
Identification and Molecular Characterization of Giant Liver Fluke (Fascioloides magna) Infection in European Fallow Deer (Dama dama) in Romania—First Report

 , , ,
, , ,  ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Coprological Examination
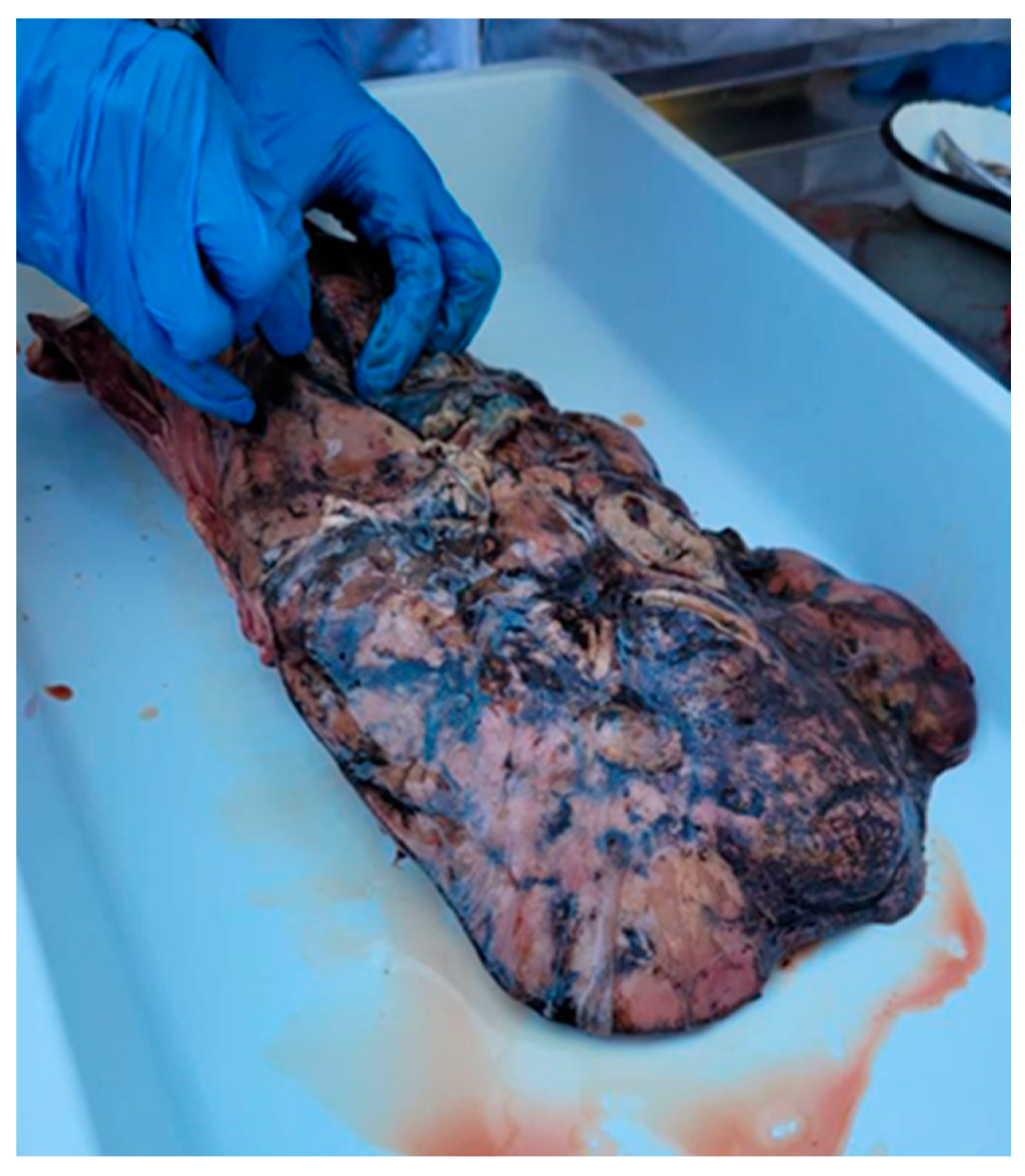
2.3. Necropsy Examination of the Liver
2.4. PCR Protocol
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Dărăbuş, G.; Oprescu, I.; Morariu, S.; Mederle, N.; Ilie, M.; Imre, M. Parazitologie și Boli Parazitare, 3rd ed.; Mirton: Timişoara, Romania, 2022. [Google Scholar]
- Feis, M.E.; Goedknegt, A.M.; Thieltges, D.W.; Buschbaum, C.; Wegner, K.M. Biological invasions and host–parasite coevolution: Different coevolutionary trajectories along separate parasite invasion fronts. Zoology 2016, 119, 366–374. [Google Scholar] [CrossRef]
- Konjević, D.; Bujanić, M.; Beck, A.; Beck, R.; Martinković, F.; Janicki, Z. First record of chronic Fascioloides magna infection in roe deer (Capreolus capreolus). Int. J. Parasitol. Parasites Wildl. 2021, 15, 173–176. [Google Scholar] [CrossRef] [PubMed]
- Ullrich, K. The presence of rare or little-known parasites of mammals and birds in Bohemia and Moravia (in German). Prag. Arch. Tiermed. Vergl. Pathol. 1930, 10, 19–43. [Google Scholar]
- Rajský, D.; Patus, A.; Bukovjan, K. Prvý nález Fascioloides magna (Bassi, 1875) na Slovensku. Slov. Vet. Čas. 1994, 19, 29–30. [Google Scholar]
- Rajský, D.; Čorba, J.; Varady, M.; Špakulová, M.; Cabadaj, R. Control of fascioloidosis (Fascioloides magna Bassi, 1875) in red deer and roe deer. Helminthology 2002, 39, 67–70. [Google Scholar]
- Rehbein, S.; Hamel, D.; Reindl, H.; Visser, M.; Pfister, K. Fascioloides magna and Ashworthius sidemi–two new parasites in wild ungulates in Germany. In Proceedings of the Europ Multicoll Parasi-tol (EMOP) XI, Program & Abstracts, Cluj-Napoca, Romania, 25–29 July 2012. [Google Scholar]
- Winkelmayer, R.; Prosl, H. Riesenleberegel-jetzt auch bei uns? Weidwerk 2001, 3, 42–44. [Google Scholar]
- Ursprung, J.; Joachim, A.; Prosl, H. Epidemiology and control of the giant liver fluke, Fascioloides magna, in a population of wild ungulates in the Danubian wetlands east of Vienna. Berl. Munch. Tierarztl. 2006, 119, 316–323. [Google Scholar]
- Vodnanský, M.; Rajský, D. Erfahrungen bei der Erforschung und Bekämpfung des Americanischen Rieseleberegels (Fascioloides magna) in der Nachbarländern Österreich. In Symposium Erforschung und Bekämpfung des neu in Österreich autrettenden Amerikanischen Reiseleberegels (Fascioloides magna); Niederosterreichischer Landesjagdverband: Wien, Austria, 2001; pp. 10–13. [Google Scholar]
- Majoros, G.; Sztojkov, V. (1994): Appearance of the American liver fluke Fascioloides magna (Bassi, 1875) (Trematoda: Fasciolata) in Hungary. Parasitol. Hung. 1994, 27, 27–38. [Google Scholar]
- Marinculić, A.; Džakula, N.; Janicki, Z.; Hardy, Z.; Lučinger, S.; Živičnjak, T. Appearance of American liver fluke (Fascioloides magna, Bassi, 1875) in Croatia—A case report. Vet. Arhiv 2002, 72, 319–325. [Google Scholar]
- Marinkovic, D.; Nesic, V. Zlatibor: Serbain Veterinary Society, Belgrade; Changes on the liver of fallow deer (Dama dama) caused by american giant liver fluke (Fascioloides magna). In Proceedings of the XX Veterinary Meeting of Serbia, Zlatibor, Serbia, 9–12 September 2008; pp. 298–309. [Google Scholar]
- Samuel, W.M.; Pybus, M.J.; Kocan, A.A. (Red.): Parasitic Diseases of Wild Mammals; Manson Publishing/The Veterinary Press, Iowa State University Press: London, UK, 2001; Volume 121, p. 560. [Google Scholar]
- Boggs, A.D.; DePerno, C.S.; Flowers, J.R. Range Expansion of Fascioloides magna in North Carolina. Southeast Nat. 2018, 17, 365–370. [Google Scholar] [CrossRef]
- Malcicka, M. Life history and biology of Fascioloides magna (Trematoda) and its native and exotic hosts. Ecol. Evol. 2015, 5, 1381–1397. [Google Scholar] [CrossRef] [PubMed]
- Karamon, J.; Larska, M.; Jasik, A.; Sell, B. First report of the giant liver fluke (Fascioloides magna) infection in farmed fallow deer (Dama dama) in Poland–pathomorphological changes and molecular identification. Bull. Vet. Inst. Pulawy. 2015, 59, 339–344. [Google Scholar] [CrossRef]
- Králová-Hromadová, I.; Bazsalovicsova, E.; Demiaszkiewicz, A.W. Molecular characterization of Fascioloides magna (Trematoda: Fasciolidae) from south-western Poland based on mitochondrial markers. Acta Parasitol. 2015, 60, 544–547. [Google Scholar] [CrossRef]
- Stiles, C.; Bujanić, M.; Martinković, F.; Šoštarić Zuckermann, I.C.; Konjević, D. Severe Pulmonary Fascioloidosis in a Wild Mouflon (Ovis Musimon)—A Case Report. Helminthologia 2021, 58, 394–399. [Google Scholar] [CrossRef] [PubMed]
- Mathieu, A.; Thacker, C.; Teske, I.; Jenkins, E.; Wagner, B.; Macbeth, B.; Raverty, S.; Pybus, M. Fascioloides magna in free-ranging rocky mountain bighorn sheep (Ovis canadensis). J. Wildl. Dis. 2022, 58, 592–598. [Google Scholar] [CrossRef] [PubMed]
- Pybus, M.J.; Butterworth, E.W.; Woods, J.G. An expanding population of the giant liver fluke (Fascioloides magna) in elk (Cervus canadensis) and other ungulates in Canada. J. Wildl. Dis. 2015, 51, 431–445. [Google Scholar] [CrossRef] [PubMed]
- Kuleš, J.; Lovrić, L.; Gelemanović, A.; Ljubić, B.B.; Rubić, I.; Bujanić, M.; Konjević, D. Complementary liver and serum protein profile in wild boars infected by the giant liver fluke Fascioloides magna using tandem mass tags quantitative approach. J. Proteomics. 2021, 247, 104332. [Google Scholar] [CrossRef]
- Foreyt, W.J.; Drew, M.L. Experimental infection of liver flukes, Fasciola hepatica and Fascioloides magna, in Bison (Bison bison). J. Wildl. Dis. 2010, 46, 283–286. [Google Scholar] [CrossRef]
- Konjević, D.; Bujanić, M.; Erman, V.; Gudan Kurilj, A.; Živičnjak, T.; Severin, K.; Tomić, S.; Martinković, F. New data on wild boar (Sus scrofa L.) a dead-end host for large American liver fl uke (Fascioloides magna). Helminthology 2017, 54, 77–80. [Google Scholar] [CrossRef]
- Leontovyč, R.; Košťáková, M.; Siegelová, V.; Melounová, K.; Pankrác, J.; Vrbová, K.; Horák, P.; Kašný, M. Highland cattle and Radix labiata, the hosts of Fascioloides magna. BMC Vet. Res. 2014, 10, 41. [Google Scholar] [CrossRef]
- Shury, T.K.; Pybus, M.J.; Nation, N.; Cool, N.L.; Rettie, W.J. Fascioloides magna in Moose (Alces alces) From Elk Island National Park, Alberta. Vet. Pathol. 2019, 56, 476–485. [Google Scholar] [CrossRef]
- Hasse, K.E.; Garner, M.M.; Knightly, F.A.; Sobotyk, C.; Luksovsky, J.L.; Verocai, G.G. Fatal Fascioloides magna in a lesser spot-nosed guenon (Cercopithecus petaurista). J. Zoo. Wildl. Med. 2021, 52, 1309–1313. [Google Scholar] [CrossRef] [PubMed]
- Špakulová., M.; Rajský, D.; Sokol, J.; Vodňanský, M. Giant Liver Fluke (Fascioloides magna), an Important Liver Parasite of Ruminants; Parpress: Brastislava, Slovakia, 2003. [Google Scholar]
- Chapman, D.; Chapman, N. Fallow Deer Their History, Distribution and Biology; Terence Dalton Lavenham: Suffolk, UK, 1975. [Google Scholar]
- Ordinul Ministrului Mediului, Apelor și Pădurilor nr. 1571/07.06.2022 Privind Aprobarea Cotelor de Recoltă Pentru Unele Specii de Faună de Interes Cinegetic. Available online: https://mmediu.ro/articol/ordinul-ministrului-mediului-apelor-si-padurilor-nr-1571 (accessed on 11 August 2023).
- Cotta, V.; Bodea, M. Vânatul României; Editura Agrosilvică: București, Romania, 1969. [Google Scholar]
- Sinclair, A.R.E.; Fryxell, J.M.; Caughley, G. Wildlife Ecology, Conservation, and Management, 2nd ed.; Sinclair, R.E., Fryxell, J.M., Eds.; BLACKWELL PUBLISHING © by Anthony: Canberra, Australia, 2006. [Google Scholar]
- Popovici, D.C.; Mederle, N.; Marin, A.M.; Moraru, M.M.F.; Albu-Kaya, M.G.; Ionescu, O. Evaluation of the parasitic load in fallow deer (Dama dama L.) from Arad County. Lucr. Științ. Med. Vet. Timișoara 2022, 55, 172–179. [Google Scholar]
- Hora, Ș.; Genchi, C.; Ferrari, N.; Morariu, S.; Mederle, N.; Dărăbuș, G. Frequency of gastrointestinal and pulmonary helminth infections in wild deer from western Romania. Vet Parasitol Reg. Stud. Rep. Vol. 2017, 8, 75–77. [Google Scholar] [CrossRef]
- Popovici, D.C.; Mederle, N.; Marin, A.M.; Moraru, M.M.F.; Ghilean, B.M.; Kaya, D.A.; Ionescu, O. Evaluation of endoparasitism in fallow deer (Dama dama L.) from Bihor county (Romania) hunting grounds. Lucr. Științ. Med. Vet. Timișoara 2023, 56, 213–219. [Google Scholar]
- Doniță, N.; Popescu, A.; Paucă-Comănescu, M.; Mihăilescu, S.; Biriș, I.A. Habitatele din România; Editura Tehnică Silvică: Timișoara, Romania, 2005. [Google Scholar]
- Zajac, A.M.; Conboy, G.A. Veterinary Clinical Parasitology, 8th ed.; John Willey & Sons, Ltd.: Hoboken, NJ, USA, 2012. [Google Scholar]
- Lotfy, W.M.; Brant, S.V.; Ashmawy, K.I.; Devkota, R.; Mkoji, G.M.; Loker, E.S. A molecular approach for identification of paramphistomes from Africa and Asia. Vet. Parasitol. 2010, 174, 234–240. [Google Scholar] [CrossRef] [PubMed]
- Cucher, M.; Carnevale, S.; Prepelitchi, L.; Labbé, J.; Wisnivesky-Colli, C. PCR diagnosis of Fasciola hepatica in field-collected Lymnaea columella and Lymnaea viatrix snails. Vet. Parasitol. 2006, 137, 74–82. [Google Scholar] [CrossRef] [PubMed]
- Deeeper, A.; Guignon, V.; Blanc, G.; Audic, S.; Buffet, S.; Chevenet, F.; Dufayard, J.F.; Guindon, S.; Lefort, V.; Lescot, M.; et al. Phylogeny.fr: Robust phylogenetic analysis for the non-specialist. Nucleic Acids Res. 2008, 36, 465–469. [Google Scholar] [CrossRef] [PubMed]
- Lemoine, F.; Correia, D.; Lefort, V.; Doppelt-Azeroual, O.; Mareuil, F.; Cohen-Boulakia, S.; Gascuel, O. NGPhylogeny.fr: New generation phylogenetic services for non-specialists. Nucleic Acids Res. 2019, 47, 260–265. [Google Scholar] [CrossRef]
- Letunic, I.; Bork, P. Interactive Tree of Life (iTOL) v5: An online tool for phylogenetic tree display and annotation. Nucleic Acids Res. 2021, 49, 293–296. [Google Scholar] [CrossRef]
- Novobilský, A.; Horácková, E.; Hirtová, L.; Modrý, D.; Koudela, B. The giant liver fluke Fascioloides magna (Bassi 1875) in cervids in the Czech Republic and potential of its spreading to Germany. Parasitol. Res. 2007, 100, 549–553. [Google Scholar] [CrossRef]
- Ursprung, J.; Prosl, H. The American Giant Liver Fluke (Fascioloides magna) in the Danube floodplain forests east of Vienna, Austria. Occurrence and control 2000–2010. Wien. Tierarztl. Monatsschrift. 2011, 98, 275–284. [Google Scholar]
- Sattmann, H.; Hörweg, C.; Gaub, L.; Feix, A.S.; Haider, M.; Walochnik, J.; Rabitsch, W.; Prosl, H. Wherefrom and whereabouts of an alien: The American liver fluke Fascioloides magna in Austria: An overview. Wien. Klin. Wochenschr. 2014, 126, 23–31. [Google Scholar] [CrossRef]
- Halász, T.; Tari, T.; Nagy, E.; Nagy, G.; Csivincsik, Á. Hatchability of Fascioloides magna Eggs in Cervids. Pathogens 2023, 12, 741. [Google Scholar] [CrossRef] [PubMed]
- Trailovic, S.M.; Marinkovic, D.; Kulisic, Z. Diagnosis and therapy of liver fluke (Fascioloides magna) infection in fallow deer (Dama dama) in Serbia. J. Wildl. Dis. 2016, 52, 319–326. [Google Scholar] [CrossRef] [PubMed]
- Rehbein, S.; Visser, M.; Hamel, D.; Reindl, H. Occurrence of the giant liver fluke, Fascioloides magna, in sympatric wild ungulates in one area in the Upper Palatinate Forest (northeastern Bavaria, Germany). Parasitol. Res. 2021, 120, 553–561. [Google Scholar] [CrossRef]
- Houszka, M.; Piekarska, J.; Podkowik, M.; Gorczykowski, M.; Bania, J. Morphology and molecular study of Fascioloides magna—a growing threat to cervids (Cervidae) in Poland. J. Vet. Res. 2016, 60, 435–439. [Google Scholar] [CrossRef]
- Juhásová, L.; Bazsalovicsová, E.; Králová-Hromadová, I.; Karamon, J. A genetic structure of novel population of Fascioloides magna from Poland, Podkarpackie Province, indicates an expanding second European natural focus of fascioloidosis. Acta Parasitol. 2016, 61, 790–795. [Google Scholar] [CrossRef]
- Demiaszkiewicz, A.W.; Pyziel, A.M.; Kuligowska, I.; Lachowicz, J. Fascioloides magna, a parasite of red deer in Lower Silesian Wilderness. Med. Weter. 2016, 72, 110–112. [Google Scholar]
- Demiaszkiewicz, A.W.; Kowalczyk, R.; Filip, K.J.; Pyziel, A.M. Fascioloides magna: A parasite of roe deer in Bory Zielonogórskie. Med. Weter. 2018, 74, 257–260. [Google Scholar] [CrossRef]
- Králová-Hromadová, I.; Bazsalovicsová, E.; Stefka, J.; Spakulová, M.; Vávrová, S.; Szemes, T.; Tkach, V.; Trudgett, A.; Pybus, M. Multiple origins of European populations of the giant liver fluke Fascioloides magna (Trematoda: Fasciolidae), a liver parasite of ruminants. Int. J. Parasitol. 2011, 41, 373–383. [Google Scholar] [CrossRef]
- Husch, C.; Sattmann, H.; Hörweg, C.; Ursprung, J.; Walochnik, J. Genetic homogeneity of Fascioloides magna in Austria. Vet. Parasitol. 2017, 243, 75–78. [Google Scholar] [CrossRef] [PubMed]
- Rajković-Janje, R.; Bosnić, S.; Rimac, D.; Gojmerac, T. The prevalence of American liver fluke Fascioloides magna (Bassi 1875) in red deer from Croatian hunting grounds. Eur. J. Wildl. Res. 2008, 54, 525–528. [Google Scholar] [CrossRef]
- Nagy, E.; Jócsák, I.; Csivincsik, Á.; Zsolnai, A.; Halász, T.; Nyúl, A.; Plucsinszki, Z.; Simon, T.; Szabó, S.; Turbók, J.; et al. Establishment of Fascioloides magna in a new region of Hungary: Case report. Parasitol. Res. 2018, 117, 3683–3687. [Google Scholar] [CrossRef]
- Sindičić, M.; Davinack, A.; Bujanić, M.; Bugarski, D.; Mirčeta, J.; Ferroglio, E.; Konjević, D. A new insight into genetic structure of Danube and Italian foci of fascioloidosis. Vet. Parasitol. 2023, 314, 109854. [Google Scholar] [CrossRef]
- Rehbein, S.; Visser, M. Sika Deer (Cervus nippon) are not “Dead-End Hosts” of the Giant Liver Fluke, Fascioloides magna (Bassi, 1875) Ward, 1917. J. Wildl. Dis. 2022, 58, 194–197. [Google Scholar] [CrossRef]
- Filip-Hutsch, K.; Pyziel-Serafin, A.M.; Hutsch, T.; Bulak, K.; Czopowicz, M.; Merta, D.; Kobielski, J.; Demiaszkiewicz, A.W. The Occurrence of Fascioloides Magna (Bassi, 1875) in the Wild Cervid Population in the Lower Silesian Wilderness-Epidemiological and Pathological Aspects. J. Vet. Res. 2022, 66, 381–387. [Google Scholar] [CrossRef] [PubMed]
- Plötz, C.; Rehbein, S.; Bamler, H.; Reindl, H.; Pfister, K.; Scheuerle, M.C. Fascioloides magna-epizootiology in a deer farm in Germany. Berl. Munch. Tierarztl. Wochenschr. 2015, 128, 177–182. [Google Scholar] [PubMed]
- Hirtová, L.; Modrý, D.; Faltýnková, A. Epizootiology of fascioloidosis in a fenced heard of fallow deer (Dama dama). Proceedings of the 12th Helminthological Days held at Dolní Ve4stonice (Czech Republic), May 5–8. Helminthology 2003, 40, 180. [Google Scholar]
- Florijancic, T.; Opacak, A.; Marinculic, A.; Janicki, Z.; Puskadija, Z. The impact of habitat on distribuion of giant liver fluke (Fascioloides magna) in Eastern Croatia. Cereal. Res. Commun. 2008, 36, 1543–1546. [Google Scholar]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Popovici, D.-C.; Dărăbuș, G.; Marin, A.-M.; Ionescu, O.; Moraru, M.M.F.; Imre, M.; Tîrziu, E.; Mederle, N. Identification and Molecular Characterization of Giant Liver Fluke (Fascioloides magna) Infection in European Fallow Deer (Dama dama) in Romania—First Report. Microorganisms 2024, 12, 527. https://doi.org/10.3390/microorganisms12030527
Popovici D-C, Dărăbuș G, Marin A-M, Ionescu O, Moraru MMF, Imre M, Tîrziu E, Mederle N. Identification and Molecular Characterization of Giant Liver Fluke (Fascioloides magna) Infection in European Fallow Deer (Dama dama) in Romania—First Report. Microorganisms. 2024; 12(3):527. https://doi.org/10.3390/microorganisms12030527
Chicago/Turabian StylePopovici, Dan-Cornel, Gheorghe Dărăbuș, Ana-Maria Marin, Ovidiu Ionescu, Maria Monica Florina Moraru, Mirela Imre, Emil Tîrziu, and Narcisa Mederle. 2024. "Identification and Molecular Characterization of Giant Liver Fluke (Fascioloides magna) Infection in European Fallow Deer (Dama dama) in Romania—First Report" Microorganisms 12, no. 3: 527. https://doi.org/10.3390/microorganisms12030527
APA StylePopovici, D. -C., Dărăbuș, G., Marin, A. -M., Ionescu, O., Moraru, M. M. F., Imre, M., Tîrziu, E., & Mederle, N. (2024). Identification and Molecular Characterization of Giant Liver Fluke (Fascioloides magna) Infection in European Fallow Deer (Dama dama) in Romania—First Report. Microorganisms, 12(3), 527. https://doi.org/10.3390/microorganisms12030527