Microorganisms in Fermented Apple Beverages: Current Knowledge and Future Directions

 , ,
, ,
Abstract
:1. Introduction
2. Microbial Diversity: From Apple to Cider
2.1. Yeast and Mold Diversity
2.2. Bacterial Diversity
2.3. Factors Influencing Microbial Diversity
3. Microbial Contribution to Cider Organoleptic Quality
3.1. Yeast Contribution
3.2. Bacterial Contribution
4. Safety Assessment of Fermented Apple Beverages
4.1. Biogenic Amines
4.2. Mycotoxins
4.3. Pathogens
5. Functional Improvement of Apple Fermented Beverages
5.1. Control of the Microbial Ecosystem to Improve or Modulate Cider Quality
5.1.1. Rational Design of Starter Cultures
5.1.2. Control of the Fermentation Process Parameters
5.1.3. Control of Cider Quality by LAB
5.2. Health Benefits of Apple Fermented Beverages
6. Conclusions
Conflicts of Interest
References
- Lea, A.G.H. Cidermaking. In Fermented Beverage Production; Lea, A.G.H., Piggott, J.R., Eds.; Kluwer Academic/Plenum Publishers: New York, NY, USA, 1995; pp. 66–96. ISBN 978-1-4757-5216-8. [Google Scholar]
- Nogueira, A.; Wosiacki, G. Apple Cider Fermentation. In Handbook of Plant-Based Fermented Food and Beverage Technology; Hui, Y.H., Ed.; CRC Press: Boca Raton, FL, USA, 2012; pp. 209–235. ISBN 978-1-4398-4904-0. [Google Scholar]
- Jarvis, B. CIDER (CYDER; HARD CIDER): The Product and its Manufacture. In Encyclopedia of Food Sciences and Nutrition, 2nd ed.; Caballero, B., Ed.; Academic Press: Oxford, UK, 2003; pp. 1312–1318. ISBN 978-0-12-227055-0. [Google Scholar]
- AICV—Publications. Available online: http://www.aicv.org/pages/aicv/publications.html (accessed on 18 July 2017).
- Richards, A.; Morgan, J. The Book of Apples, 1st ed.; Ebury Press: London, UK, 1993; ISBN 978-0-09-177759-3. [Google Scholar]
- Watson, B. Cider Styles and Traditions. In Cider, Hard and Sweet: History, Traditions, and Making Your Own, 3rd ed.; The Countryman Press: Woodstock, Vermont, 2013; pp. 95–108. ISBN 978-1-58157-689-4. [Google Scholar]
- Downing, D.L. Apple cider. In Processed Apple Products; Downing Donald L.: New York, NY, USA, 1989; pp. 169–188. ISBN 978-1-4684-8225-6. [Google Scholar]
- National Apple Museum. Available online: http://www.nationalapplemuseum.com/appleciderandmore.html (accessed on 6 March 2017).
- Coton, E.; Coton, M.; Guichard, H. Cider (Cyder; Hard Cider): The Product and Its Manufacture. In Encyclopedia of Food and Health; Academic Press: Oxford, UK, 2016; pp. 119–128. ISBN 978-0-12-384953-3. [Google Scholar]
- Boulongne, I. La Vie Paysanne Autrefois. Available online: http://viepaysanneautrefois.free.fr/ (accessed on 6 March 2017).
- L’histoire du Cidre en France. Available online: http://www.lalittorale.fr/lhistoire-du-cidre-en-france/ (accessed on 12 January 2017).
- Jarvis, B.; Lea, A.G.H. Sulphite binding in ciders. Int. J. Food Sci. Technol. 2000, 35, 113–127. [Google Scholar] [CrossRef]
- Sánchez, A.; Rodríguez, R.; Coton, M.; Coton, E.; Herrero, M.; García, L.A.; Díaz, M. Population dynamics of lactic acid bacteria during spontaneous malolactic fermentation in industrial cider. Food Res. Int. 2010, 43, 2101–2107. [Google Scholar] [CrossRef]
- Beech, F.W. 5—Yeasts in Cider-Making. In The Yeasts, 2nd ed.; Rose, A.H., Harrison, J.S., Eds.; Academic Press: San Diego, CA, USA, 1993; pp. 169–213. ISBN 978-0-12-596415-9. [Google Scholar]
- Handelsman, J. Metagenetics: Spending our inheritance on the future. Microb. Biotechnol. 2009, 2, 138–139. [Google Scholar] [CrossRef] [PubMed]
- Tringe, S.G.; Hugenholtz, P. A renaissance for the pioneering 16S rRNA gene. Curr. Opin. Microbiol. 2008, 11, 442–446. [Google Scholar] [CrossRef] [PubMed]
- Trček, J.; Mahnič, A.; Rupnik, M. Diversity of the microbiota involved in wine and organic apple cider submerged vinegar production as revealed by DHPLC analysis and next-generation sequencing. Int. J. Food Microbiol. 2016, 223, 57–62. [Google Scholar] [CrossRef] [PubMed]
- David, V.; Terrat, S.; Herzine, K.; Claisse, O.; Rousseaux, S.; Tourdot-Maréchal, R.; Masneuf-Pomarede, I.; Ranjard, L.; Alexandre, H. High-throughput sequencing of amplicons for monitoring yeast biodiversity in must and during alcoholic fermentation. J. Ind. Microbiol. Biotechnol. 2014, 41, 811–821. [Google Scholar] [CrossRef] [PubMed]
- Makarova, K.; Slesarev, A.; Wolf, Y.; Sorokin, A.; Mirkin, B.; Koonin, E.; Pavlov, A.; Pavlova, N.; Karamychev, V.; Polouchine, N.; et al. Comparative genomics of the lactic acid bacteria. Proc. Natl. Acad. Sci. USA 2006, 103, 15611–15616. [Google Scholar] [CrossRef] [PubMed]
- Sun, Z.; Harris, H.M.B.; McCann, A.; Guo, C.; Argimón, S.; Zhang, W.; Yang, X.; Jeffery, I.B.; Cooney, J.C.; Kagawa, T.F.; et al. Expanding the biotechnology potential of lactobacilli through comparative genomics of 213 strains and associated genera. Nat. Commun. 2015, 6, 8322. [Google Scholar] [CrossRef] [PubMed]
- Herrera, C.M.; de Vega, C.; Canto, A.; Pozo, M.I. Yeasts in floral nectar: A quantitative survey. Ann. Bot. 2009, 103, 1415–1423. [Google Scholar] [CrossRef] [PubMed]
- Pozo, M.I.; Herrera, C.M.; Bazaga, P. Species richness of yeast communities in floral nectar of southern Spanish plants. Microb. Ecol. 2011, 61, 82–91. [Google Scholar] [CrossRef] [PubMed]
- Pusey, P.L.; Stockwell, V.O.; Mazzola, M. Epiphytic bacteria and yeasts on apple blossoms and their potential as antagonists of Erwinia amylovora. Phytopathology 2009, 99, 571–581. [Google Scholar] [CrossRef] [PubMed]
- Graça, A.; Santo, D.; Esteves, E.; Nunes, C.; Abadias, M.; Quintas, C. Evaluation of microbial quality and yeast diversity in fresh-cut apple. Food Microbiol. 2015, 51, 179–185. [Google Scholar] [CrossRef] [PubMed]
- Batzer, J.C.; Sisson, A.J.; Harrington, T.C.; Mayfield, D.A.; Gleason, M.L. Temporal patterns in appearance of sooty blotch and flyspeck fungi on apples. Microb. Ecol. 2012, 64, 928–941. [Google Scholar] [CrossRef] [PubMed]
- Ismail, S.I.; Batzer, J.C.; Harrington, T.C.; Crous, P.W.; Lavrov, D.V.; Li, H.; Gleason, M.L. Ancestral state reconstruction infers phytopathogenic origins of sooty blotch and flyspeck fungi on apple. Mycologia 2016, 108, 292–302. [Google Scholar] [CrossRef] [PubMed]
- Alonso, S.; Laca, A.; Rendueles, M.; Mayo, B.; Díaz, M. Cider apple native microbiota characterization by PCR-DGGE. J. Inst. Brew. 2015, 121, 287–289. [Google Scholar] [CrossRef]
- Morrissey, W.F.; Davenport, B.; Querol, A.; Dobson, A.D. The role of indigenous yeasts in traditional Irish cider fermentations. J. Appl. Microbiol. 2004, 97, 647–655. [Google Scholar] [CrossRef] [PubMed]
- Coton, E.; Coton, M.; Levert, D.; Casaregola, S.; Sohier, D. Yeast ecology in French cider and black olive natural fermentations. Int. J. Food. Microbiol. 2006, 108, 130–135. [Google Scholar] [CrossRef] [PubMed]
- Suarez Valles, B.; Bedrinana, R.P.; Tascon, N.F.; Simon, A.Q.; Madrera, R.R. Yeast species associated with the spontaneous fermentation of cider. Food Microbiol. 2007, 24, 25–31. [Google Scholar] [CrossRef] [PubMed]
- Pando Bedrinana, R.; Querol Simon, A.; Suarez Valles, B. Genetic and phenotypic diversity of autochthonous cider yeasts in a cellar from Asturias. Food Microbiol. 2010, 27, 503–508. [Google Scholar] [CrossRef] [PubMed]
- Shade, A.; McManus, P.S.; Handelsman, J. Unexpected Diversity during Community Succession in the Apple Flower Microbiome. mBio 2013, 4, e00602–e00612. [Google Scholar] [CrossRef] [PubMed]
- Sánchez, A.; Coton, M.; Coton, E.; Herrero, M.; García, L.A.; Díaz, M. Prevalent lactic acid bacteria in cider cellars and efficiency of Oenococcus oeni strains. Food Microbiol. 2012, 32, 32–37. [Google Scholar] [CrossRef] [PubMed]
- Claisse, O.; Lonvaud-Funel, A. Détection de bactéries lactiques produisant du 3-hydroxypropionaldéhyde (précurseur d’acroléine) à partir du glycérol par tests moléculaires. Le Lait 2001, 81, 173–181. [Google Scholar] [CrossRef]
- Puertas, A.I.; Arahal, D.R.; Ibarburu, I.; Elizaquível, P.; Aznar, R.; Dueñas, M.T. Lactobacillus sicerae sp. nov., a lactic acid bacterium isolated from Spanish natural cider. Int. J. Syst. Evol. Microbiol. 2014, 64, 2949–2955. [Google Scholar] [CrossRef] [PubMed]
- Garai, G.; Dueñas, M.T.; Irastorza, A.; Moreno-Arribas, M.V. Biogenic amine production by lactic acid bacteria isolated from cider. Lett. Appl. Microbiol. 2007, 45, 473–478. [Google Scholar] [CrossRef] [PubMed]
- Salih, A.G.; Drilleau, J.F.; Cavin, F.F.; Divies, C.; Bourgeois, C.M. A Survey of Microbiological Aspects of Cider Making. J. Inst. Brew. 1988, 94, 5–8. [Google Scholar] [CrossRef]
- El Khoury, M.; Campbell-Sills, H.; Salin, F.; Guichoux, E.; Claisse, O.; Lucas, P.M. Biogeography of Oenococcus oeni reveals distinctive but nonspecific populations in wine-producing regions. Appl. Environ. Microbiol. 2017, 83. [Google Scholar] [CrossRef] [PubMed]
- Coton, M.; Romano, A.; Spano, G.; Ziegler, K.; Vetrana, C.; Desmarais, C.; Lonvaud-Funel, A.; Lucas, P.; Coton, E. Occurrence of biogenic amine-forming lactic acid bacteria in wine and cider. Food Microbiol. 2010, 27, 1078–1085. [Google Scholar] [CrossRef] [PubMed]
- Swings, J.; De Ley, J. The biology of Zymomonas. Bacteriol. Rev. 1977, 41, 1–46. [Google Scholar] [PubMed]
- Bauduin, R.; Le Quere, J.M.; Coton, E.; Primault, J. Factors leading to the expression of ‘framboisé’ in French ciders. LWT Food Sci. Technol. 2006, 39, 966–971. [Google Scholar] [CrossRef]
- Keller, S.E.; Chirtel, S.J.; Merker, R.I.; Taylor, K.T.; Tan, H.L.; Miller, A.J. Influence of fruit variety, harvest technique, quality sorting, and storage on the native microflora of unpasteurized apple cider. J. Food Prot. 2004, 67, 2240–2247. [Google Scholar] [CrossRef] [PubMed]
- Mills, D.A.; Rawsthorne, H.; Parker, C.; Tamir, D.; Makarova, K. Genomic analysis of Oenococcus oeni PSU-1 and its relevance to winemaking. FEMS Microbiol. Rev. 2005, 29, 465–475. [Google Scholar] [CrossRef] [PubMed]
- Bridier, J.; Claisse, O.; Coton, M.; Coton, E.; Lonvaud-Funel, A. Evidence of distinct populations and specific subpopulations within the species Oenococcus oeni. Appl. Environ. Microbiol. 2010, 76, 7754–7764. [Google Scholar] [CrossRef] [PubMed]
- Campbell-Sills, H.; El Khoury, M.; Favier, M.; Romano, A.; Biasioli, F.; Spano, G.; Sherman, D.J.; Bouchez, O.; Coton, E.; Coton, M.; et al. Phylogenomic analysis of Oenococcus oeni reveals specific domestication of strains to cider and wines. Genome Biol. Evol. 2015, 7, 1506–1518. [Google Scholar] [CrossRef] [PubMed]
- Sternes, P.R.; Borneman, A.R. Consensus pan-genome assembly of the specialised wine bacterium Oenococcus oeni. BMC Genom. 2016, 17. [Google Scholar] [CrossRef]
- Pérez-Ramos, A.; Mohedano, M.L.; Puertas, A.; Lamontanara, A.; Orru, L.; Spano, G.; Capozzi, V.; Dueñas, M.T.; López, P. Draft genome sequence of Pediococcus parvulus 2.6, a probiotic β-glucan producer strain. Genome Announc. 2016, 4. [Google Scholar] [CrossRef] [PubMed]
- Puertas, A.I.; Capozzi, V.; Llamas, M.G.; López, P.; Lamontanara, A.; Orrù, L.; Russo, P.; Spano, G.; Dueñas, M.T. Draft genome sequence of Lactobacillus collinoides CUPV237, an exopolysaccharide and riboflavin producer isolated from cider. Genome Announc. 2016, 4. [Google Scholar] [CrossRef] [PubMed]
- Ottesen, A.R.; White, J.R.; Skaltsas, D.N.; Newell, M.J.; Walsh, C.S. Impact of organic and conventional management on the phyllosphere microbial ecology of an apple crop. J. Food Prot. 2009, 72, 2321–2325. [Google Scholar] [CrossRef] [PubMed]
- Yashiro, E.; Spear, R.N.; McManus, P.S. Culture-dependent and culture-independent assessment of bacteria in the apple phyllosphere. J. Appl. Microbiol. 2011, 110, 1284–1296. [Google Scholar] [CrossRef] [PubMed]
- Granado, J.; Thurig, B.; Kieffer, E.; Petrini, L.; Fliessbach, A.; Tamm, L.; Weibel, F.P.; Wyss, G.S. Culturable fungi of stored ‘golden delicious’ apple fruits: A one-season comparison study of organic and integrated production systems in Switzerland. Microb. Ecol. 2008, 56, 720–732. [Google Scholar] [CrossRef] [PubMed]
- Fleet, G.H. Yeast interactions and wine flavour. Int. J. Food Microbiol. 2003, 86, 11–22. [Google Scholar] [CrossRef]
- Azhu Valappil, Z.; Fan, X.; Zhang, H.Q.; Rouseff, R.L. Impact of thermal and nonthermal processing technologies on unfermented apple cider aroma volatiles. J. Agric. Food Chem. 2009, 57, 924–929. [Google Scholar] [CrossRef] [PubMed]
- Lilly, M.; Lambrechts, M.G.; Pretorius, I.S. Effect of increased yeast alcohol acetyltransferase activity on flavor profiles of wine and distillates. Appl. Environ. Microbiol. 2000, 66, 744–753. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Zhao, G.A.; Wang, L.P. Controlled formation of volatile components in cider making using a combination of Saccharomyces cerevisiae and Hanseniaspora valbyensis yeast species. J. Ind. Microbiol. Biotechnol. 2006, 33, 192–196. [Google Scholar] [CrossRef] [PubMed]
- Sumby, K.M.; Grbin, P.R.; Jiranek, V. Microbial modulation of aromatic esters in wine: Current knowledge and future prospects. Food Chem. 2010, 121, 1–16. [Google Scholar] [CrossRef]
- Mangas, J.J.; Cabranes, C.; Moreno, J.; Gomis, D.B. Influence of Cider-Making Technology on Cider Taste. LWT Food Sci. Technol. 1994, 27, 583–586. [Google Scholar] [CrossRef]
- Herrero, M. Influence des Fermentations Alcoolique et Malolactique sur la Composition Chimique des Cidres À Distiller en Cours D’élaboaration; Université de Caen: Caen, France, 2011. [Google Scholar]
- Rapp, A.; Mandery, H. New progess in wine and wine research. Experientia 1987, 45, 873–884. [Google Scholar]
- Rous, C.V.; Snow, R.; Kunkee, R.E. Reduction of Higher Alcohols by Fermentation with a Leucine-Auxotrophic Mutant of Wine Yeast. J. Inst. Brew. 1983, 89, 274–278. [Google Scholar] [CrossRef]
- Jepsen, O.M. The sensory and analitical evaluation of apple juice volatiles. Int. Fed. Fruit Juice Prod. 1978, 15, 349–361. [Google Scholar]
- Ye, M.; Yue, T.; Yuan, Y. Evolution of polyphenols and organic acids during the fermentation of apple cider. J. Sci. Food Agric. 2014, 94, 2951–2957. [Google Scholar] [CrossRef] [PubMed]
- Lea, A.G.H.; Arnold, G.M. The Phenolics of Cider: Bitterness and Astringency. J. Sci. Food Agric. 1978, 29, 478–483. [Google Scholar] [CrossRef]
- Symoneaux, R.; Baron, A.; Marnet, N.; Bauduin, R.; Chollet, S. Impact of apple procyanidins on sensory perception in model cider (part 1): Polymerisation degree and concentration. LWT Food Sci. Technol. 2014, 57, 22–27. [Google Scholar] [CrossRef]
- Symoneaux, R.; Chollet, S.; Bauduin, R.; Le Quéré, J.M.; Baron, A. Impact of apple procyanidins on sensory perception in model cider (part 2): Degree of polymerization and interactions with the matrix components. LWT Food Sci. Technol. 2014, 57, 28–34. [Google Scholar] [CrossRef]
- Vanbeneden, N.; Van Roey, T.; Willems, F.; Delvaux, F.; Delvaux, F.R. Release of phenolic flavour precursors during wort production: Influence of process parameters and grist composition on ferulic acid release during brewing. Food Chem. 2008, 111, 83–91. [Google Scholar] [CrossRef]
- Stratford, M.; James, S.A. 12-Non-alcoholic beverages and yeasts. In Yeasts in Food; Woodhead Publishing Series in Food Science; Technology and Nutrition; Boekhout, T., Robert, V., Eds.; Woodhead Publishing: Sawston, UK; 2003; pp. 309–345. ISBN 978-1-85573-706-8. [Google Scholar]
- Smith, B.D.; Divol, B. Brettanomyces bruxellensis, a survivalist prepared for the wine apocalypse and other beverages. Food Microbiol. 2016, 59, 161–175. [Google Scholar] [CrossRef] [PubMed]
- Buron, N.; Coton, M.; Legendre, P.; Ledauphin, J.; Kientz-Bouchart, V.; Guichard, H.; Barillier, D.; Coton, E. Implications of Lactobacillus collinoides and Brettanomyces/Dekkera anomala in phenolic off-flavour defects of ciders. Int. J. Food Microbiol. 2012, 153, 159–165. [Google Scholar] [CrossRef] [PubMed]
- Wedral, D.; Shewfelt, R.; Frank, J. The challenge of Brettanomyces in wine. LWT Food Sci. Technol. 2010, 43, 1474–1479. [Google Scholar] [CrossRef]
- Mehlomakulu, N.N.; Setati, M.E.; Divol, B. Characterization of novel killer toxins secreted by wine-related non-Saccharomyces yeasts and their action on Brettanomyces spp. Int. J. Food Microbiol. 2014, 188, 83–91. [Google Scholar] [CrossRef] [PubMed]
- Quirós, C.; Herrero, M.; García, L.A.; Díaz, M. Effects of SO2 on lactic acid bacteria physiology when used as a preservative compound in malolactic fermentation. J. Inst. Brew. 2012, 118, 89–96. [Google Scholar] [CrossRef]
- Lonvaud-Funel, A. Lactic acid bacteria in the quality improvement and depreciation of wine. Antonie Leeuwenhoek 1999, 76, 317–331. [Google Scholar] [CrossRef]
- Henick-Kling, T. Control of malolactic fermentation in wine: Energetics, flavour modification and methods of starter culture preparation. J. Appl. Bacteriol. 1995, 79, 29–37. [Google Scholar]
- Osborne, J.P.; Mira de Orduña, R.; Pilone, G.J.; Liu, S.Q. Acetaldehyde metabolism by wine lactic acid bacteria. FEMS Microbiol. Lett. 2000, 191, 51–55. [Google Scholar] [CrossRef] [PubMed]
- Ugliano, M.; Moio, L. Changes in the concentration of yeast-derived volatile compounds of red wine during malolactic fermentation with four commercial starter cultures of Oenococcus oeni. J. Agric. Food Chem. 2005, 53, 10134–10139. [Google Scholar] [CrossRef] [PubMed]
- Swiegers, J.H.; Bartowsky, E.J.; Henschke, P.A.; Pretorius, I.S. Yeast and bacterial modulation of wine aroma and flavour. Aust. J. Grape Wine Res. 2005, 11, 139–173. [Google Scholar] [CrossRef]
- Maicas, S.; Gil, J.V.; Pardo, I.; Ferrer, S. Improvement of volatile composition of wines by controlled addition of malolactic bacteria. Food Res. Int. 1999, 32, 491–496. [Google Scholar] [CrossRef]
- Bartowsky, E.J.; Henschke, P.A. The ‘buttery’ attribute of wine—diacetyl—desirability, spoilage and beyond. Int. J. Food Microbiol. 2004, 96, 235–252. [Google Scholar] [CrossRef] [PubMed]
- Sauvageot, N.; Muller, C.; Hartke, A.; Auffray, Y.; Laplace, J.-M. Characterisation of the diol dehydratase pdu operon of Lactobacillus collinoides. FEMS Microbiol. Lett. 2002, 209, 66–71. [Google Scholar] [CrossRef]
- Sauvageot, N.; Pichereau, V.; Louarme, L.; Hartke, A.; Auffray, Y.; Laplace, J.-M. Purification, characterization and subunits identification of the diol dehydratase of Lactobacillus collinoides. Eur. J. Biochem. 2002, 269, 5731–5737. [Google Scholar] [CrossRef] [PubMed]
- Garai-Ibabe, G.; Ibarburu, I.; Berregi, I.; Claisse, O.; Lonvaud-Funel, A.; Irastorza, A.; Dueñas, M.T. Glycerol metabolism and bitterness producing lactic acid bacteria in cidermaking. Int. J. Food Microbiol. 2008, 121, 253–261. [Google Scholar] [CrossRef] [PubMed]
- Ledauphin, J.; Lefrancois, A.; Marquet, N.; Beljean-Leymarie, M.; Barillier, D. Development of an accurate and sensitive gas chromatographic method for the determination of acrolein content in Calvados and cider. LWT Food Sci. Technol. 2006, 39, 1045–1052. [Google Scholar] [CrossRef]
- Carr, J.G.; Davies, P.A. The Ecology and Classification of Strains of Lactobacillus collinoides nov. spec.: A Bacterium Commonly Found in Fermenting Apple Juice. J. Appl. Bacteriol. 1972, 35, 463–471. [Google Scholar] [CrossRef] [PubMed]
- Sauvageot, N.; Gouffi, K.; Laplace, J.M.; Auffray, Y. Glycerol metabolism in Lactobacillus collinoides: Production of 3-hydroxypropionaldehyde, a precursor of acrolein. Int. J. Food Microbiol. 2000, 55, 167–170. [Google Scholar] [CrossRef]
- Coton, M.; Laplace, J.M.; Coton, E. Zymomonas mobilis subspecies identification by amplified ribosomal DNA restriction analysis. Lett. Appl. Microbiol. 2005, 40, 152–157. [Google Scholar] [CrossRef] [PubMed]
- Jarvis, B. Cider (Cyder; Hard Cider). In Encyclopedia of Food Microbiology, 2nd ed.; Batt, C.A., Tortorello, M.L., Eds.; Academic Press: Oxford, UK, 2014; pp. 437–443. ISBN 978-0-12-384733-1. [Google Scholar]
- Ladero, V.; Cruz Martin, M.; Fernandez, M.; Alvarez, M.A. Toxicological Effects of Dietary Biogenic Amines. Curr. Nutr. Food Sci. 2010, 6, 145–156. [Google Scholar] [CrossRef]
- Ladero, V.; Coton, M.; Fernández, M.; Buron, N.; Martín, M.C.; Guichard, H.; Coton, E.; Alvarez, M.A. Biogenic amines content in Spanish and French natural ciders: Application of qPCR for quantitative detection of biogenic amine-producers. Food Microbiol. 2011, 28, 554–561. [Google Scholar] [CrossRef] [PubMed]
- Izquierdo-Pulido, M.; Albalá-Hurtado, S.; Mariné-Font, A.; Vidal-Carou, M.C. Biogenic amines in Spanish beers: Differences among breweries. Z. Lebensm. Unters. Forsch. 1996, 203, 507–511. [Google Scholar] [CrossRef]
- Marcobal, A.; Martín-Alvarez, P.J.; Polo, M.C.; Muñoz, R.; Moreno-Arribas, M.V. Formation of biogenic amines throughout the industrial manufacture of red wine. J. Food Prot. 2006, 69, 397–404. [Google Scholar] [CrossRef] [PubMed]
- Vidal-Carou, M.C.; Ambatlle-Espunyes, A.; Ulla-Ulla, M.C.; Mariné-Font, A. Histamine and Tyramine in Spanish Wines: Their Formation During the Winemaking Process. Am. J. Enol. Vitic. 1990, 41, 160–167. [Google Scholar]
- Martín-Álvarez, P.J.; Marcobal, Á.; Polo, C.; Moreno-Arribas, M.V. Influence of technological practices on biogenic amine contents in red wines. Eur. Food Res. Technol. 2006, 222, 420–424. [Google Scholar] [CrossRef]
- Alvarez, M.A.; Moreno-Arribas, M.V. The problem of biogenic amines in fermented foods and the use of potential biogenic amine-degrading microorganisms as a solution. Trends Food Sci. Technol. 2014, 39, 146–155. [Google Scholar] [CrossRef]
- Garai-Ibabe, G.; Irastorza, A.; Dueñas, M.T.; Martín-Álvarez, P.J.; Moreno-Arribas, V.M. Evolution of amino acids and biogenic amines in natural ciders as a function of the year and the manufacture steps. Int. J. Food Sci. Technol. 2013, 48, 375–381. [Google Scholar] [CrossRef]
- Del Campo, G.; Lavado, I.; Dueñas, M.; Irastoza, M. Histamine production by some lactic acid bacteria isolated from ciders. Food Sci. Technol. Int. 2000, 6, 117–121. [Google Scholar]
- Callejón, S.; Sendra, R.; Ferrer, S.; Pardo, I. Identification of a novel enzymatic activity from lactic acid bacteria able to degrade biogenic amines in wine. Appl. Microbiol. Biotechnol. 2014, 98, 185–198. [Google Scholar] [CrossRef] [PubMed]
- Binder, E.M.; Tan, L.M.; Chin, L.J.; Handl, J.; Richard, J. Worldwide occurrence of mycotoxins in commodities, feeds and feed ingredients. Anim. Feed Sci. Technol. 2007, 137, 265–282. [Google Scholar] [CrossRef]
- Moake, M.M.; Padilla-Zakour, O.I.; Worobo, R.W. Comprehensive Review of Patulin Control Methods in Foods. Compr. Rev. Food Sci. Food Saf. 2005, 4, 8–21. [Google Scholar] [CrossRef]
- McKinley, E.R.; Carlton, W.W. Patulin. In Mycotoxins and Phytoalexins; Shanna, R.P., Salunkhe, D.K., Eds.; CRC Press: Boca Raton, FL, USA, 1991; pp. 191–236. [Google Scholar]
- Postupolski, J.; Rybińska, K.; Kurpińska-Jaworska, J.; Ledzion, E.; Szczesna, M.; Karłowski, K. European Union legislation related to patulin. Rocz. Panstw. Zakl. Hig. 2003, 54, 355–361. [Google Scholar] [PubMed]
- Harris, K.L.; Bobe, G.; Bourquin, L.D. Patulin surveillance in apple cider and juice marketed in Michigan. J. Food Prot. 2009, 72, 1255–1261. [Google Scholar] [CrossRef] [PubMed]
- Tannous, J.; Atoui, A.; El Khoury, A.; Francis, Z.; Oswald, I.P.; Puel, O.; Lteif, R. A study on the physicochemical parameters for Penicillium expansum growth and patulin production: Effect of temperature, pH, and water activity. Food Sci. Nutr. 2016, 4, 611–622. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Cruz, M.L.; Mansilla, M.L.; Tadeo, J.L. Mycotoxins in fruits and their processed products: Analysis, occurrence and health implications. J. Adv. Res. 2010, 1, 113–122. [Google Scholar] [CrossRef]
- Pollution du Fruit et Patuline−IFPC—Cidre, Pomme, Pommier. Available online: http://www.ifpc.eu/bibliographie/recolte-qualite-des-fruits/qualite-du-fruit.html (accessed on 10 March 2017).
- Mihajlovic, B.; Dixon, B.; Couture, H.; Farber, J. Qualitative Microbiological Risk Assessment of Unpasteurized Fruit Juice and Cider. Int. Food Risk Anal. J. 2013, 3, 1–19. [Google Scholar] [CrossRef]
- Merwin, I.A.; Valois, S.; Padilla-Zakour, O.I. Cider Apples and Cider-Making Techniques in Europe and North America. In Horticultural Reviews; Janick, J., Ed.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2007; Volume 34, pp. 365–415. ISBN 978-0-470-38014-7. [Google Scholar]
- Garcia, L.; Henderson, J.; Fabri, M.; Oke, M. Potential sources of microbial contamination in unpasteurized apple cider. J. Food Prot. 2006, 69, 137–144. [Google Scholar] [CrossRef] [PubMed]
- Kniel, K.E.; Sumner, S.S.; Lindsay, D.S.; Hackney, C.R.; Pierson, M.D.; Zajac, A.M.; Golden, D.A.; Fayer, R. Effect of organic acids and hydrogen peroxide on Cryptosporidium parvum viability in fruit juices. J. Food Prot. 2003, 66, 1650–1657. [Google Scholar] [CrossRef] [PubMed]
- Bartowsky, E.J. Oenococcus oeni and malolactic fermentation-moving into the molecular arena. Aust. J. Grape Wine Res. 2005, 11, 174–187. [Google Scholar] [CrossRef]
- Magalhães, F.; Krogerus, K.; Vidgren, V.; Sandell, M.; Gibson, B. Improved cider fermentation performance and quality with newly generated Saccharomyces cerevisiae × Saccharomyces eubayanus hybrids. J. Ind. Microbiol. Biotechnol. 2017. [Google Scholar] [CrossRef] [PubMed]
- Betteridge, A.; Grbin, P.; Jiranek, V. Improving Oenococcus oeni to overcome challenges of wine malolactic fermentation. Trends Biotechnol. 2015, 33, 547–553. [Google Scholar] [CrossRef] [PubMed]
- Schümann, C.; Michlmayr, H.; Eder, R.; del Hierro, A.M.; Kulbe, K.D.; Mathiesen, G.; Nguyen, T.-H. Heterologous expression of Oenococcus oeni malolactic enzyme in Lactobacillus plantarum for improved malolactic fermentation. AMB Express 2012, 2, 19. [Google Scholar] [CrossRef] [PubMed]
- Du Toit, M.; Engelbrecht, L.; Lerm, E.; Krieger-Weber, S. Lactobacillus: The next generation of malolactic fermentation starter cultures—An overview. Food Bioprocess Technol. 2011, 4, 876–906. [Google Scholar] [CrossRef]
- Suárez Valles, B.; Pando Bedriñana, R.; Lastra Queipo, A.; Mangas Alonso, J.J. Screening of cider yeasts for sparkling cider production (Champenoise method). Food Microbiol. 2008, 25, 690–697. [Google Scholar] [CrossRef] [PubMed]
- De Arruda Moura Pietrowski, G.; dos Santos, C.M.E.; Sauer, E.; Wosiacki, G.; Nogueira, A. Influence of fermentation with Hanseniaspora sp. yeast on the volatile profile of fermented apple. J. Agric. Food Chem. 2012, 60, 9815–9821. [Google Scholar] [CrossRef] [PubMed]
- Kot, W.; Neve, H.; Heller, K.J.; Vogensen, F.K. Bacteriophages of Leuconostoc, Oenococcus, and Weissella. Front. Microbiol. 2014, 5, 186. [Google Scholar] [CrossRef] [PubMed]
- Costantini, A.; Doria, F.; Saiz, J.-C.; Garcia-Moruno, E. Phage-host interactions analysis of newly characterized Oenococcus oeni bacteriophages: Implications for malolactic fermentation in wine. Int. J. Food Microbiol. 2017, 246, 12–19. [Google Scholar] [CrossRef] [PubMed]
- Endersen, L.; O’Mahony, J.; Hill, C.; Ross, R.P.; McAuliffe, O.; Coffey, A. Phage therapy in the food industry. Annu. Rev. Food Sci. Technol. 2014, 5, 327–349. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, M.; Viñas, I.; Colàs, P.; Anguera, M.; Usall, J.; Abadias, M. Effectiveness of a bacteriophage in reducing Listeria monocytogenes on fresh-cut fruits and fruit juices. Food Microbiol. 2014, 38, 137–142. [Google Scholar] [CrossRef] [PubMed]
- Aneja, K.R.; Dhiman, R.; Aggarwal, N.K.; Aneja, A. Emerging preservation techniques for controlling spoilage and pathogenic microorganisms in fruit juices. Int. J. Microbiol. 2014, 2014, e758942. [Google Scholar] [CrossRef] [PubMed]
- Ye, M.; Yue, T.; Yuan, Y. Effects of sequential mixed cultures of Wickerhamomyces anomalus and Saccharomyces cerevisiae on apple cider fermentation. FEMS Yeast Res. 2014, 14, 873–882. [Google Scholar] [CrossRef] [PubMed]
- Priya, P.; Munishamanna, K.B. Microbial fermentation of blended tomato juice by yeast and lactic acid bacteria for nutritional improvement. Environ. Ecol. 2013, 31, 181–186. [Google Scholar]
- Peng, B.; Li, F.; Cui, L.; Guo, Y. Effects of fermentation temperature on key aroma compounds and sensory properties of apple wine. J. Food Sci. 2015, 80, S2937–S2943. [Google Scholar] [CrossRef] [PubMed]
- Herrero, M.; García, L.A.; Díaz, M. Volatile compounds in cider: Inoculation time and fermentation temperature effects. J. Inst. Brew. 2006, 112, 210–214. [Google Scholar] [CrossRef]
- Satora, P.; Tarko, T.; Duda-Chodak, A.; Sroka, P.; Tuszyński, T.; Czepielik, M. Influence of prefermentative treatments and fermentation on the antioxidant and volatile profiles of apple wines. J. Agric. Food Chem. 2009, 57, 11209–11217. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Lovitt, R.W. Strategies for enhanced malolactic fermentation in wine and cider maturation. J. Chem. Technol. Biotechnol. 2006, 81, 1130–1140. [Google Scholar] [CrossRef]
- Devi Avaiyarasi, N.; David Ravindran, A.; Venkatesh, P.; Arul, V. In vitro selection, characterization and cytotoxic effect of bacteriocin of Lactobacillus sakei GM3 isolated from goat milk. Food Control 2016, 69, 124–133. [Google Scholar] [CrossRef]
- Wen, L.S.; Philip, K.; Ajam, N. Purification, characterization and mode of action of plantaricin K25 produced by Lactobacillus plantarum. Food Control 2016, 60, 430–439. [Google Scholar] [CrossRef]
- Hu, Y.; Liu, X.; Shan, C.; Xia, X.; Wang, Y.; Dong, M.; Zhou, J. Novel bacteriocin produced by Lactobacillus alimentarius FM-MM4 from a traditional Chinese fermented meat Nanx Wudl: Purification, identification and antimicrobial characteristics. Food Control 2017, 77, 290–297. [Google Scholar] [CrossRef]
- Laplace, J.M.; Sauvageot, N.; Hartke, A.; Auffray, Y. Characterization of Lactobacillus collinoides response to heat, acid and ethanol treatments. Appl. Microbiol. Biotechnol. 1999, 51, 659–663. [Google Scholar] [CrossRef]
- Gálvez, A.; Maqueda, M.; Valdivia, E.; Quesada, A.; Montoya, E. Characterization and partial purification of a broad spectrum antibiotic AS-48 produced by Streptococcus faecalis. Can. J. Microbiol. 1986, 32, 765–771. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Viedma, P.; Abriouel, H.; Omar, N.B.; Valdivia, E.; López, R.L.; Gálvez, A. Inactivation of exopolysaccharide and 3-hydroxypropionaldehyde-producing lactic acid bacteria in apple juice and apple cider by enterocin AS-48. Food Chem. Toxicol. Int. J. Public Br. Ind. Biol. Res. Assoc. 2008, 46, 1143–1151. [Google Scholar] [CrossRef] [PubMed]
- Dueñas, M.; Irastorza, A.; Fernández, K.; Bilbao, A. Heterofermentative Lactobacilli causing ropiness in Basque country ciders. J. Food Prot. 1995, 58, 76–80. [Google Scholar] [CrossRef]
- Ibarburu, I.; Puertas, A.I.; Berregi, I.; Rodríguez-Carvajal, M.A.; Prieto, A.; Dueñas, M.T. Production and partial characterization of exopolysaccharides produced by two Lactobacillus suebicus strains isolated from cider. Int. J. Food Microbiol. 2015, 214, 54–62. [Google Scholar] [CrossRef] [PubMed]
- Korakli, M.; Gänzle, M.G.; Vogel, R.F. Metabolism by bifidobacteria and lactic acid bacteria of polysaccharides from wheat and rye, and exopolysaccharides produced by Lactobacillus sanfranciscensis. J. Appl. Microbiol. 2002, 92, 958–965. [Google Scholar] [CrossRef] [PubMed]
- Dueñas-Chasco, M.T.; Rodríguez-Carvajal, M.A.; Tejero Mateo, P.; Franco-Rodríguez, G.; Espartero, J.L.; Irastorza-Iribas, A.; Gil-Serrano, A.M. Structural analysis of the exopolysaccharide produced by Pediococcus damnosus 2.6. Carbohydr. Res. 1997, 303, 453–458. [Google Scholar] [CrossRef]
- Dueñas-Chasco, M.T.; Rodríguez-Carvajal, M.A.; Tejero-Mateo, P.; Espartero, J.L.; Irastorza-Iribas, A.; Gil-Serrano, A.M. Structural analysis of the exopolysaccharides produced by Lactobacillus spp. G-77. Carbohydr. Res. 1998, 307, 125–133. [Google Scholar] [CrossRef]
- Ibarburu, I.; Soria-Díaz, M.E.; Rodríguez-Carvajal, M.A.; Velasco, S.E.; Tejero-Mateo, P.; Gil-Serrano, A.M.; Irastorza, A.; Dueñas, M.T. Growth and exopolysaccharide (EPS) production by Oenococcus oeni I4 and structural characterization of their EPSs. J. Appl. Microbiol. 2007, 103, 477–486. [Google Scholar] [CrossRef] [PubMed]
- Larpin, S.; Sauvageot, N.; Pichereau, V.; Laplace, J.-M.; Auffray, Y. Biosynthesis of exopolysaccharide by a Bacillus licheniformis strain isolated from ropy cider. Int. J. Food Microbiol. 2002, 77, 1–9. [Google Scholar] [CrossRef]
- Grande, M.J.; Lucas, R.; Abriouel, H.; Valdivia, E.; Ben Omar, N.; Maqueda, M.; Martínez-Cañamero, M.; Gálvez, A. Inhibition of Bacillus licheniformis LMG 19409 from ropy cider by enterocin AS-48. J. Appl. Microbiol. 2006, 101, 422–428. [Google Scholar] [CrossRef] [PubMed]
- Hassan, Y.I.; Zhou, T.; Bullerman, L.B. Sourdough lactic acid bacteria as antifungal and mycotoxin-controlling agents. Rev. Agaroquim. Tecnol. Aliment. 2016, 22, 79–90. [Google Scholar] [CrossRef]
- Ahmad Rather, I.; Seo, B.J.; Rejish Kumar, V.J.; Choi, U.-H.; Choi, K.-H.; Lim, J.H.; Park, Y.-H. Isolation and characterization of a proteinaceous antifungal compound from Lactobacillus plantarum YML007 and its application as a food preservative. Lett. Appl. Microbiol. 2013, 57, 69–76. [Google Scholar] [CrossRef] [PubMed]
- Zoghi, A.; Khosravi-Darani, K.; Sohrabvandi, S.; Attar, H.; Alavi, S.A. Effect of probiotics on patulin removal from synbiotic apple juice. J. Sci. Food Agric. 2016. [Google Scholar] [CrossRef] [PubMed]
- Juodeikiene, G.; Basinskiene, L.; Bartkiene, E.; Matusevicius, P. Mycotoxin Decontamination Aspects in Food, Feed and Renewables Using Fermentation Processes. In Structure and Function of Food Engineering; InTech: Rijeka, Croatia, 2012. [Google Scholar] [CrossRef]
- Prado, F.C.; Parada, J.L.; Pandey, A.; Soccol, C.R. Trends in non-dairy probiotic beverages. Food Res. Int. 2008, 41, 111–123. [Google Scholar] [CrossRef]
- Kandylis, P.; Pissaridi, K.; Bekatorou, A.; Kanellaki, M.; Koutinas, A.A. Dairy and non-dairy probiotic beverages. Curr. Opin. Food Sci. 2016, 7, 58–63. [Google Scholar] [CrossRef]
- Marsh, A.J.; Hill, C.; Ross, R.P.; Cotter, P.D. Fermented beverages with health-promoting potential: Past and future perspectives. Trends Food Sci. Technol. 2014, 38, 113–124. [Google Scholar] [CrossRef] [Green Version]
- Konowalchuk, J.; Speirs, J.I. Antiviral effect of apple beverages. Appl. Environ. Microbiol. 1978, 36, 798–801. [Google Scholar] [PubMed]
- Gawkowski, D.; Chikindas, M.L. Non-dairy probiotic beverages: The next step into human health. Benef. Microbes 2013, 4, 127–142. [Google Scholar] [CrossRef] [PubMed]
- De Souza Neves Ellendersen, L.; Granato, D.; Bigetti Guergoletto, K.; Wosiacki, G. Development and sensory profile of a probiotic beverage from apple fermented with Lactobacillus casei. Eng. Life Sci. 2012, 12, 475–485. [Google Scholar] [CrossRef]
- Sornplang, P.; Piyadeatsoontorn, S. Probiotic isolates from unconventional sources: A review. J. Anim. Sci. Technol. 2016, 58, 26. [Google Scholar] [CrossRef] [PubMed]
- Upadrasta, A.; O’Sullivan, L.; O’Sullivan, O.; Sexton, N.; Lawlor, P.G.; Hill, C.; Fitzgerald, G.F.; Stanton, C.; Ross, R.P. The effect of dietary supplementation with spent cider yeast on the Swine distal gut microbiome. PLoS ONE 2013, 8, e75714. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernández de Palencia, P.; Werning, M.L.; Sierra-Filardi, E.; Dueñas, M.T.; Irastorza, A.; Corbí, A.L.; López, P. Probiotic properties of the 2-substituted (1,3)-β-d-glucan-producing bacterium Pediococcus parvulus 2.6. Appl. Environ. Microbiol. 2009, 75, 4887–4891. [Google Scholar] [CrossRef] [PubMed]
- Waters, D.M.; Mauch, A.; Coffey, A.; Arendt, E.K.; Zannini, E. Lactic acid bacteria as a cell factory for the delivery of functional biomolecules and ingredients in cereal-based beverages: A review. Crit. Rev. Food Sci. Nutr. 2015, 55, 503–520. [Google Scholar] [CrossRef] [PubMed]
- García-Ruiz, A.; González de Llano, D.; Esteban-Fernández, A.; Requena, T.; Bartolomé, B.; Moreno-Arribas, M.V. Assessment of probiotic properties in lactic acid bacteria isolated from wine. Food Microbiol. 2014, 44, 220–225. [Google Scholar] [CrossRef] [PubMed]
- Foligné, B.; Dewulf, J.; Breton, J.; Claisse, O.; Lonvaud-Funel, A.; Pot, B. Probiotic properties of non-conventional lactic acid bacteria: Immunomodulation by Oenococcus oeni. Int. J. Food Microbiol. 2010, 140, 136–145. [Google Scholar] [CrossRef] [PubMed]
- Dimopoulou, M.; Bardeau, T.; Ramonet, P.-Y.; Miot-Certier, C.; Claisse, O.; Doco, T.; Petrel, M.; Lucas, P.; Dols-Lafargue, M. Exopolysaccharides produced by Oenococcus oeni: From genomic and phenotypic analysis to technological valorization. Food Microbiol. 2016, 53, 10–17. [Google Scholar] [CrossRef] [PubMed]
- Zannini, E.; Waters, D.M.; Coffey, A.; Arendt, E.K. Production, properties, and industrial food application of lactic acid bacteria-derived exopolysaccharides. Appl. Microbiol. Biotechnol. 2016, 100, 1121–1135. [Google Scholar] [CrossRef] [PubMed]
- Sabokbar, N.; Khodaiyan, F.; Moosavi-Nasab, M. Optimization of processing conditions to improve antioxidant activities of apple juice and whey based novel beverage fermented by kefir grains. J. Food Sci. Technol. 2015, 52, 3422–3432. [Google Scholar] [CrossRef] [PubMed]


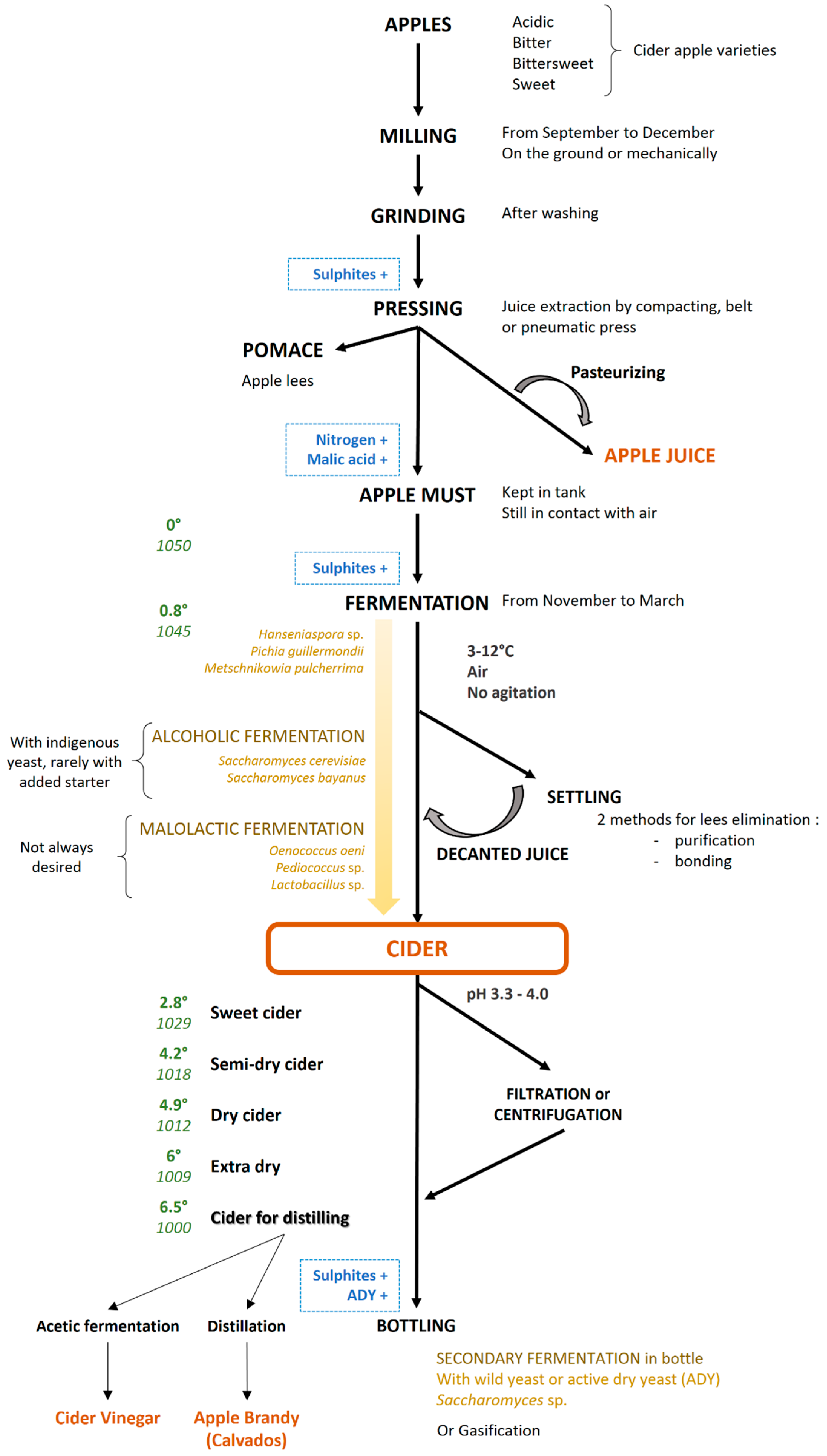{kind=link}
| Family | Origin | Genus/Species | References |
|---|---|---|---|
| Lactobacillaceae | Apple flower Fresh-cut apple Cider | Lactobacillus brevis | [24,33,34,35,36] |
| Lactobacillus (para)collinoides | |||
| Lactobacillus casei | |||
| Lactobacillus diolivorans | |||
| Lactobacillus hilgardii | |||
| Lactobacillus sicerae | |||
| Lactobacillus suebicus | |||
| Pediococcus ethanolidurans | |||
| Pediococcus parvulus | |||
| Leuconostocaceae | Cider | Oenococcus oeni | [37,38] |
| Leuconostoc mesenteroides | |||
| Acetobacteraceae | Apple flower Apple cider vinegar | Acetobacter sp. | [17,32] |
| Komagataeibacter sp. | |||
| Gluconobacter sp. | |||
| Sporolactobacilliaceae | Cider | Sporolactobacillus sp. | [39] |
| Sphingomonadaceae | Cider | Zymomonas mobilis | [40,41] |
| Enterobacteriaceae | Apple surface Apple flower | Coliforms Enterobacteriaceae a | [24,27,32] |
| Biogenic Amine | Producer | References |
|---|---|---|
| Histamine | Lactobacillus paracollinoides | [35,86,94] |
| Lactobacillus hilgardii | ||
| Lactobacillus diolivorans | ||
| Lactobacillus collinoides | ||
| Oenococcus oeni | ||
| Putrescine | Lactobacillus collinoides | [39,89] |
| Oenococcus oeni | ||
| Lactobacillus brevis | ||
| Lactobacillus mali | ||
| Leuconostoc mesenteroides | ||
| Pediococcus parvulus | ||
| Lactobacillus paracollinoides | ||
| Tyramine | Sporolactobacillus sp. | [35,86,93] |
| Lactobacillus brevis | ||
| Lactobacillus diolivorans | ||
| Oenococcus oeni | ||
| Pediococcus parvulus |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cousin, F.J.; Le Guellec, R.; Schlusselhuber, M.; Dalmasso, M.; Laplace, J.-M.; Cretenet, M. Microorganisms in Fermented Apple Beverages: Current Knowledge and Future Directions. Microorganisms 2017, 5, 39. https://doi.org/10.3390/microorganisms5030039
Cousin FJ, Le Guellec R, Schlusselhuber M, Dalmasso M, Laplace J-M, Cretenet M. Microorganisms in Fermented Apple Beverages: Current Knowledge and Future Directions. Microorganisms. 2017; 5(3):39. https://doi.org/10.3390/microorganisms5030039
Chicago/Turabian StyleCousin, Fabien J., Rozenn Le Guellec, Margot Schlusselhuber, Marion Dalmasso, Jean-Marie Laplace, and Marina Cretenet. 2017. "Microorganisms in Fermented Apple Beverages: Current Knowledge and Future Directions" Microorganisms 5, no. 3: 39. https://doi.org/10.3390/microorganisms5030039
APA StyleCousin, F. J., Le Guellec, R., Schlusselhuber, M., Dalmasso, M., Laplace, J. -M., & Cretenet, M. (2017). Microorganisms in Fermented Apple Beverages: Current Knowledge and Future Directions. Microorganisms, 5(3), 39. https://doi.org/10.3390/microorganisms5030039