


A Retrospective Analysis of Salmonella Isolates across 11 Animal Species (1982–1999) Led to the First Identification of Chromosomally Encoded blaSCO-1 in the USA

 , ,
, ,  , , and
, , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Isolates
2.2. Antimicrobial Susceptibility Testing
2.3. Whole-Genome Sequencing (WGS) and Genome Analysis
2.4. Genotypic and Phenotypic Correlation Analysis
2.5. Genome Visualization and Analysis
3. Results
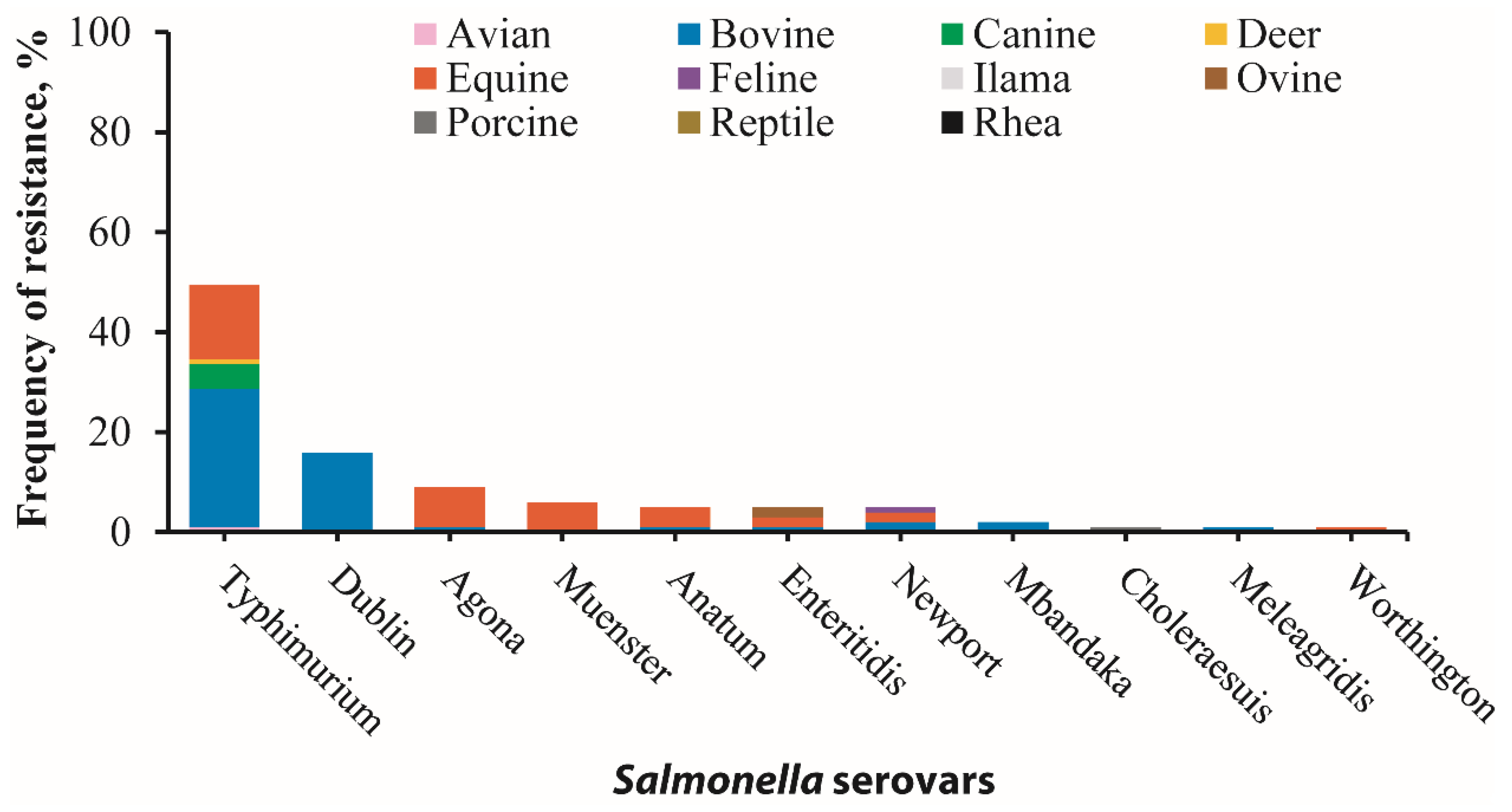
3.1. Identification of 36 Salmonella Serovars in 11 Animal Species
3.2. Salmonella Antimicrobial Resistance (AMR) Phenotypes
3.3. Antimicrobial Resistance Genes in Salmonella Isolates
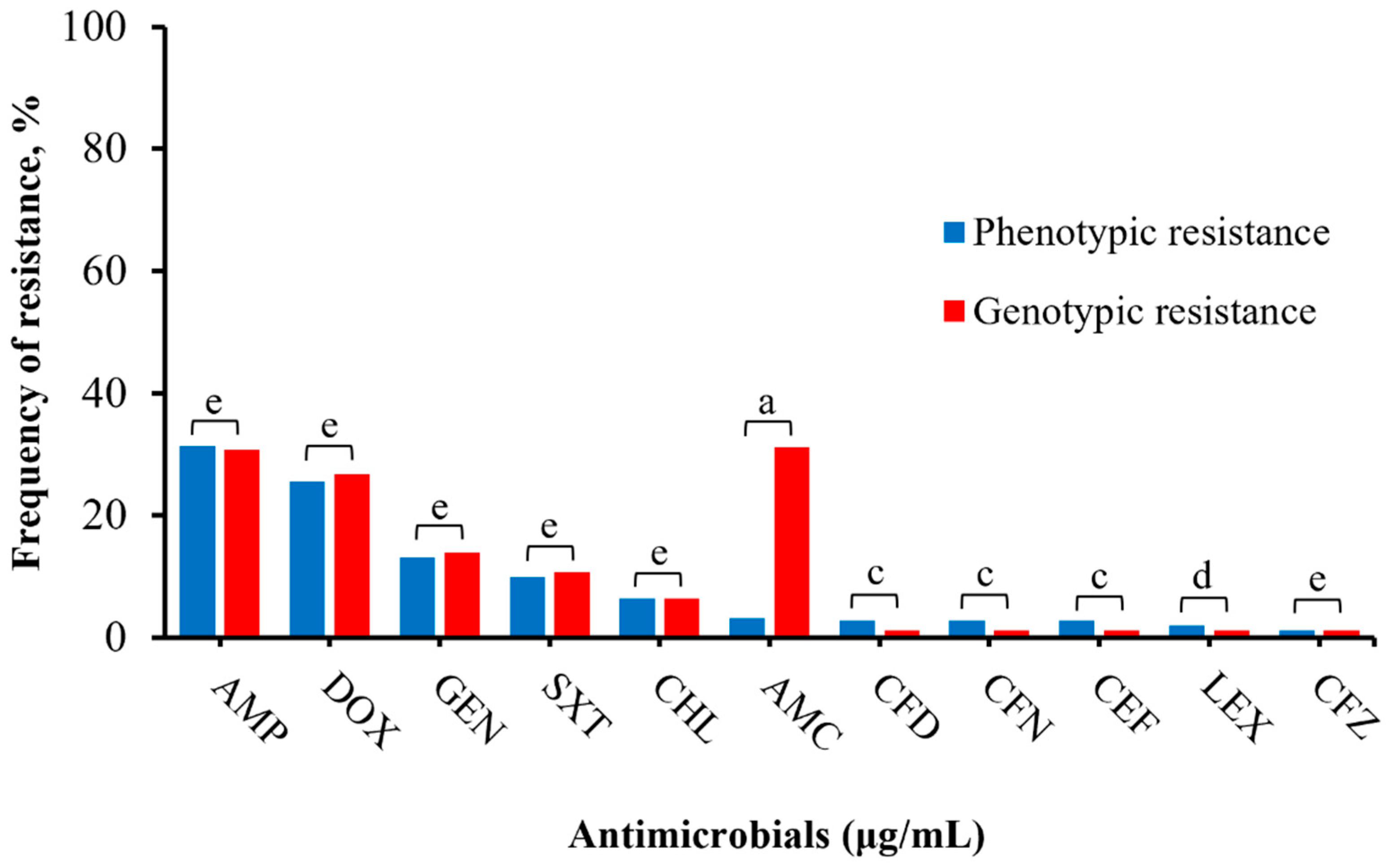
3.4. Phenotypic and Genotypic Resistance Correlation in Salmonella Isolates
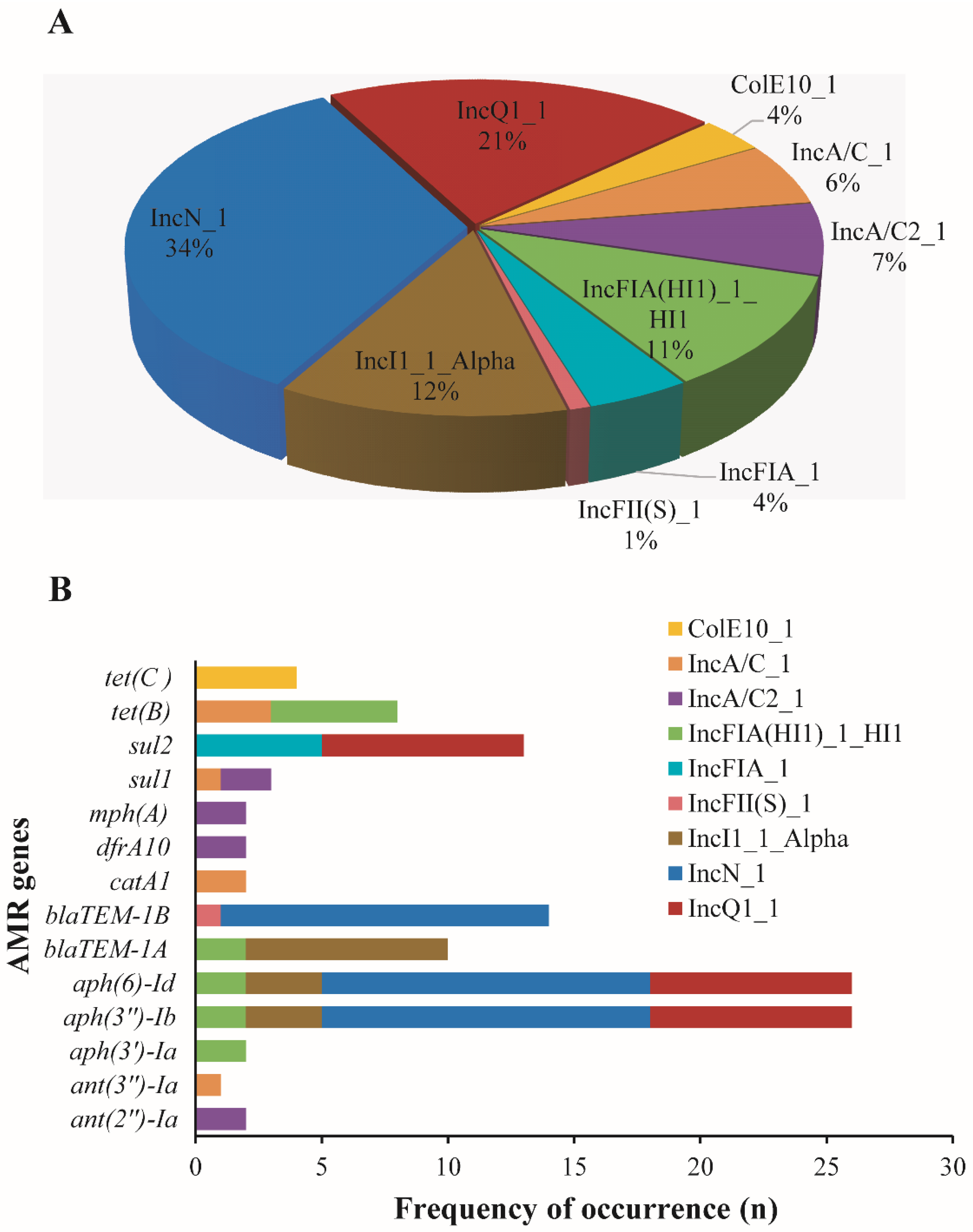
3.5. Plasmid-Mediated AMR Genes
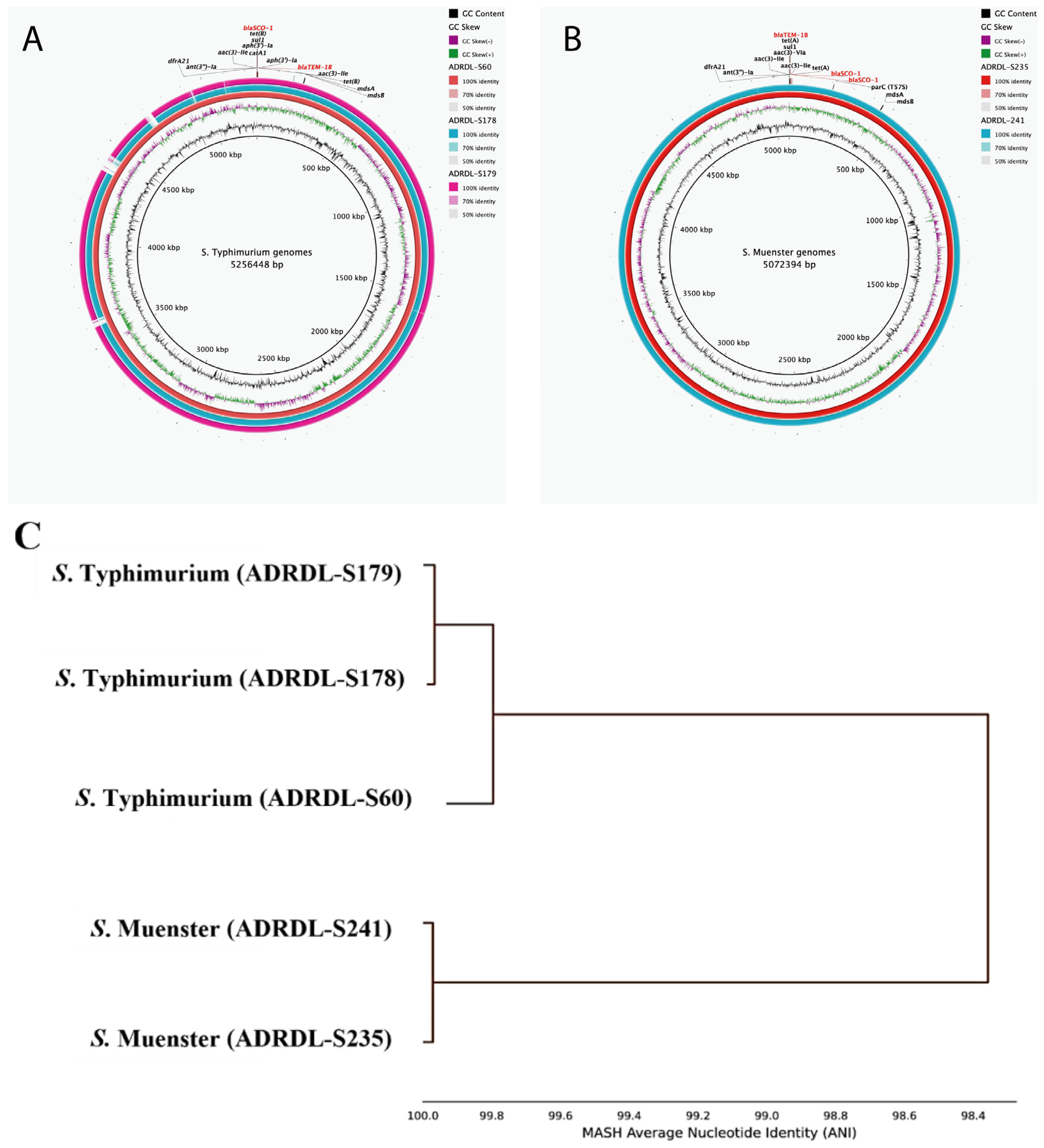
3.6. Characteristics of blaSCO-1 Positive Isolates
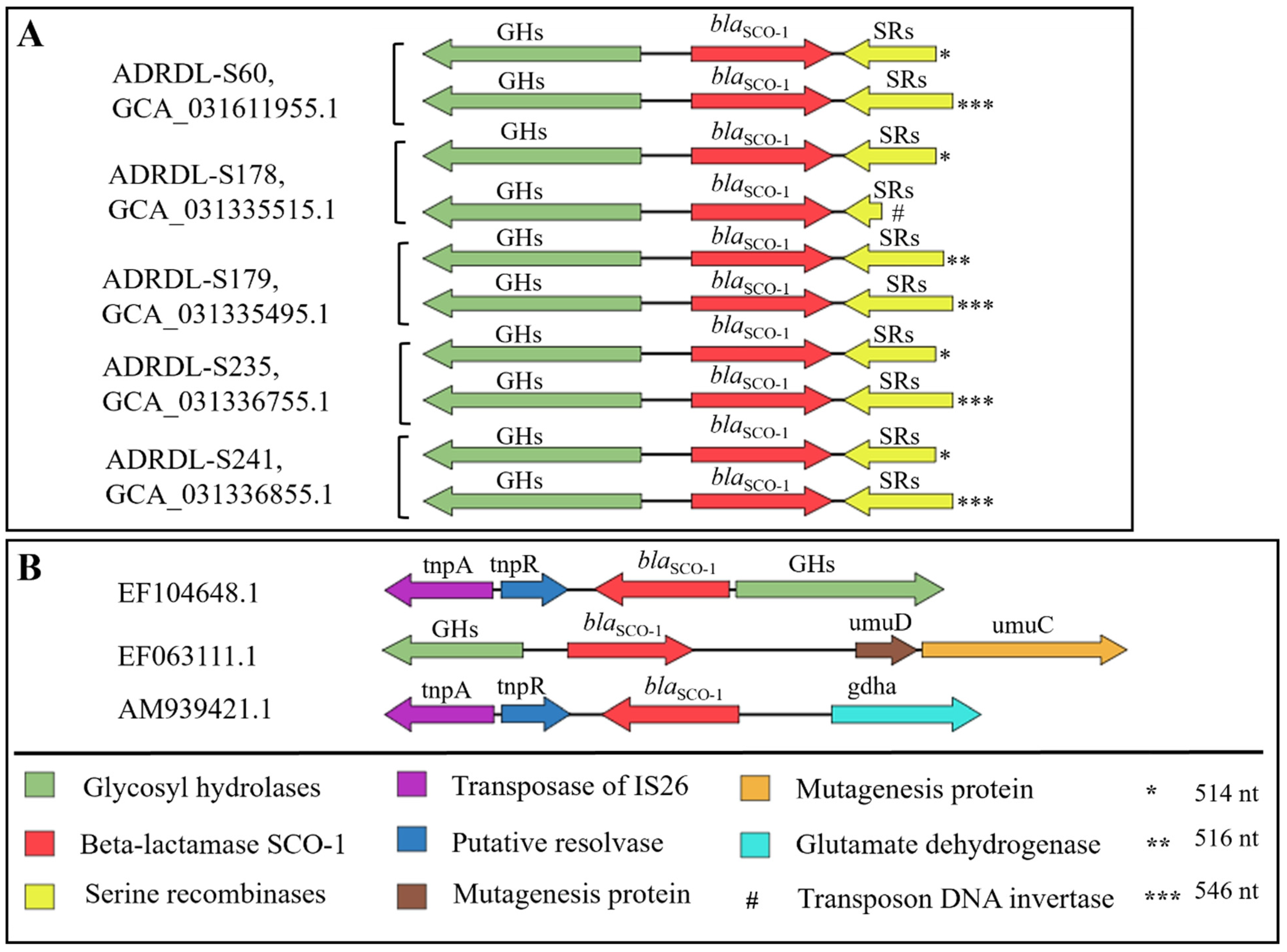
3.7. Comparison of the Genetic Environment of blaSCO-1 Gene
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Syed Abu Thahir, S.; Rajendiran, S.; Shaharudin, R.; Veloo, Y. Multidrug-resistant Salmonella species and their mobile genetic elements from poultry farm environments in Malaysia. Antibiotics 2023, 12, 1330. [Google Scholar] [CrossRef] [PubMed]
- Kijima, M.; Shirakawa, T.; Uchiyama, M.; Kawanishi, M.; Ozawa, M.; Koike, R. Trends in the serovar and antimicrobial resistance in clinical isolates of Salmonella enterica from cattle and pigs between 2002 and 2016 in Japan. J. Appl. Microbiol. 2019, 127, 1869–1875. [Google Scholar] [CrossRef] [PubMed]
- Centers for Disease Control and Prevention (CDC). CDC Yellow Book: Salmonellosis, Nontyphoidal. 2024. Available online: https://wwwnc.cdc.gov/travel/yellowbook/2024/infections-diseases/salmonellosis-nontyphoidal (accessed on 25 February 2024).
- Eng, S.-K.; Pusparajah, P.; Ab Mutalib, N.-S.; Ser, H.-L.; Chan, K.-G.; Lee, L.-H. Salmonella: A review on pathogenesis, epidemiology and antibiotic resistance. Front. Life Sci. 2015, 8, 284–293. [Google Scholar] [CrossRef]
- Li, C.; Tyson, G.H.; Hsu, C.-H.; Harrison, L.; Strain, E.; Tran, T.-T.; Tillman, G.E.; Dessai, U.; McDermott, P.F.; Zhao, S. Long-read sequencing reveals evolution and acquisition of antimicrobial resistance and virulence genes in Salmonella enterica. Front. Microbiol. 2021, 12, 777817. [Google Scholar] [CrossRef] [PubMed]
- Yao, Y.; Maddamsetti, R.; Weiss, A.; Ha, Y.; Wang, T.; Wang, S.; You, L. Intra- and interpopulation transposition of mobile genetic elements driven by antibiotic selection. Nat. Ecol. Evol. 2022, 6, 555–564. [Google Scholar] [CrossRef] [PubMed]
- Jia, C.; Wang, Z.; Huang, C.; Teng, L.; Zhou, H.; An, H.; Liao, S.; Liu, Y.; Huang, L.; Tang, B.; et al. Mobilome-driven partitions of the resistome in Salmonella. mSystems 2023, 8, e0088323. [Google Scholar] [CrossRef] [PubMed]
- Bradford, P.A. Extended-spectrum beta-lactamases in the 21st century: Characterization, epidemiology, and detection of this important resistance threat. Clin. Microbiol. Rev. 2001, 14, 933–951. [Google Scholar] [CrossRef]
- Vale, A.P.; Shubin, L.; Cummins, J.; Leonard, F.C.; Barry, G. Detection of blaOXA-1, blaTEM-1, and virulence factors in E. coli isolated from seals. Front. Vet. Sci. 2021, 8, 583759. [Google Scholar] [CrossRef]
- Mather, A.E.; Reid, S.W.; Maskell, D.J.; Parkhill, J.; Fookes, M.C.; Harris, S.R.; Brown, D.J.; Coia, J.E.; Mulvey, M.R.; Gilmour, M.W.; et al. Distinguishable epidemics of multidrug-resistant Salmonella Typhimurium DT104 in different hosts. Science 2013, 341, 1514–1517. [Google Scholar] [CrossRef]
- Shivani, C.; Abha, K.; Alka, G.; Sampat, N. Comparative genome analysis of three pathogenic strains of E. coli, Salmonella, and Shigella. Int. J. Sch. Res. Rev. 2015, 4, 68–80. [Google Scholar]
- Katiyar, A.; Sharma, P.; Dahiya, S.; Singh, H.; Kapil, A.; Kaur, P. Genomic profiling of antimicrobial resistance genes in clinical isolates of Salmonella Typhi from patients infected with Typhoid fever in India. Sci. Rep. 2020, 10, 8299. [Google Scholar] [CrossRef] [PubMed]
- McDermott, P.F.; Tyson, G.H.; Kabera, C.; Chen, Y.; Li, C.; Folster, J.P.; Ayers, S.L.; Lam, C.; Tate, H.P.; Zhao, S. Whole-genome sequencing for detecting antimicrobial resistance in nontyphoidal Salmonella. Antimicrob. Agents Chemother. 2016, 60, 5515–5520. [Google Scholar] [CrossRef] [PubMed]
- Gong, J.; Zeng, X.; Zhang, P.; Zhang, D.; Wang, C.; Lin, J. Characterization of the emerging multidrug-resistant Salmonella enterica serovar Indiana strains in China. Emerg. Microbes Infect. 2019, 8, 29–39. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Kang, X.; Ed-Dra, A.; Zhou, X.; Jia, C.; Müller, A.; Liu, Y.; Kehrenberg, C.; Yue, M. Genome-based assessment of antimicrobial resistance and virulence potential of isolates of non-Pullorum/Gallinarum Salmonella serovars recovered from dead poultry in China. Microbiol. Spectr. 2022, 10, e0096522. [Google Scholar] [CrossRef] [PubMed]
- Elbediwi, M.; Tang, Y.; Yue, M. Genomic characterization of ESBL-producing Salmonella Thompson isolates harboring mcr-9 from dead chick embryos in China. Vet. Microbiol. 2023, 278, 109634. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Biswas, S.; Paudyal, N.; Pan, H.; Li, X.; Fang, W.; Yue, M. Antibiotic resistance in Salmonella Typhimurium isolates recovered from the food chain through National Antimicrobial Resistance Monitoring System between 1996 and 2016. Front. Microbiol. 2019, 10, 985. [Google Scholar] [CrossRef] [PubMed]
- Clinical and Laboratory Standards Institute (CLSI). Performance Standards for Antimicrobial Susceptibility Testing, 30th ed.; CLSI Supplement M100; CLSI: Wayne, PA, USA, 2020. [Google Scholar]
- Clinical and Laboratory Standards Institute (CLSI). Performance Standards for Antimicrobial Susceptibility Testing, 33rd ed.; CLSI Supplement M100; CLSI: Wayne, PA, USA, 2023. [Google Scholar]
- National Antimicrobial Resistance Monitoring System for Enteric Bacteria (NARMS). 2019. Available online: https://www.cdc.gov/narms/antibiotics-tested.html (accessed on 25 February 2024).
- IDEXX. Diagnostic Update: Microbiology Guide to Interpreting Minimum Inhibitory Concentration (MIC). 2019. Available online: https://www.idexx.com/files/microbiology-guide-interpreting-mic.pdf (accessed on 25 February 2024).
- Feldgarden, M.; Brover, V.; Haft, D.H.; Prasad, A.B.; Slotta, D.J.; Tolstoy, I.; Tyson, G.H.; Zhao, S.; Hsu, C.H.; McDermott, P.F.; et al. Validating the AMRFinder tool and resistance gene database by using antimicrobial resistance genotype-phenotype correlations in a collection of isolates. Antimicrob. Agents Chemother. 2019, 63, e00483-19. [Google Scholar] [CrossRef]
- Florensa, A.F.; Kaas, R.S.; Clausen, P.T.L.C.; Aytan-Aktug, D.; Aarestrup, F.M. ResFinder—An open online resource for identification of antimicrobial resistance genes in next-generation sequencing data and prediction of phenotypes from genotypes. Microb. Genom. 2022, 8, 000748. [Google Scholar] [CrossRef]
- Seemann, T. ABRicate: Mass Screening of Contigs for Antimicrobial Resistance Genes. 2016. Available online: https://github.com/tseemann/abricate (accessed on 25 February 2024).
- Carattoli, A.; Zankari, E.; García-Fernández, A.; Voldby Larsen, M.; Lund, O.; Villa, L.; Møller Aarestrup, F.; Hasman, H. In silico detection and typing of plasmids using PlasmidFinder and plasmid multilocus sequence typing. Antimicrob. Agents Chemother. 2014, 58, 3895–3903. [Google Scholar] [CrossRef]
- Khezri, A.; Avershina, E.; Ahmad, R. Plasmid identification and plasmid-mediated antimicrobial gene detection in Norwegian isolates. Microorganisms 2020, 9, 52. [Google Scholar] [CrossRef]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Chitsaz, F.; Derbyshire, M.K.; Gonzales, N.R.; Gwadz, M.; Lu, S.; Marchler, G.H.; Song, J.S.; Thanki, N.; Yamashita, R.A.; et al. The conserved domain database in 2023. Nucleic Acids Res. 2023, 51, D384–D388. [Google Scholar] [CrossRef] [PubMed]
- Scarpellini, G. Idostatistics: Cohen’s Kappa Free Calculator. 2022. Available online: https://idostatistics.com/cohen-kappa-free-calculator/ (accessed on 25 February 2024).
- Landis, J.R.; Koch, G.G. The measurement of observer agreement for categorical data. Biometrics 1977, 33, 159–174. [Google Scholar] [CrossRef] [PubMed]
- Gargano, V.; Gambino, D.; Migliore, S.; Vitale, M.; Sciortino, S.; Costa, A.; Vicari, D. Can human handling increase the presence of multidrug resistance (MDR) in Salmonella spp. isolated from food sources? Microorganisms 2021, 9, 2018. [Google Scholar] [CrossRef] [PubMed]
- Galán-Relaño, Á.; Valero Díaz, A.; Huerta Lorenzo, B.; Gómez-Gascón, L.; Mena Rodríguez, M.Á.; Carrasco Jiménez, E.; Pérez Rodríguez, F.; Astorga Márquez, R.J. Salmonella and salmonellosis: An update on public health implications and control strategies. Animals 2023, 13, 3666. [Google Scholar] [CrossRef] [PubMed]
- Tollefson, L.; Fedorka-Cray, P.J.; Angulo, F.J. Public health aspects of antibiotic resistance monitoring in the USA. Acta Vet. Scand. Suppl. 1999, 92, 67–75. [Google Scholar] [PubMed]
- Smith, S.M.; Palumbo, P.E.; Edelson, P.J. Salmonella strains resistant to multiple antibiotics: Therapeutic implications. Pediatr. Infect Dis. 1984, 3, 455–460. [Google Scholar] [CrossRef] [PubMed]
- Tadesse, D.A.; Singh, A.; Zhao, S.; Bartholomew, M.; Womack, N.; Ayers, S.; Fields, P.I.; McDermott, P.F. Antimicrobial resistance in Salmonella in the United States from 1948 to 1995. Antimicrob. Agents Chemother. 2016, 60, 2567–2571. [Google Scholar] [CrossRef]
- Cohen, M.L.; Tauxe, R.V. Drug-resistant Salmonella in the United States: An epidemiologic perspective. Science 1986, 234, 964–969. [Google Scholar] [CrossRef]
- Zamudio, R.; Boerlin, P.; Beyrouthy, R.; Madec, J.Y.; Schwarz, S.; Mulvey, M.R.; Zhanel, G.G.; Cormier, A.; Chalmers, G.; Bonnet, R.; et al. Dynamics of extended-spectrum cephalosporin resistance genes in Escherichia coli from Europe and North America. Nat. Commun. 2022, 13, 7490. [Google Scholar] [CrossRef]
- Yin, X.; Dudley, E.G.; Pinto, C.N.; M’ikanatha, N.M. Fluoroquinolone sales in food animals and quinolone resistance in non-typhoidal Salmonella from retail meats: United States, 2009–2018. J. Glob. Antimicrob. Resist. 2022, 29, 163–167. [Google Scholar] [CrossRef]
- Galarce, N.; Arriagada, G.; Sánchez, F.; Escobar, B.; Miranda, M.; Matus, S.; Vilches, R.; Varela, C.; Zelaya, C.; Peralta, J.; et al. Phenotypic and genotypic antimicrobial resistance in Escherichia coli strains isolated from household dogs in Chile. Front. Vet. Sci. 2023, 10, 1233127. [Google Scholar] [CrossRef]
- Lee, L.A.; Puhr, N.D.; Maloney, E.K.; Bean, N.H.; Tauxe, R.V. Increase in antimicrobial-resistant Salmonella infections in the United States, 1989–1990. J. Infect Dis. 1994, 170, 128–134. [Google Scholar] [CrossRef]
- Otto, S.J.G.; Ponich, K.L.; Cassis, R.; Goertz, C.; Peters, D.; Checkley, S.L. Antimicrobial resistance of bovine Salmonella enterica ssp. enterica isolates from the Alberta Agriculture and Forestry Disease Investigation Program (2006–2014). Can. Vet. J. 2018, 59, 1195–1201. [Google Scholar]
- Spier, S.J. Salmonellosis. Vet. Clin. North Equine Pract. 1993, 9, 385–397. [Google Scholar] [CrossRef]
- Alzahrani, K.O.; AL-Reshoodi, F.M.; Alshdokhi, E.A.; Alhamed, A.S.; Al Hadlaq, M.A.; Mujallad, M.I.; Mukhtar, L.E.; Alsufyani, A.T.; Alajlan, A.A.; Al Rashidy, M.S.; et al. Antimicrobial resistance and genomic characterization of Salmonella enterica isolates from chicken meat. Front. Microbiol. 2023, 14, 1104164. [Google Scholar] [CrossRef]
- Webber, M.A.; Piddock, L.J.V. The importance of efflux pumps in bacterial antibiotic resistance. J. Antimicrob. Chemother. 2003, 51, 9–11. [Google Scholar] [CrossRef] [PubMed]
- Maunsell, T.J.; Nguyen, S.; El Garach, F.; Miossec, C.; Cuinet, E.; Woehrlé, F.; Fanning, S.; Niedziela, D.A. A study of the correlation between phenotypic antimicrobial susceptibility testing methods and the associated genotypes determined by whole genome sequencing for a collection of Escherichia coli of bovine origin. bioRxiv. 2021. [Google Scholar] [CrossRef]
- Papagiannitsis, C.C.; Loli, A.; Tzouvelekis, L.S.; Tzelepi, E.; Arlet, G.; Miriagou, V. SCO-1, a novel plasmid-mediated class A β-lactamase with carbenicillinase characteristics from Escherichia coli. Antimicrob. Agents Chemother. 2007, 51, 2185–2188. [Google Scholar] [CrossRef]
- Poirel, L.; Corvec, S.; Rapoport, M.; Mugnier, P.; Petroni, A.; Pasteran, F.; Faccone, D.; Galas, M.; Drugeon, H.; Cattoir, V.; et al. Identification of the novel narrow-spectrum beta-lactamase SCO-1 in Acinetobacter spp. from Argentina. Antimicrob. Agents Chemother. 2007, 51, 2179–2184. [Google Scholar] [CrossRef] [PubMed]
- Ktari, S.; Arlet, G.; Verdet, C.; Jaoua, S.; Kachrid, A.; Ben Redjeb, S.; Mahjoubi-Rhimi, F.; Hammami, A. Molecular epidemiology and genetic environment of acquired bla ACC-1 in Salmonella enterica serotype Livingstone causing a large nosocomial outbreak in Tunisia. Microbial Drug Resistance (Larchmont, N.Y.). Microb. Drug Resist. 2009, 15, 279–286. [Google Scholar] [CrossRef] [PubMed]
- Jin, W.; Wachino, J.-I.; Kimura, K.; Yamada, K.; Arakawa, Y. New plasmid-mediated aminoglycoside 6′-N-acetyltransferase, AAC(6′)-Ian, and ESBL, TLA-3, from a Serratia marcescens clinical isolate. J. Antimicrob. Chemother. 2015, 70, 1331–1337. [Google Scholar] [CrossRef] [PubMed]
- Ruppé, E.; Cherkaoui, A.; Wagner, N.; La Scala, G.C.; Beaulieu, J.-Y.; Girard, M.; Frey, J.; Lazarevic, V.; Schrenzel, J. In vivo selection of a multidrug-resistant Aeromonas salmonicida during medicinal leech therapy. New Microbes New Infect. 2018, 21, 23–27. [Google Scholar] [CrossRef] [PubMed]
- Opazo-Capurro, A.; Higgins, P.G.; Wille, J.; Seifert, H.; Cigarroa, C.; González-Muñoz, P.; Quezada-Aguiluz, M.; Domínguez-Yévenes, M.; Bello-Toledo, H.; Vergara, L.; et al. Genetic features of Antarctic Acinetobacter radioresistens strain A154 harboring multiple antibiotic-resistance genes. Front. Cell Infect. Microbiol. 2019, 9, 328. [Google Scholar] [CrossRef]
- Debergh, H.; Maex, M.; Garcia-Graells, C.; Boland, C.; Saulmont, M.; Van Hoorde, K.; Saegerman, C. First Belgian report of ertapenem resistance in an ST11 Klebsiella pneumoniae strain isolated from a dog carrying blaSCO-1 and blaDHA-1 combined with permeability defects. Antibiotics 2022, 11, 1253. [Google Scholar] [CrossRef]
- Monte, D.F.; Lincopan, N.; Berman, H.; Cerdeira, L.; Keelara, S.; Thakur, S.; Fedorka-Cray, P.J.; Landgraf, M. Genomic features of high-priority Salmonella enterica serovars circulating in the food production chain, Brazil, 2000–2016. Sci. Rep. 2019, 9, 11058. [Google Scholar] [CrossRef]
- Papagiannitsis, C.C.; Tzouvelekis, L.S.; Kotsakis, S.D.; Tzelepi, E.; Miriagou, V. Sequence of pR3521, an IncB plasmid from Escherichia coli encoding ACC-4, SCO-1, and TEM-1 beta-lactamases. Antimicrob. Agents Chemother. 2011, 55, 376–381. [Google Scholar] [CrossRef] [PubMed]
- Johnson, R.C. Site-specific DNA inversion by serine recombinases. Microbiol. Spectr. 2015, 3, 1–36. [Google Scholar] [CrossRef]
- Nicolas, E.; Lambin, M.; Dandoy, D.; Galloy, C.; Nguyen, N.; Oger, C.A.; Hallet, B. The Tn3-family of replicative transposons. Microbiol. Spectr. 2015, 3, 693–726. [Google Scholar] [CrossRef]
- Centers for Disease Control and Prevention (CDC). Outbreak of Salmonella Infections Linked to Pet Bearded Dragons. 2020. Available online: https://www.cdc.gov/Salmonella/muenster-10-20/index.html (accessed on 25 February 2024).
- Patel, K.; Stapleton, G.S.; Trevejo, R.T.; Tellier, W.T.; Higa, J.; Adams, J.K.; Hernandez, S.M.; Sanchez, S.; Nemeth, N.M.; Debess, E.E.; et al. Human salmonellosis outbreak linked to Salmonella Typhimurium epidemic in wild songbirds, United States, 2020–2021. Emerg. Infect. Dis. 2023, 29, 2298–2306. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antimicrobial Class | AMG | Number of Isolates Harboring the AMG |
|---|---|---|
| Aminoglycosides | aph(6)-Ic | 16 |
| aph(6)-Id | 49 | |
| aac(3)-Via | 4 | |
| aph(3′)-IIa | 16 | |
| aadA2 | 17 | |
| aadA15 | 1 | |
| aac(3)-IId | 11 | |
| aac(3)-IIe | 5 | |
| ant(2″)-Ia | 18 | |
| aph(3′)-Ia | 55 | |
| aadA7 | 1 | |
| aph(3″)-Ib | 52 | |
| ant(3″)-Ia | 44 | |
| Beta-lactams | blaTEM-1A | 8 |
| blaTEM-1B | 71 | |
| blaCARB-2 | 1 | |
| blaCMY-2 | 3 | |
| blaOXA-2 | 1 | |
| blaSCO-1 | 5 | |
| Tetracycline | tet(A) | 39 |
| tet(B) | 24 | |
| tet(C) | 3 | |
| tet(G) | 1 | |
| Phenicol | catA1 | 12 |
| cmlA1 | 1 | |
| floR | 4 | |
| Sulfonamide | sul1 | 66 |
| sul2 | 22 | |
| Trimethoprim | dfrA12 | 9 |
| dfrA21 | 7 | |
| dfrA10 | 11 | |
| Quinolone | gyrA (S83F) | 2 |
| parC (T57S) | 128 | |
| Fosfomycin | fosA7 | 23 |
| Macrolide | mph(A) | 12 |
| Bleomycin | ble | 16 |
| Multi-drug efflux | mdsA | 249 |
| mdsB | 249 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Iduu, N.V.; Raiford, D.; Conley, A.; Scaria, J.; Nelson, J.; Ruesch, L.; Price, S.; Yue, M.; Gong, J.; Wei, L.; et al. A Retrospective Analysis of Salmonella Isolates across 11 Animal Species (1982–1999) Led to the First Identification of Chromosomally Encoded blaSCO-1 in the USA. Microorganisms 2024, 12, 528. https://doi.org/10.3390/microorganisms12030528
Iduu NV, Raiford D, Conley A, Scaria J, Nelson J, Ruesch L, Price S, Yue M, Gong J, Wei L, et al. A Retrospective Analysis of Salmonella Isolates across 11 Animal Species (1982–1999) Led to the First Identification of Chromosomally Encoded blaSCO-1 in the USA. Microorganisms. 2024; 12(3):528. https://doi.org/10.3390/microorganisms12030528
Chicago/Turabian StyleIduu, Nneka Vivian, Donna Raiford, Austin Conley, Joy Scaria, Julie Nelson, Laura Ruesch, Stuart Price, Min Yue, Jiansen Gong, Lanjing Wei, and et al. 2024. "A Retrospective Analysis of Salmonella Isolates across 11 Animal Species (1982–1999) Led to the First Identification of Chromosomally Encoded blaSCO-1 in the USA" Microorganisms 12, no. 3: 528. https://doi.org/10.3390/microorganisms12030528
APA StyleIduu, N. V., Raiford, D., Conley, A., Scaria, J., Nelson, J., Ruesch, L., Price, S., Yue, M., Gong, J., Wei, L., & Wang, C. (2024). A Retrospective Analysis of Salmonella Isolates across 11 Animal Species (1982–1999) Led to the First Identification of Chromosomally Encoded blaSCO-1 in the USA. Microorganisms, 12(3), 528. https://doi.org/10.3390/microorganisms12030528