G.; Otieno, F.O.; Nyothach, E.; Agingu, W.; Bhaumik, R.; Green, S.J.; van Eijk, A.M.; Kwaro, D.; Phillips-Howard, P.A.; et al. High prevalence of Lactobacillus crispatus dominated vaginal microbiome among Kenya secondary schoolgirls: Negative effects of poor quality menstrual hygiene management and sexual activity. Front. Cell Infect. Microbiol. 2021, 11, 617537. [Google Scholar] [CrossRef] [PubMed]
- Nugent, R.P.; Krohn, M.A.; Hillier, S.L. Reliability of diagnosing bacterial vaginosis is improved by a standardized method of gram stain interpretation. J. Clin. Microbiol. 1991, 29, 297–301. [Google Scholar] [CrossRef] [PubMed]
- National AIDS and STI Control Programme, Ministry of Health, Kenya. Algorithms for Managing Common STI Syndromes; NASCOP: Nairobi, Kenya, 2015. Available online: http://www.nascop.or.ke/index.php/recently-launched-guidelines/ (accessed on 6 June 2023).
- Centers for Disease Control and Prevention. 2015 Sexually Transmitted Diseases Treatment Guidelines. MMWR Recomm. Rep. 2021, 70. Available online: https://www.cdc.gov/std/treatment-guidelines/bv.htm (accessed on 16 August 2021).
- British Association of Sexual Health and HIV. Available online: https://www.bashhguidelines.org/current-guidelines/vaginal-discharge/bacterial-vaginosis-2012/ (accessed on 22 March 2021).
- Sherrard, J.; Wilson, J.; Donders, G.; Mendling, W.; Jensesn, J.S. 2018 European (IUSTI/WHO) International Union against Sexually Transmitted Infections (IUSTI) World Health Organisation (WHO) Guideline on the Management of Vaginal Discharge. Int. J. STD AIDS 2018, 29, 1258–1272. Available online: https://iusti.org/wp-content/uploads/2019/12/Vagdx2018.pdf (accessed on 6 June 2023). [CrossRef]
- Mehta, S.D.; Zulaika, G.; Agingu, W.; Nyothach, E.; Bhaumik, R.; Green, S.J.; van Eijk, A.M.; Kwaro, D.; Otieno, F.; Phillips-Howard, P. Analysis of Bacterial vaginosis, the vaginal microbiome, and sexually transmitted infections following the provision of menstrual cups in Kenyan schools: Results of a nested study within a cluster randomized controlled trial. PLoS Med. 2023, 20, e1004258. [Google Scholar] [CrossRef]
- Kabudula, C.W.; Houle, B.; Collinson, M.A.; Kahn, K.; Tollman, S.; Clark, S. Assessing changes in household socioeconomic status in rural South Africa, 2001–2013: A distributional analysis using household asset indicators. Soc. Indic. Res. 2017, 133, 1047–1073. [Google Scholar] [CrossRef] [Green Version]
- Alexander, K.T.; Zulaika, G.; Nyothach, E.; Oduor, C.; Mason, L.; Obor, D.; Eleveld, A.; Laserson, K.F.; Phillips-Howard, P.A. Do water, sanitation and hygiene conditions in primary schools consistently support schoolgirls’ menstrual needs? A longitudinal study in rural western Kenya. Int. J. Environ. Res. Public Health 2018, 15, 1682. [Google Scholar] [CrossRef] [Green Version]
- Naqib, A.; Poggi, S.; Wang, W.; Hyde, M.; Kunstman, K.; Green, S.J. Making and sequencing heavily multiplexed, high-throughput 16S ribosomal RNA gene amplicon libraries using a flexible, two-stage PCR protocol. In Gene Expression Analysis; Humana Press: New York, NY, USA, 2018; pp. 149–169. [Google Scholar]
- Zhang, J.; Kobert, K.; Flouri, T.; Stamatakis, A. PEAR: A fast and accurate Illumina Paired-End reAd mergeR. Bioinformatics 2014, 30, 614–620. [Google Scholar] [CrossRef] [Green Version]
- Holm, J.B.; Humphrys, M.S.; Robinson, C.K.; Settles, M.L.; Ott, S.; Fu, L.; Yang, H.; Gajer, P.; He, X.; McComb, E.; et al. Ultrahigh-throughput multiplexing and sequencing of >500-base-pair amplicon regions on the Illumina HiSeq 2500 Platform. mSystems 2019, 4, e00029-19. [Google Scholar] [CrossRef] [Green Version]
- France, M.; Ma, B.; Gajer, P.; Brown, S.; Humphrys, M.S.; Holm, J.B.; Waetjen, L.E.; Brotman, R.M.; Ravel, J. VALENCIA: A nearest centroid classification method for vaginal microbial communities based on composition. Microbiome 2020, 8, 166. [Google Scholar] [CrossRef]
- Davis, N.M.; Proctor, D.M.; Holmes, S.P.; Relman, D.A.; Callahan, B.J. Simple statistical identification and removal of contaminant sequences in marker-gene and metagenomics data. Microbiome 2018, 6, 226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berry, D.; Widder, S. Deciphering microbial interactions and detecting keystone species with co-occurrence networks. Front. Microbiol. 2014, 5, 219. [Google Scholar] [CrossRef] [Green Version]
- Peschel, S.; Müller, C.L.; von Mutius, E.; Boulesteix, A.-L.; Depner, M. NetCoMi: A network construction and comparison for microbiome data in R. Brief. Bioinform. 2021, 22, bbaa290. [Google Scholar] [CrossRef] [PubMed]
- Yoon, G.; Gaynanova, I.; Müller, C.L. Microbial Networks in SPRING—Semi-parametric rank-based correlation and partial correlation estimation for quantitative microbiome data. Front. Genet. 2019, 10, 516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nepusz, T.; Csardi, G. Community Structure Via Greed Optimization of Modularity. Available online: https://igraph.org/r/doc/cluster_fast_greedy.html (accessed on 7 June 2023).
- Clauset, A.; Newman, M.E.J.; Moore, C. Finding community structure in very large networks. Phys. Rev. E 2004, 70, 066111. [Google Scholar] [CrossRef] [Green Version]
- Ma, B.; Wang, Y.; Ye, S.; Liu, S.; Stirling, E.; Gilbert, J.A.; Faust, K.; Knight, R.; Jansson, J.K.; Cardona, C.; et al. Earth microbial co-occurrence network reveals interconnection pattern across microbiomes. Microbiome 2020, 8, 82. [Google Scholar] [CrossRef]
- Quaye, E.K.; Adjei, R.L.; Isawumi, A.; Allen, D.J.; Caporaso, J.G.; Quaye, O. Altered faecal microbiota composition and structure of Ghanaian children with acute gastroenteritis. Int. J. Mol. Sci. 2023, 24, 3607. [Google Scholar] [CrossRef]
- Jiao, J.; Bie, M.; Xu, X.; Duan, D.; Li, Y.; Wu, Y.; Zhao, L. Entamoeba gingivalis is associated with periodontal conditions in Chinese young patients: A cross-sectional study. Front. Cell Infect. Microbiol. 2022, 12, 1020730. [Google Scholar] [CrossRef]
- Guo, B.; Zhang, L.; Sun, H.; Gao, M.; Yu, N.; Zhang, Q.; Mou, A.; Liu, Y. Microbial co-occurrence network topological properties link with reactor parameters and reveal importance of low-abundance genera. NPJ Biofilms Microbiomes 2022, 8, 3. [Google Scholar] [CrossRef]
- Salliss, M.E.; Maarsingh, J.D.; Garza, C.; Laniewski, P.; Herbst-Kralovetz, M.M. Veillonellaceae family members uniquely alter the cervical metabolic microenvironment in a human three-dimensional epithelial model. NPJ Biofilms Microbiomes 2021, 7, 57. [Google Scholar] [CrossRef]
- Wang, B.; Xiao, B.B.; Shang, C.G.; Wang, K.; Na, R.S.; Nu, X.X.; Liao, Q. Molecular analysis of the relationship between specific vaginal bacteria and bacterial vaginosis metronidazole therapy failure. Eur. J. Clin. Microbiol. Infect. Dis. 2014, 33, 1749–1756. [Google Scholar] [CrossRef] [PubMed]
- Van de Wijgert, J.H.H.M.; Borgdorff, H.; Verhelst, R.; Crucitti, T.; Francis, S.; Verstraelen, H.; Jespers, V. The vaginal microbiota: What have we learned after a decade of molecular characterization? PLoS ONE 2014, 9, e105998. [Google Scholar]
- Srinivasan, S.; Hoffman, N.G.; Morgan, M.T.; Matsen, F.A.; Fiedler, T.L.; Hall, R.W.; Ross, F.J.; McCoy, C.O.; Bumgarner, R.; Marrazzo, J.M.; et al. Bacterial ccommunities in women with bacterial vaginosis: High resolution phylogenetic analyses reveal relationships of microbiota to clinical criteria. PLoS ONE 2012, 7, e37818. [Google Scholar] [CrossRef] [Green Version]
- Petrova, M.I.; Lievens, E.; Malik, S.; Imholz, N.; Lebeer, S. Lactobacillus species as biomarkers and agents that can promote various aspects of vaginal health. Front. Physiol. 2015, 6, 81. [Google Scholar] [CrossRef] [Green Version]
- Baquero, F.; Coque, T.M.; Galan, J.C.; Martinez, J.L. The origin of niches and species in the bacterial world. Front. Microbiol. 2021, 12, 657986. [Google Scholar] [CrossRef] [PubMed]
- Tamarelle, J.; Ma, B.; Gajer, P.; Humprhys, M.S.; Terplan, M.; Mark, K.S.; Thiébaut, A.C.; Forney, L.J.; Brotman, R.M.; Delarocque-Astagneau, E.; et al. Non-optimal vaginal microbiota after azithromycin treatment for Chlamydia trachomatis infection. J. Infect. Dis. 2020, 221, 627–635. [Google Scholar] [CrossRef] [PubMed]
- Kenyon, C.R.; Delva, W.; Brotman, R.M. Differential sexual network connectivity offers a parsimonious explanation for population-level variations in the prevalence of bacterial vaginoisis: A data-driven, model-supported hypothesis. BMC Womens Health 2019, 19, 8. [Google Scholar] [CrossRef] [Green Version]
- Ranallo, R.T.; McDonald, L.C.; Halpin, A.L.; Hiltke, T.; Young, V.B. The state of microbiome science at the intersection of infectious diseases and antimicrobial resistance. J. Infect. Dis. 2021, 223 (Suppl. S3), S187–S193. [Google Scholar] [CrossRef]
- Mott, P.D.; Taylor, C.M.; Lillis, R.A.; Ardizzone, C.M.; Albritton, H.L.; Luo, M.; Calabresi, K.G.; Martin, D.H.; Myers, L.; Quayle, A.J. Differences in the genital microbiota in women who naturally clear Chlamydia trachomatis infection compared to women who do not clear: A pilot study. Front. Cell Infect. Microbiol. 2012, 11, 615770. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics 1 | Persistently BV and STI Negative, N = 179 n (%) | Incident STI with No Prior BV, N = 39 n (%) | Incident STI with Prior BV, N = 20 n (%) | Incident STI and BV at Same Time, N = 14 n (%) |
|---|---|---|---|---|
| At Baseline | ||||
| Randomization status | ||||
| Control arm | 92 (51.4) | 16 (41.0) | 13 (65.0) | 11 (78.6) |
| Cup arm | 87 (48.6) | 23 (59.0) | 7 (35.0) | 3 (21.4) |
| Median age in years | ||||
| <16.9 years | 101 (56.4) | 21 (53.9) | 11 (55.0) | 4 (28.6) |
| ≥16.9 years | 78 (43.6) | 18 (46.1) | 9 (45.0) | 10 (71.4) |
| Socioeconomic status score | ||||
| Highest quintiles | 129 (72.1) | 26 (66.7) | 12 (60.0) | 11 (78.6) |
| Lowest quintile | 50 (27.9) | 13 (33.3) | 8 (40.0) | 3 (21.4) |
| Water, sanitation, and hygiene score | ||||
| Higher | 70 (39.1) | 16 (41.0) | 9 (45.0) | 6 (42.9) |
| Lower | 109 (60.9) | 23 (59.0) | 11 (55.0) | 8 (57.1) |
| Sexually active | ||||
| No | 132 (74.2) | 24 (61.5) | 13 (65.0) | 10 (71.4) |
| Yes | 46 (25.8) | 15 (38.5) | 7 (35.0) | 4 (28.6) |
| Ever engaged in sex in exchange for money, favors, or things | ||||
| INo | 162 (91.0) | 33 (84.6) | 17 (85.0) | 13 (92.9) |
| Yes | 16 (9.0) | 6 (15.4) | 3 (15.0) | 1 (7.1) |
| Ever been forced, tricked, or coerced to have sex | ||||
| No | 146 (82.0) | 26 (66.7) | 18 (90.0) | 10 (71.4) |
| Yes | 32 (18.0) | 13 (33.3) | 2 (10.0) | 4 (28.6) |
| Currently has a boyfriend | ||||
| No | 173 (96.7) | 36 (92.3) | 17 (85.0) | 13 (92.9) |
| Yes | 6 (3.3) | 3 (7.7) | 3 (15.0) | 1 (7.1) |
| Community State Type (CST) | ||||
| CST-I | 101 (57.7) | 14 (35.9) | 4 (20.0) | 6 (42.9) |
| CST-II | 4 (2.3) | 1 (2.6) | 1 (5.0) | 0 (0.0) |
| CST-III | 53 (30.3) | 19 (48.7) | 7 (35.0) | 6 (42.9) |
| CST-IV | 12 (6.9) | 3 (7.7) | 8 (40.0) | 1 (7.1) |
| CST-V | 5 (2.9) | 2 (5.1) | 0 (0.0) | 1 (7.1) |
| At time of Incident STI (or time of sampling if persistently negative) | Persistently BV and STI Negative, N = 179 n (%) | Incident STI with no prior BV, N = 39 n (%) | Incident STI with Prior BV, N = 20 n (%) | Incident STI and BV at same time, N = 14 n (%) |
| Time of incident STI or time of sampling if persistently negative | ||||
| 12 months | 74 (41.3) | 22 (56.4) | 6 (30.0) | 3 (21.4) |
| 30 months | 105 (58.7) | 17 (43.6) | 14 (70.0) | 11 (78.6) |
| Median age in years | ||||
| <18.8 years | 95 (53.1) | 24 (61.5) | 8 (40.0) | 4 (28.6) |
| ≥18.8 years | 84 (46.9) | 15 (38.5) | 12 (60.0) | 10 (71.4) |
| Socioeconomic status score | ||||
| Median or higher | 102 (57.6) | 26 (68.4) | 9 (47.4) | 6 (46.2) |
| Below median | 75 (42.4) | 12 (31.6) | 10 (52.6) | 7 (53.8) |
| Ever sexually active | ||||
| No | 75 (42.6) | 12 (32.4) | 4 (21.1) | 2 (14.3) |
| Yes | 101 (57.4) | 25 (67.6) | 15 (78.9) | 12 (85.7) |
| Ever engaged in sex in exchange for money, favors, or things | ||||
| No | 161 (91.0) | 31 (81.6) | 17 (89.5) | 11 (78.6) |
| Yes | 16 (9.0) | 7 (18.4) | 2 (10.5) | 3 (21.4) |
| Ever been forced, tricked, or coerced to have sex | ||||
| No | 151 (85.8) | 28 (73.7) | 18 (94.7) | 10 (71.4) |
| Yes | 25 (14.2) | 10 (26.3) | 1 (5.3) | 4 (28.6) |
| Currently has a boyfriend | ||||
| No | 142 (80.2) | 24 (63.2) | 10 (52.6) | 6 (42.9) |
| Yes | 35 (18.9) | 14 (36.8) | 9 (47.4) | 8 (57.1) |
| Ever been pregnant 2 | ||||
| No | 103 (97.2) | 23 (85.2) | 10 (71.4) | 10 (83.3) |
| Yes | 3 (2.8) | 4 (14.8) | 4 (28.6) | 2 (16.7) |
| Community State Type | ||||
| CST-I | 89 (49.7) | 7 (18.0) | 1 (5.0) | 0 (0) |
| CST-II | 1 (0.6) | 0 (0) | 0 (0) | 0 (0) |
| CST-III | 66 (36.9) | 22 (56.4) | 13 (65.0) | 3 (21.4) |
| CST-IV | 21 (11.7) | 10 (25.6) | 6 (30.0) | 11 (78.6) |
| CST-V | 2 (1.1) | 0 (0) | 0 (0) | 0 (0) |
| Network Properties | Persistently Negative, No STI and No BV N = 179 | Incident STI, No Prior BV N = 39 |
|---|---|---|
| Components and sizes | 3 components 24 (1) 2 (1) 1 (5) | 4 components 9 (2) 4 (1) 3 (1) 1 (6) |
| Largest connected component (LCC) | ||
| Relative LCC size | 0.774 | 0.290 |
| Clustering coefficient | 0.277 | 0 |
| Modularity | 0.543 | 0.398 |
| Positive edge percentage | 71.4 | 25.0 |
| Edge density | 0.101 | 0.222 |
| Natural connectivity | 0.054 | 0.159 |
| Vertex connectivity | 1 | 1 |
| Edge connectivity | 1 | 1 |
| Average dissimilarity | 0.680 | 0.697 |
| Average path length | 3.66 | 2.84 |
| Whole network | ||
| Number of components | 7 | 10 |
| Clustering coefficient | 0.277 | 0 |
| Modularity | 0.563 | 0.719 |
| Positive edge percentage | 72.4 | 61.9 |
| Edge density | 0.062 | 0.045 |
| Natural connectivity | 0.04 | 0.039 |
| Network Centrality Measures 1 | |||
| Taxon | Persistently Negative, No STI and No BV N = 179 | Incident STI, no Prior BV N = 39 | Absolute Difference |
| Degree Centrality | |||
| Fannyhessea vaginae (Atopobium) | 0.167 | 0 | 0.167 |
| Prevotella melaninogenica | 0.167 | 0 | 0.167 |
| Gemella haemolysans/Gemella asaccharolytica | 0.167 | 0 | 0.167 |
| Bacterial vaginosus associated bacterium 1 (BVAB1) | 0.167 | 0.067 | 0.1 |
| Sneathia amnii | 0.133 | 0 | 0.133 |
| Lactobacillus jensenii | 0.033 | 0.1 | 0.067 |
| Fusobacterium equinum | 0.033 | 0.1 | 0.067 |
| Veillonella | 0.067 | 0.067 | 0 |
| Staphylococus hominus | 0.033 | 0.067 | 0.034 |
| Betweenness Centrality | |||
| Prevotella melaninogenica | 0.593 | 0 | 0.593 |
| Fusobacterium nucleatum | 0.443 | 0 | 0.443 |
| Porphyromonas asaccharolytica | 0.423 | 0 | 0.423 |
| Gemella haemolysans/Gemella asaccharolytica | 0.387 | 0 | 0.387 |
| Fannyhessea vaginae (Atopobium) | 0.273 | 0 | 0.273 |
| Lactobacillus jensenii | 0 | 0.679 | 0.679 |
| Staphylococcus hominis | 0 | 0.536 | 0.536 |
| Fusobacterium equinum | 0 | 0.464 | 0.464 |
| Veillonella | 0 | 0.429 | 0.429 |
| BVAB1 | 0.178 | 0.25 | 0.072 |
| Closeness Centrality | |||
| Prevotella melaninogenica | 0.525 | 0 | 0.525 |
| Gemella haemolysans/Gemella asaccharolytica | 0.505 | 0 | 0.505 |
| Fannyhessea vaginae (Atopobium) | 0.49 | 0 | 0.49 |
| BVAB1 | 0.48 | 0.483 | 0.003 |
| Sneathia amnii | 0.458 | 0 | 0.458 |
| Lactobacillus jensenii | 0.32 | 0.614 | 0.294 |
| Fusobacterium equinum | 0.33 | 0.583 | 0.352 |
| Staphylococcus hominis | 0.31 | 0.577 | 0.24 |
| Veillonella | 0.387 | 0.533 | 0.146 |
| Eigenvector Centrality | |||
| Prevotella melaninogenica | 1 | 0 | 1 |
| Gemella haemolysans/Gemella asaccharolytica | 0.912 | 0 | 0.912 |
| Fannyhessea vaginae (Atopobium) | 0.865 | 0 | 0.865 |
| BVAB1 | 0.787 | 0.465 | 0.779 |
| Sneathia amnii | 0.755 | 0 | 0.755 |
| Staphylococcus hominis | 0.221 | 1 | 0.779 |
| Fusobacterium equinum | 0.263 | 0.999 | 0.736 |
| Lactobacillus jensenii | 0.23 | 0.943 | 0.68 |
| Veillonella | 0.483 | 0.643 | 0.16 |
| Persistently STI and BV Negative, N = 179 1 | Incident STI with No BV, N = 39 2 | |||
|---|---|---|---|---|
| Betweenness Centrality Mean (SD) | Eigenvector Centrality Mean (SD) | Betweenness Centrality Mean (SD) | Eigenvector Centrality Mean (SD) | |
| Mean (standard deviation) | 0.022 (0.038) | 0.079 (0.155) | 0.055 (0.100) | |
| Characteristics at Baseline | ||||
| Intervention arm | * | |||
| Control | 0.023 (0.043) | 0.082 (0.173) | 0.086 (0.145) | 0.338 (0.303) |
| Menstrual cup | 0.022 (0.032) | 0.076 (0.135) | 0.033 (0.042) | 0.200 (0.184) |
| Water, sanitation, and hygiene score | * | |||
| Higher score | 0.024 (0.039) | 0.083 (0.151) | 0.047 (0.096) | 0.200 (0.254) |
| Lower score | 0.022 (0.038) | 0.076 (0.159) | 0.060 (0.105) | 0.296 (0.237) |
| Median age | ||||
| <16.9 years | 0.022 (0.039) | 0.080 (0.159) | 0.041 (0.102) | 0.226 (0.262) |
| 16.9 years or older | 0.024 (0.037) | 0.077 (0.152) | 0.070 (0.099) | 0.292 (0.228) |
| Socioeconomic status score | * | |||
| Higher quartiles | 0.025 (0.039) | 0.082 (0.161) | 0.072 (0.116) | 0.301 (0.277) |
| Lowest quartile | 0.018 (0.034) | 0.072 (0.141) | 0.021 (0.046) | 0.158 (0.127) |
| Sexually active | ||||
| No | 0.020 (0.038) | 0.075 (0.152) | 0.060 (0.104) | 0.291 (0.262) |
| Yes | 0.030 (0.038) | 0.092 (0.168) | 0.046 (0.097) | 0.201 (0.214) |
| Experienced coerced sex | ** | |||
| No | 0.020 (0.037) | 0.077 (0.150) | 0.055 (0.101) | 0.280 (0.257) |
| Yes | 0.035 (0.039) | 0.089 (0.183) | 0.054 (0.103) | 0.209 (0.224) |
| Had transactional sex | ||||
| No | 0.021 (0.036) | 0.075 (0.146) | 0.060 (0.108) | 0.286 (0.255) |
| Yes | 0.039 (0.051) | 0.124 (0.246) | 0.023 (0.025) | 0.091 (0.065) |
| Has a boyfriend | ** | |||
| No | 0.021 (0.037) | 0.074 (0.144) | 0.056 ().104) | 0.249 (0.241) |
| Yes | 0.068 (0.048) | 0.208 (0.356) | 0.037 (0.034) | 0.341 (0.352) |
| Vaginal Community State Type (CST) | *** | |||
| CST-I (L. crispatus dominated) | 0.024 (0.040) | 0.091 (0.136) | 0.067 (0.112) | 0.365 (0.273) |
| CST-III (L. iners dominated) | 0.024 (0.035) | 0.053 (0.159) | 0.053 (0.104) | 0.230 (0.229) |
| CST-IV (mixed) | 0.014 (0.039) | 0.015 (0.017) | 0.061 (0.092) | 0.088 (0.114) |
| Characteristics at Follow-Up | ||||
| Median age | ** | |||
| Below 18.8 years | 0.022 (0.039) | 0.106 (0.180) | 0.046 (0.095) | 0.252 (0.248) |
| 18.8 years or older | 0.023 (0.037) | 0.049 (0.115) | 0.068 (0.110) | 0.263 (0.251) |
| SES score | * | |||
| Above median | 0.027 (0.044) | 0.105 (0.191) | 0.035 (0.077) | 0.241 (0.219) |
| Below median | 0.016 (0.025) | 0.040 (0.069) | 0.087 (0.135) | 0.293 (0.313) |
| Sexually active | ||||
| No | 0.019 (0.034) | 0.079 (0.159) | 0.019 (0.029) | 0.195 (0.144) |
| Yes | 0.025 (0.039) | 0.078 (0.154) | 0.068 (0.119) | 0.230 (0.285) |
| Experienced coerced sex | ||||
| No | 0.024 (0.039) | 0.081 (0.160) | 0.047 (0.086) | 0.253 (0.235) |
| Yes | 0.013 (0.024) | 0.064 (0.124) | 0.064 (0.136) | 0.269 (0.298) |
| Had transactional sex | ||||
| No | 0.022 (0.037) | 0.074 (0.144) | 0.054 (0.106) | 0.258 (0.250) |
| Yes | 0.030 (0.042) | 0.115 (0.243) | 0.041 (0.076) | 0.255 (0.264) |
| Has a boyfriend | ||||
| No | 0.022 (0.038) | 0.081 (0.154) | 0.045 (0.096) | 0.267 (0.243) |
| Yes | 0.025 (0.034) | 0.064 (0.162) | 0.063 (0.109) | 0.240 (0.268) |
| CST at follow-up | * | *** | ** | |
| CST-I (L. crispatus dominated) | 0.024 (0.043) | 0.145 (0.120) | 0.040 (0.078) | 0.189 (0.143) |
| CST-III (L. iners dominated) | 0.025 (0.035) | 0.017 (0.040) | 0.078 (0.122) | 0.358 (0.271) |
| CST-IV (mixed) | 0.010 (0.021) | 0.002 (0.005) | 0.014 (0.016) | 0.081 (0.085) |
| Network clusters | *** | *** | ||
| 1 | 0.021 (0.032) | 0.006 (0.014) | 0.024 (0.021) | 0.073 (0.081) |
| 2 | 0.029 (0.053) | 0.272 (0.241) | 0.070 (0.124) | 0.367 (0.264) |
| 3 | 0.021 (0.033) | 0.047 (0.049) | 0.040 (0.063) | 0.122 (0.058) |
| STI etiology (comparison restricted to sole infections) | NA | NA | * | |
| C. trachomatis (n = 21) | 0.080 (0.125) | 0.359 (0.279) | ||
| N. gonorrhoeae (n = 3) | 0.021 (0.021) | 0.111 (0.017) | ||
| T. vaginalis (n = 11) | 0.031 (0.063) | 0.167 (0.146) | ||
| Persistently STI and BV Negative, N = 179 | Incident STI with No BV, N = 39 | |||||
|---|---|---|---|---|---|---|
| Cluster 1, N = 73 n (%) | Cluster 2, N = 39 n (%) | Cluster 3, N = 67 n (%) | Cluster1, N = 8 n (%) | Cluster 2, N = 23 n (%) | Cluster 3, N = 8 n (%) | |
| Baseline Characteristics | ||||||
| Intervention arm | ||||||
| Control | 39 (53.4) | 20 (51.3) | 33 (49.3) | 2 (25.0) | 11 (47.8) | 3 (37.5) |
| Menstrual cup | 34 (46.6) | 19 (48.7) | 34 (50.7) | 6 (75.0) | 12 (52.2) | 5 (62.5) |
| WASH score | * | |||||
| Lower | 27 (37.0) | 15 (38.5) | 28 (41.8) | 6 (75.0) | 5 (21.7) | 5 (62.5) |
| Higher | 46 (63.0) | 24 (61.5) | 39 (58.2) | 2 (25.0) | 18 (78.3) | 3 (37.5) |
| Median age | ||||||
| <16.9 years | 37 (50.7) | 23 (59.0) | 41 (61.2) | 5 (62.5) | 12 (52.2) | 4 (50.0) |
| 16.9 years and older | 36 (49.3) | 16 (41.0) | 26 (38.8) | 3 (37.5) | 11 (47.8) | 4 (50.0) |
| Socioeconomic score | ||||||
| Lower | 55 (75.3) | 26 (66.7) | 48 (71.6) | 6 (75.0) | 17 (73.9) | 3 (37.5) |
| Higher | 18 (24.7) | 13 (33.3) | 19 (28.4) | 2 (25.0) | 6 (26.1) | 5 (62.5) |
| Sexually active | * | |||||
| No | 50 (68.5) | 28 (71.8) | 54 (81.8) | 2 (25.0) | 18 (78.3) | 4 (50.0) |
| Yes | 23 (31.5) | 11 (28.2) | 12 (18.2) | 6 (75.0) | 5 (21.7) | 4 (50.0) |
| Experienced coerced sex | * | |||||
| No | 55 (75.3) | 33 (84.6) | 58 (87.9) | 4 (50.0) | 19 (82.6) | 5 (62.5) |
| Yes | 18 (24.7) | 6 (15.4) | 8 (12.1) | 4 (50.0) | 4 (17.4) | 3 (37.5) |
| Had transactional sex | * | |||||
| No | 64 (87.7) | 34 (87.2) | 64 (97.0) | 5 (62.5) | 22 (95.7) | 6 (75.0) |
| Yes | 9 (12.3) | 5 (12.8) | 2 (3.0) | 3 (37.5) | 1 (4.3) | 2 (25.0) |
| Has a boyfriend | ||||||
| No | 70 (95.9) | 37 (94.9) | 66 (98.5) | 8 (100) | 21 (91.3) | 7 (87.5) |
| Yes | 3 (4.1) | 2 (5.1) | 1 (1.5) | 0 (0) | 2 (8.7) | 1 (12.5) |
| Vaginal Community State Type (CST) | *** | |||||
| CST-I (L. crispatus dominated) | 29 (42.6) | 27 (79.4) | 45 (70.3) | 2 (28.6) | 11 (50.0) | 1 (14.3) |
| CST-III (L. iners dominated) | 32 (47.1) | 6 (17.6) | 15 (23.4) | 4 (57.1) | 11 (50.0) | 4 (57.1) |
| CST-IV (mixed) | 4 (10.3) | 1 (2.9) | 4 (6.3) | 1 (14.3) | 0 (0) | 2 (28.6) |
| Characteristics at Follow-Up | ||||||
| Median age | *** | |||||
| Below 18.8 years | 33 (45.2) | 31 (79.5) | 31 (46.3) | 4 (50.0) | 15 (65.2) | 5 (62.5) |
| 18.8 years or older | 40 (54.8) | 8 (20.5) | 36 (53.7) | 4 (50.0) | 8 (34.8) | 3 (37.5) |
| Socioeconomic score | ||||||
| Above median | 35 (48.6) | 23 (60.5) | 44 (65.7) | 5 (62.5) | 16 (69.6) | 5 (71.4) |
| Below median | 37 (51.4) | 15 (39.5) | 23 (34.3) | 3 (37.5) | 7 (30.4) | 2 (28.6) |
| Sexually active | ||||||
| No | 29 (40.3) | 18 (47.4) | 28 (42.4) | 2 (28.6) | 7 (30.4) | 3 (42.9) |
| Yes | 43 (59.7) | 20 (52.6) | 38 (57.8) | 5 (71.4) | 16 (69.6) | 4 (57.1) |
| Experienced coerced sex | ||||||
| No | 58 (81.7) | 34 (89.5) | 59 (88.1) | 7 (87.5) | 16 (69.6) | 5 (71.4) |
| Yes | 13 (18.3) | 4 (10.5) | 8 (11.9) | 1 (12.5) | 7 (30.4) | 2 (28.6) |
| Had transactional sex | ||||||
| No | 63 (87.5) | 35 (92.1) | 63 (94.0) | 7 (87.5) | 18 (78.3) | 6 (85.7) |
| Yes | 9 (12.5) | 3 (7.9) | 4 (6.0) | 1 (12.5) | 5 (21.7) | 1 (14.3) |
| Has a boyfriend | ||||||
| No | 55 (76.4) | 35 (92.1) | 52 (77.6) | 4 (50.0) | 16 (69.6) | 4 (57.1) |
| Yes | 17 (23.6) | 3 (7.9) | 15 (22.4) | 4 (50.0) | 7 (30.4) | 3 (42.8) |
| Vaginal CST at follow-up | *** | *** | ||||
| CST-I (L. crispatus dominated) | 2 (2.8) | 36 (92.3) | 51 (78.5) | 0 (0) | 7 (30.4) | 0 (0) |
| CST-III (L. iners dominated) | 50 (69.4) | 3 (7.7) | 13 (20.0) | 1 (12.5) | 14 (60.9) | 7 (87.5) |
| CST-IV (mixed) | 20 (27.8) | 0 (0) | 1 (1.5) | 7 (87.5) | 2 (8.7) | 1 (12.5) |
| Etiology (restricted to single infections) | ||||||
| C. trachomatis (n = 28) | 2 (28.6) | 13 (61.9) | 6 (85.7) | |||
| N. gonorrhoeae (n = 4) | 0 (0) | 2 (9.5) | 1 (14.3) | |||
| T. vaginalis (n = 12) | 5 (71.4) | 6 (28.6) | 0 (0) | |||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mehta, S.D.; Agingu, W.; Zulaika, G.; Nyothach, E.; Bhaumik, R.; Green, S.J.; van Eijk, A.M.; Otieno, F.O.; Phillips-Howard, P.A.; Schneider, J. Vaginal Microbial Network Analysis Reveals Novel Taxa Relationships among Adolescent and Young Women with Incident Sexually Transmitted Infection Compared with Those Remaining Persistently Negative over a 30-Month Period. Microorganisms 2023, 11, 2035. https://doi.org/10.3390/microorganisms11082035
Mehta SD, Agingu W, Zulaika G, Nyothach E, Bhaumik R, Green SJ, van Eijk AM, Otieno FO, Phillips-Howard PA, Schneider J. Vaginal Microbial Network Analysis Reveals Novel Taxa Relationships among Adolescent and Young Women with Incident Sexually Transmitted Infection Compared with Those Remaining Persistently Negative over a 30-Month Period. Microorganisms. 2023; 11(8):2035. https://doi.org/10.3390/microorganisms11082035
Chicago/Turabian StyleMehta, Supriya D., Walter Agingu, Garazi Zulaika, Elizabeth Nyothach, Runa Bhaumik, Stefan J. Green, Anna Maria van Eijk, Fredrick O. Otieno, Penelope A. Phillips-Howard, and John Schneider. 2023. "Vaginal Microbial Network Analysis Reveals Novel Taxa Relationships among Adolescent and Young Women with Incident Sexually Transmitted Infection Compared with Those Remaining Persistently Negative over a 30-Month Period" Microorganisms 11, no. 8: 2035. https://doi.org/10.3390/microorganisms11082035
APA StyleMehta, S. D., Agingu, W., Zulaika, G., Nyothach, E., Bhaumik, R., Green, S. J., van Eijk, A. M., Otieno, F. O., Phillips-Howard, P. A., & Schneider, J. (2023). Vaginal Microbial Network Analysis Reveals Novel Taxa Relationships among Adolescent and Young Women with Incident Sexually Transmitted Infection Compared with Those Remaining Persistently Negative over a 30-Month Period. Microorganisms, 11(8), 2035. https://doi.org/10.3390/microorganisms11082035