Inteins: Localized Distribution, Gene Regulation, and Protein Engineering for Biological Applications


Abstract
:1. Introduction
2. Hint Domain Superfamily and Different Forms
3. Intein-Hint Domain
4. Hedgehog-Hint Domain
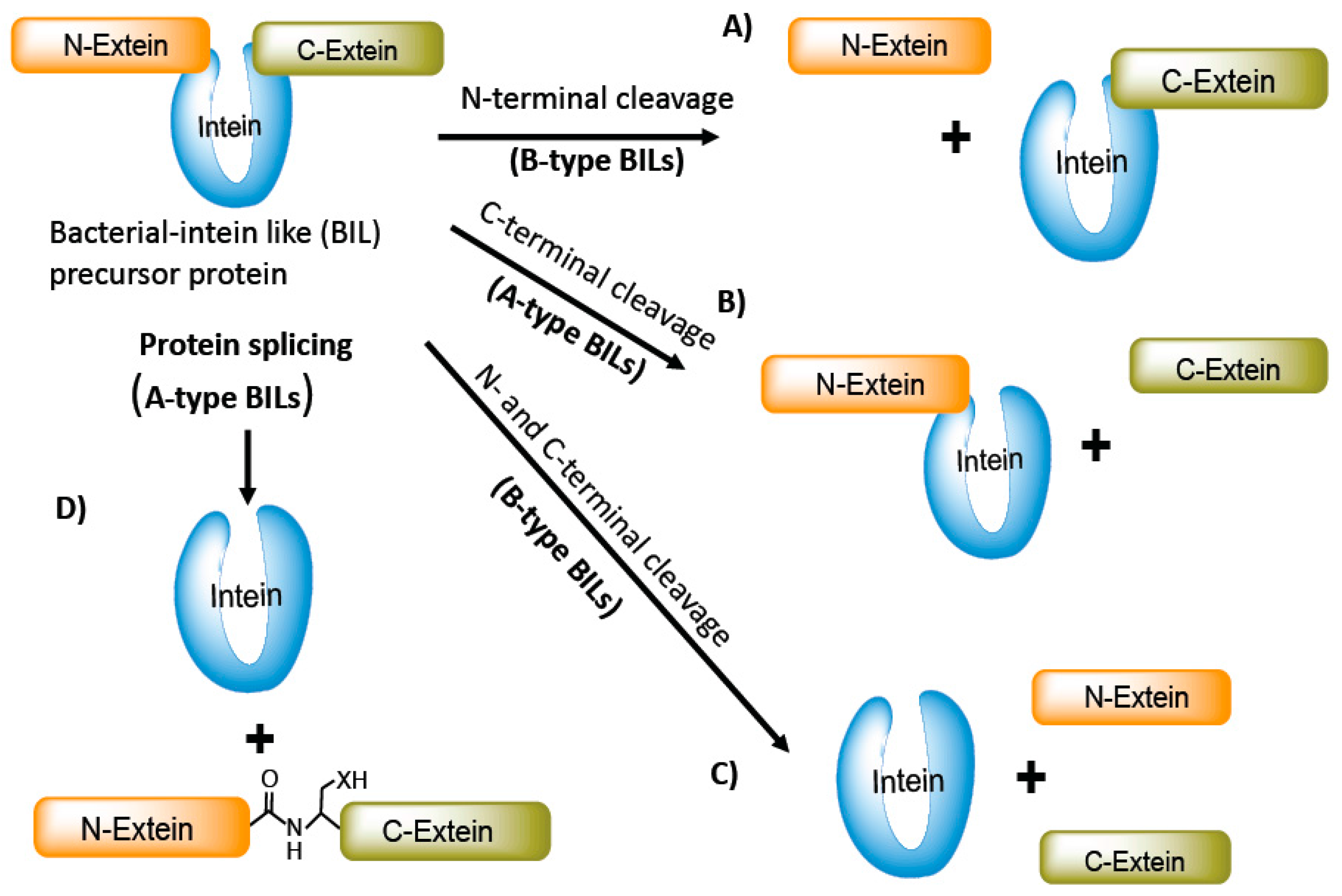
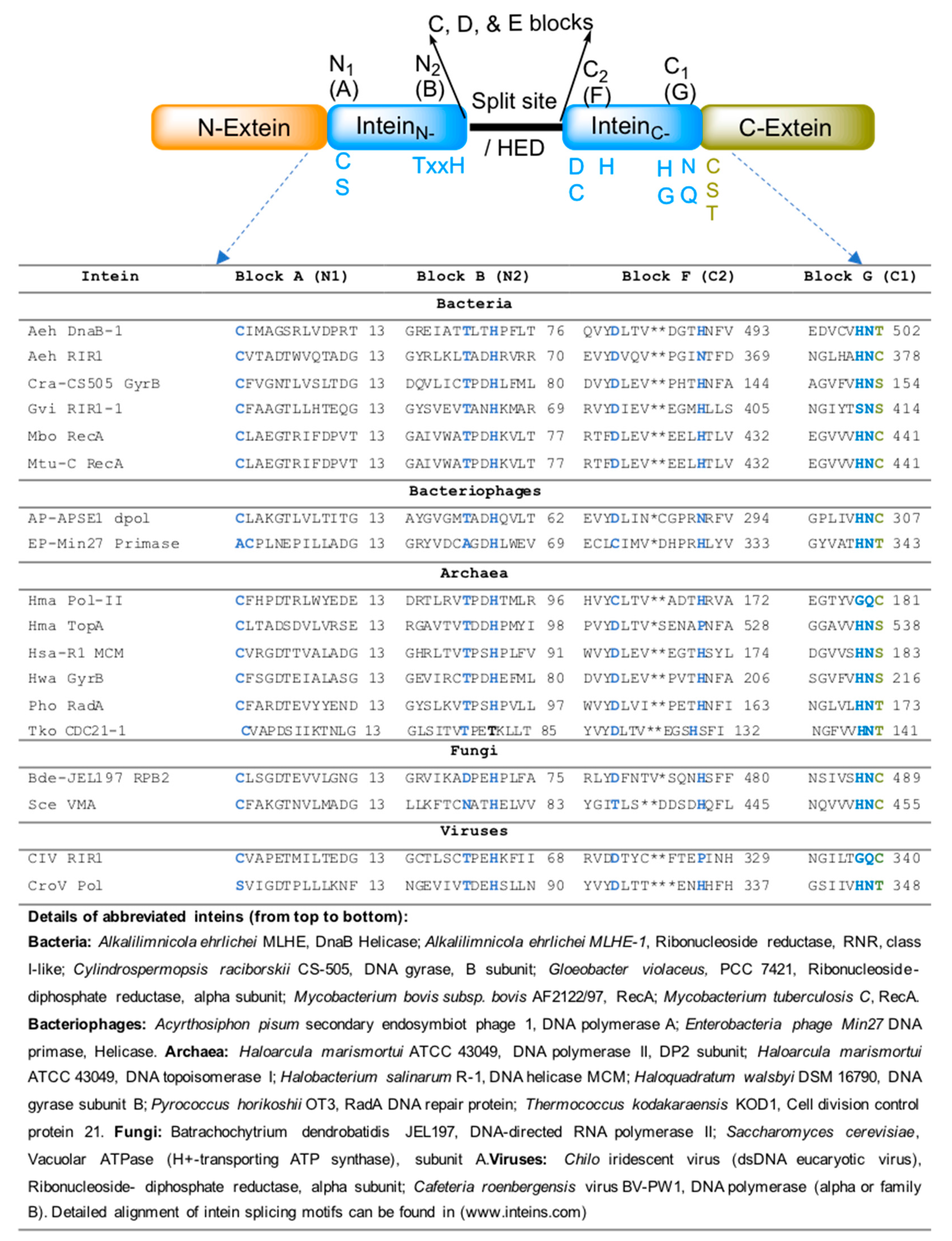
5. Bacterial-Intein-Like Hint Domain
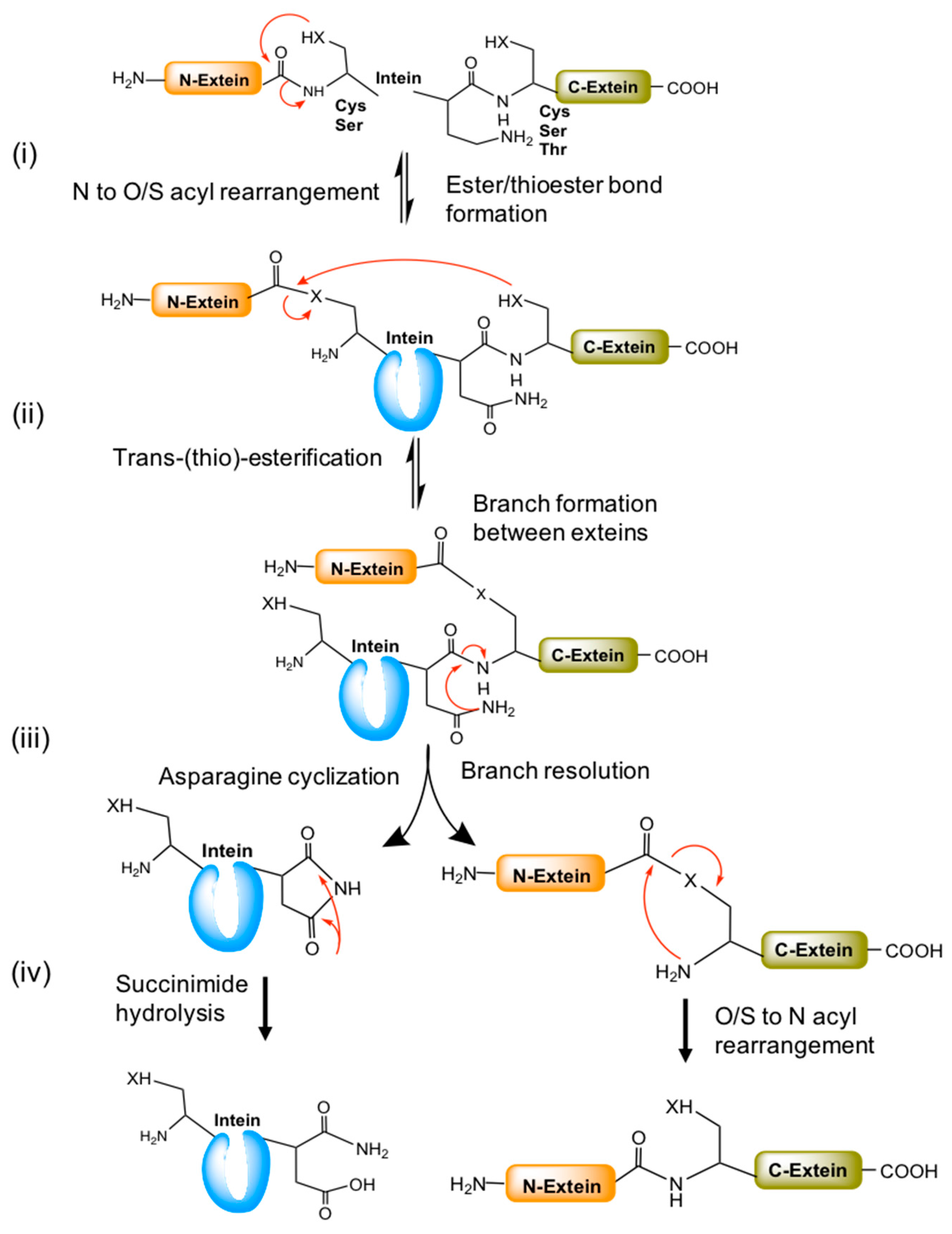
6. Mechanism of Intein-Mediated Protein Splicing
- Ester/thioester bond formation by N to O/S acyl rearrangement,
- Trans-(thio)-esterification between exteins,
- Asparagine cyclization, and
- Spontaneous hydrolysis of the amino-succinamide residue and O/S to N acyl rearrangement to form a peptide bond between exteins (Figure 4).
7. Localized Distribution of Inteins and Post-Translational Regulation
- Why do inteins localize in DNA replication, recombination, and repair proteins, and in RNA transcription machinery?
- Is there any significance of intein association with particular conserved regions (like P-loop of NTPases or catalytic sites) of essential proteins?
- Do they have a regulatory role in DNA replication machinery under stress conditions? or
- Is it just a selective pressure that retains intein association with particular regions of certain functional proteins, as argued earlier [17]?
8. Applications of Intein Splicing in Protein Engineering and Biological Applications
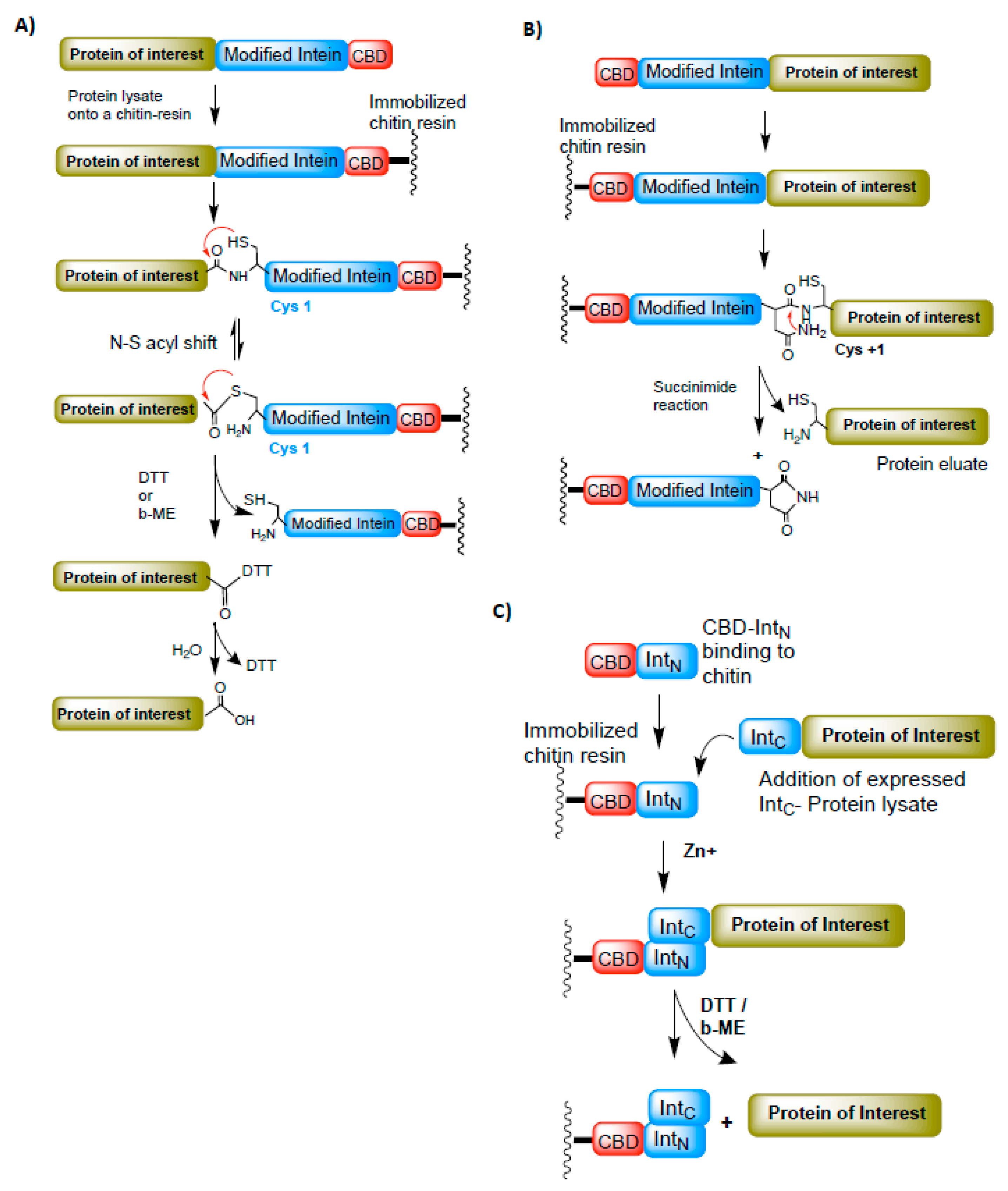
9. Inteins in Protein Purification
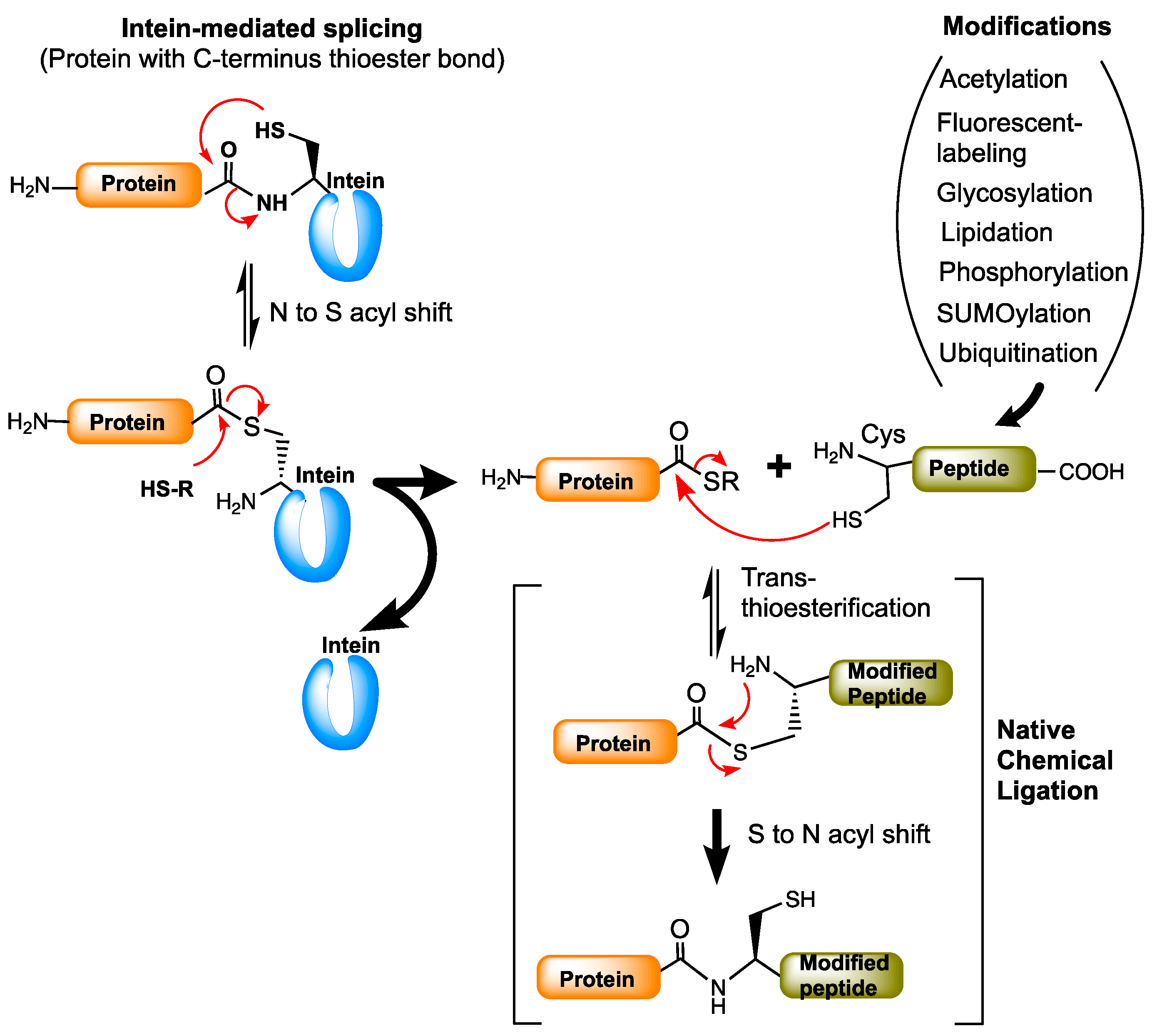
10. Protein Modifications Using Splicing Chemistry
11. Inteins as Microbial Drug Target
12. Conclusions
Acknowledgments
Conflicts of Interest
Abbreviations
| HED | Homing-endonuclease domain |
| 3Rs | replication, repair and recombination |
| BIL | bacterial intein-like |
| SRR | sterol recognition region |
| DTT | dithiothreitol |
| NTPase | nucleotide tri-phosphotases |
| β-ME | beta-Mercaptoethanol |
| NCL | native chemical ligation |
| EPL | expressed protein ligation |
| PTS | protein trans-synthesis |
| CDB | chitin-binding domain |
References
- Hirata, R.; Ohsumk, Y.; Nakano, A.; Kawasaki, H.; Suzuki, K.; Anraku, Y. Molecular structure of a gene, VMA1, encoding the catalytic subunit of H(+)-translocating adenosine triphosphatase from vacuolar membranes of Saccharomyces cerevisiae. J. Biol. Chem. 1990, 265, 6726–6733. [Google Scholar] [PubMed]
- Kane, P.; Yamashiro, C.; Wolczyk, D.; Neff, N.; Goebl, M.; Stevens, T. Protein splicing converts the yeast TFP1 gene product to the 69-kd subunit of the vacuolar H(+)-adenosine triphosphatase. Science 1990, 250, 651–657. [Google Scholar] [CrossRef] [PubMed]
- Perler, F.B. Inbase: The intein database. Nucleic Acids Res. 2002, 30, 383–384. [Google Scholar] [CrossRef] [PubMed]
- Zettler, J.; Schutz, V.; Mootz, H.D. The naturally split Npu DnaE intein exhibits an extraordinarily high rate in the protein trans-splicing reaction. FEBS Lett. 2009, 583, 909–914. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.Q.; Yang, J. Split dnaE genes encoding multiple novel inteins in Trichodesmium erythraeum. J. Biol. Chem. 2003, 278, 26315–26318. [Google Scholar] [CrossRef] [PubMed]
- Sorci, M.; Dassa, B.; Liu, H.; Anand, G.; Dutta, A.K.; Pietrokovski, S.; Belfort, M.; Belfort, G. Oriented covalent immobilization of antibodies for measurement of intermolecular binding forces between zipper-like contact surfaces of split inteins. Anal. Chem. 2013, 85, 6080–6088. [Google Scholar] [CrossRef] [PubMed]
- Hall, T.M.; Porter, J.A.; Young, K.E.; Koonin, E.V.; Beachy, P.A.; Leahy, D.J. Crystal structure of a hedgehog autoprocessing domain: Homology between hedgehog and self-splicing proteins. Cell 1997, 91, 85–97. [Google Scholar] [CrossRef]
- Chevalier, B.S.; Stoddard, B.L. Homing endonucleases: Structural and functional insight into the catalysts of intron/intein mobility. Nucleic Acids Res. 2001, 29, 3757–3774. [Google Scholar] [CrossRef] [PubMed]
- Caspi, J.; Amitai, G.; Belenkiy, O.; Pietrokovski, S. Distribution of split dnaE inteins in cyanobacteria. Mol. Microbiol. 2003, 50, 1569–1577. [Google Scholar] [CrossRef] [PubMed]
- Iwai, H.; Zuger, S.; Jin, J.; Tam, P.H. Highly efficient protein trans-splicing by a naturally split dnaE intein from Nostoc punctiforme. FEBS Lett. 2006, 580, 1853–1858. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Hu, Z.; Liu, X.Q. Protein trans-splicing by a split intein encoded in a split dnaE gene of Synechocystis sp. PCC6803. Proc. Natl. Acad. Sci. USA 1998, 95, 9226–9231. [Google Scholar] [CrossRef] [PubMed]
- Burglin, T.R. The hedgehog protein family. Genome Biol. 2008, 9, 241. [Google Scholar] [CrossRef] [PubMed]
- Amitai, G.; Belenkiy, O.; Dassa, B.; Shainskaya, A.; Pietrokovski, S. Distribution and function of new bacterial intein-like protein domains. Mol. Microbiol. 2003, 47, 61–73. [Google Scholar] [CrossRef] [PubMed]
- Dassa, B.; Haviv, H.; Amitai, G.; Pietrokovski, S. Protein splicing and auto-cleavage of bacterial intein-like domains lacking a C’-flanking nucleophilic residue. J. Biol. Chem. 2004, 279, 32001–32007. [Google Scholar] [CrossRef] [PubMed]
- Tori, K.; Cheriyan, M.; Pedamallu, C.S.; Contreras, M.A.; Perler, F.B. The Thermococcus kodakaraensis Tko CDC21-1 intein activates its N-terminal splice junction in the absence of a conserved histidine by a compensatory mechanism. Biochemistry 2012, 51, 2496–2505. [Google Scholar] [CrossRef] [PubMed]
- Frutos, S.; Goger, M.; Giovani, B.; Cowburn, D.; Muir, T.W. Branched intermediate formation stimulates peptide bond cleavage in protein splicing. Nat. Chem. Biol. 2010, 6, 527–533. [Google Scholar] [CrossRef] [PubMed]
- Novikova, O.; Jayachandran, P.; Kelley, D.S.; Morton, Z.; Merwin, S.; Topilina, N.I.; Belfort, M. Intein clustering suggests functional importance in different domains of life. Mol. Biol. Evol. 2016, 33, 783–799. [Google Scholar] [CrossRef] [PubMed]
- Novikova, O.; Topilina, N.; Belfort, M. Enigmatic distribution, evolution, and function of inteins. J. Biol. Chem. 2014, 289, 14490–14497. [Google Scholar] [CrossRef] [PubMed]
- Goodwin, T.J.; Butler, M.I.; Poulter, R.T. Multiple, non-allelic, intein-coding sequences in eukaryotic RNA polymerase genes. BMC Biol. 2006, 4, 38. [Google Scholar] [CrossRef] [PubMed]
- Swithers, K.S.; Senejani, A.G.; Fournier, G.P.; Gogarten, J.P. Conservation of intron and intein insertion sites: Implications for life histories of parasitic genetic elements. BMC Evol. Biol. 2009, 9, 303. [Google Scholar] [CrossRef] [PubMed]
- Pietrokovski, S. Identification of a virus intein and a possible variation in the protein-splicing reaction. Curr. Biol. 1998, 8, R634–R635. [Google Scholar] [CrossRef]
- Davis, E.O.; Thangaraj, H.S.; Brooks, P.C.; Colston, M.J. Evidence of selection for protein introns in the recAs of pathogenic Mycobacteria. EMBO J. 1994, 13, 699–703. [Google Scholar] [PubMed]
- Mills, K.V.; Lew, B.M.; Jiang, S.; Paulus, H. Protein splicing in trans by purified N- and C-terminal fragments of the Mycobacterium tuberculosis recA intein. Proc. Natl. Acad. Sci. USA 1998, 95, 3543–3548. [Google Scholar] [CrossRef] [PubMed]
- Frischkorn, K.; Springer, B.; Bottger, E.C.; Davis, E.O.; Colston, M.J.; Sander, P. In vivo splicing and functional characterization of Mycobacterium leprae recA. J. Bacteriol. 2000, 182, 3590–3592. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Li, L.; Du, Z.; Liu, J.; Reitter, J.N.; Mills, K.V.; Linhardt, R.J.; Wang, C. Intramolecular disulfide bond between catalytic cysteines in an intein precursor. J. Am. Chem. Soc. 2012, 134, 2500–2503. [Google Scholar] [CrossRef] [PubMed]
- Topilina, N.I.; Green, C.M.; Jayachandran, P.; Kelley, D.S.; Stanger, M.J.; Piazza, C.L.; Nayak, S.; Belfort, M. Sufb intein of mycobacterium tuberculosis as a sensor for oxidative and nitrosative stresses. Proc. Natl. Acad. Sci. USA 2015, 112, 10348–10353. [Google Scholar] [CrossRef] [PubMed]
- Topilina, N.I.; Novikova, O.; Stanger, M.; Banavali, N.K.; Belfort, M. Post-translational environmental switch of RadA activity by extein-intein interactions in protein splicing. Nucleic Acids Res. 2015, 43, 6631–6648. [Google Scholar] [CrossRef] [PubMed]
- Lennon, C.W.; Stanger, M.; Belfort, M. Protein splicing of a recombinase intein induced by ssDNA and DNA damage. Genes Dev. 2016, 30, 2663–2668. [Google Scholar] [CrossRef] [PubMed]
- Chong, S.; Mersha, F.B.; Comb, D.G.; Scott, M.E.; Landry, D.; Vence, L.M.; Perler, F.B.; Benner, J.; Kucera, R.B.; Hirvonen, C.A.; et al. Single-column purification of free recombinant proteins using a self-cleavable affinity tag derived from a protein splicing element. Gene 1997, 192, 271–281. [Google Scholar] [CrossRef]
- Chong, S.; Montello, G.E.; Zhang, A.; Cantor, E.J.; Liao, W.; Xu, M.Q.; Benner, J. Utilizing the c-terminal cleavage activity of a protein splicing element to purify recombinant proteins in a single chromatographic step. Nucleic Acids Res. 1998, 26, 5109–5115. [Google Scholar] [CrossRef] [PubMed]
- Southworth, M.W.; Amaya, K.; Evans, T.C.; Xu, M.Q.; Perler, F.B. Purification of proteins fused to either the amino or carboxy terminus of the Mycobacterium xenopi gyrase a intein. Biotechniques 1999, 27, 110–114. [Google Scholar] [PubMed]
- Banki, M.R.; Feng, L.; Wood, D.W. Simple bioseparations using self-cleaving elastin-like polypeptide tags. Nat. Methods 2005, 2, 659–661. [Google Scholar] [CrossRef] [PubMed]
- Banki, M.R.; Gerngross, T.U.; Wood, D.W. Novel and economical purification of recombinant proteins: Intein-mediated protein purification using in vivo polyhydroxybutyrate (PHB) matrix association. Protein Sci. 2005, 14, 1387–1395. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Li, N.; Wang, Y.; Wu, Y.; Mu, T.; Zheng, Y.; Huang, L.; Fang, X. Ubiquitin-intein and SUMO2-intein fusion systems for enhanced protein production and purification. Protein Expr. Purif. 2012, 82, 174–178. [Google Scholar] [CrossRef] [PubMed]
- Wood, D.W.; Derbyshire, V.; Wu, W.; Chartrain, M.; Belfort, M.; Belfort, G. Optimized single-step affinity purification with a self-cleaving intein applied to human acidic fibroblast growth factor. Biotechnol. Prog. 2000, 16, 1055–1063. [Google Scholar] [CrossRef] [PubMed]
- Xie, Y.G.; Luan, C.; Zhang, H.W.; Han, F.F.; Feng, J.; Choi, Y.J.; Groleau, D.; Wang, Y.Z. Effects of thioredoxin: SUMO and intein on soluble fusion expression of an antimicrobial peptide OG2 in Escherichia coli. Protein Pept. Lett. 2013, 20, 54–60. [Google Scholar] [CrossRef] [PubMed]
- Aranko, A.S.; Oeemig, J.S.; Zhou, D.; Kajander, T.; Wlodawer, A.; Iwai, H. Structure-based engineering and comparison of novel split inteins for protein ligation. Mol. Biosyst. 2014, 10, 1023–1034. [Google Scholar] [CrossRef] [PubMed]
- Lu, W.; Sun, Z.; Tang, Y.; Chen, J.; Tang, F.; Zhang, J.; Liu, J.N. Split intein facilitated tag affinity purification for recombinant proteins with controllable tag removal by inducible auto-cleavage. J. Chromatogr. A 2011, 1218, 2553–2560. [Google Scholar] [CrossRef] [PubMed]
- Shah, N.H.; Dann, G.P.; Vila-Perello, M.; Liu, Z.; Muir, T.W. Ultrafast protein splicing is common among cyanobacterial split inteins: Implications for protein engineering. J. Am. Chem. Soc. 2012, 134, 11338–11341. [Google Scholar] [CrossRef] [PubMed]
- Stevens, A.J.; Brown, Z.Z.; Shah, N.H.; Sekar, G.; Cowburn, D.; Muir, T.W. Design of a split intein with exceptional protein splicing activity. J. Am. Chem. Soc. 2016, 138, 2162–2165. [Google Scholar] [CrossRef] [PubMed]
- Stevens, A.J.; Sekar, G.; Shah, N.H.; Mostafavi, A.Z.; Cowburn, D.; Muir, T.W. A promiscuous split intein with expanded protein engineering applications. Proc. Natl. Acad. Sci. USA 2017, 114, 8538–8543. [Google Scholar] [CrossRef] [PubMed]
- Wong, S.; Mosabbir, A.A.; Truong, K. An engineered split intein for photoactivated protein trans-splicing. PLoS ONE 2015, 10, e0135965. [Google Scholar] [CrossRef] [PubMed]
- Ramirez, M.; Valdes, N.; Guan, D.; Chen, Z. Engineering split intein dnaE from nostoc punctiforme for rapid protein purification. Protein Eng. Des. Sel. 2013, 26, 215–223. [Google Scholar] [CrossRef] [PubMed]
- Guan, D.; Chen, Z. Affinity purification of proteins in tag-free form: Split intein-mediated ultrarapid purification (sirp). Methods Mol. Biol. 2017, 1495, 1–12. [Google Scholar] [PubMed]
- Dawson, P.E.; Muir, T.W.; Clark-Lewis, I.; Kent, S.B. Synthesis of proteins by native chemical ligation. Science 1994, 266, 776–779. [Google Scholar] [CrossRef] [PubMed]
- Evans, T.C., Jr.; Benner, J.; Xu, M.Q. Semisynthesis of cytotoxic proteins using a modified protein splicing element. Protein Sci. 1998, 7, 2256–2264. [Google Scholar] [CrossRef] [PubMed]
- Muir, T.W.; Sondhi, D.; Cole, P.A. Expressed protein ligation: A general method for protein engineering. Proc. Natl. Acad. Sci. USA 1998, 95, 6705–6710. [Google Scholar] [CrossRef] [PubMed]
- Paulus, H. Protein splicing and related forms of protein autoprocessing. Ann. Rev. Biochem. 2000, 69, 447–496. [Google Scholar] [CrossRef] [PubMed]
- Vila-Perello, M.; Muir, T.W. Biological applications of protein splicing. Cell 2010, 143, 191–200. [Google Scholar] [CrossRef] [PubMed]
- Chacko, B.M.; Qin, B.Y.; Tiwari, A.; Shi, G.; Lam, S.; Hayward, L.J.; De Caestecker, M.; Lin, K. Structural basis of heteromeric Smad protein assembly in TGF-beta signaling. Mol. Cell 2004, 15, 813–823. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.W.; Hu, M.; Chai, J.; Seoane, J.; Huse, M.; Li, C.; Rigotti, D.J.; Kyin, S.; Muir, T.W.; Fairman, R.; et al. Crystal structure of a phosphorylated SMAD2. Recognition of phosphoserine by the MH2 domain and insights on SMAD function in TGF-beta signaling. Mol. Cell 2001, 8, 1277–1289. [Google Scholar] [CrossRef]
- Buskas, T.; Ingale, S.; Boons, G.J. Glycopeptides as versatile tools for glycobiology. Glycobiology 2006, 16, 113R–136R. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Ai, Y.; Wang, J.; Haracska, L.; Zhuang, Z. Chemically ubiquitylated PCNA as a probe for eukaryotic translesion DNA synthesis. Nat. Chem. Biol. 2010, 6, 270–272. [Google Scholar] [CrossRef] [PubMed]
- Chatterjee, C.; Muir, T.W. Chemical approaches for studying histone modifications. J. Biol. Chem. 2010, 285, 11045–11050. [Google Scholar] [CrossRef] [PubMed]
- Maag, D.; Fekete, C.A.; Gryczynski, Z.; Lorsch, J.R. A conformational change in the eukaryotic translation preinitiation complex and release of EIF1 signal recognition of the start codon. Mol. Cell 2005, 17, 265–275. [Google Scholar] [CrossRef] [PubMed]
- Wood, D.W.; Camarero, J.A. Intein applications: From protein purification and labeling to metabolic control methods. J. Biol. Chem. 2014, 289, 14512–14519. [Google Scholar] [CrossRef] [PubMed]
- Evans, T.C., Jr.; Martin, D.; Kolly, R.; Panne, D.; Sun, L.; Ghosh, I.; Chen, L.; Benner, J.; Liu, X.Q.; Xu, M.Q. Protein trans-splicing and cyclization by a naturally split intein from the dnaE gene of Synechocystis species PCC6803. J. Biol. Chem. 2000, 275, 9091–9094. [Google Scholar] [CrossRef] [PubMed]
- Camarero, J.A.; Fushman, D.; Sato, S.; Giriat, I.; Cowburn, D.; Raleigh, D.P.; Muir, T.W. Rescuing a destabilized protein fold through backbone cyclization. J. Mol. Biol. 2001, 308, 1045–1062. [Google Scholar] [CrossRef] [PubMed]
- Camarero, J.A.; Muir, T.W. Biosynthesis of a head-to-tail cyclized protein with improved biological activity. J. Am. Chem. Soc. 1999, 121, 5597–5598. [Google Scholar] [CrossRef]
- Iwai, H.; Lingel, A.; Pluckthun, A. Cyclic green fluorescent protein produced in vivo using an artificially split PI-Pfui intein from Pyrococcus furiosus. J. Biol. Chem. 2001, 276, 16548–16554. [Google Scholar] [CrossRef] [PubMed]
- Iwai, H.; Pluckthun, A. Circular beta-lactamase: Stability enhancement by cyclizing the backbone. FEBS Lett. 1999, 459, 166–172. [Google Scholar] [CrossRef]
- Aboye, T.L.; Camarero, J.A. Biological synthesis of circular polypeptides. J. Biol. Chem. 2012, 287, 27026–27032. [Google Scholar] [CrossRef] [PubMed]
- Topilina, N.I.; Mills, K.V. Recent advances in in vivo applications of intein-mediated protein splicing. Mob. DNA 2014, 5. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Zheng, Y.; Callahan, B.; Belfort, M.; Liu, Y. Cisplatin inhibits protein splicing, suggesting inteins as therapeutic targets in Mycobacteria. J. Biol. Chem. 2011, 286, 1277–1282. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Proteins | Function | Category | Organism |
|---|---|---|---|
| RecA/RadA | DNA-strand exchange | DNA repair | Eubacteria/Archaea |
| DnaB | Replicative DNA helicase | DNA replication | Eubacteria |
| Rad54/SWI-SNF2 | dsDNA dependent ATPase | SF2 helicases & Chromatin remodeler | Eubacteria |
| UvrD/Rep/PcrA | ATP dependent DNA helicase (SF1 family) | DNA mismatch repair | Eubacteria |
| DnaE, Catalytic α-subunit of DNA pol III | DNA polymerase | DNA Replication | Eubacteria |
| Bacterial DNA polymerase I (PolA) | DNA synthesis | DNA Replication | Eubacteria |
| Bacterial DNA polymerase II (PolB) | DNA synthesis | DNA Replication | Eubacteria |
| DNA polymerase III τ and γ | DNA synthesis | DNA Replication | Eubacteria |
| RuvB | APT dependent DNA helicase | Holliday junction branch migration | Eubacteria |
| Ribonucleoisde diphosphate reductase | Ribonucleotide diphosphate reductase | DNA synthesis | Eubacteria; Archaea |
| DNA gyrase subunit A & B | Stabilizing the DNA (DNA replication, resection) | Topoisomerase | Eubacteria |
| DnaG | DNA primase | DNA replication | Eubacteria |
| RecG | ATP dependent DNA helicase | DNA replication | Eubacteria |
| Replication factor-C small unit (RFC) | DNA clamp loader | DNA Replication | Archaea |
| DNA polymerase II large unit (PolC/DP2) | DNA synthesis | DNA Replication | Archaea |
| DNA polymerase II small unit (PolB) | DNA synthesis | DNA Replication | Archaea |
| Mini-chromosome maintenance protein (MCM) | Replicative DNA helicase | DNA replication | Archaea |
© 2018 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pavankumar, T.L. Inteins: Localized Distribution, Gene Regulation, and Protein Engineering for Biological Applications. Microorganisms 2018, 6, 19. https://doi.org/10.3390/microorganisms6010019
Pavankumar TL. Inteins: Localized Distribution, Gene Regulation, and Protein Engineering for Biological Applications. Microorganisms. 2018; 6(1):19. https://doi.org/10.3390/microorganisms6010019
Chicago/Turabian StylePavankumar, Theetha L. 2018. "Inteins: Localized Distribution, Gene Regulation, and Protein Engineering for Biological Applications" Microorganisms 6, no. 1: 19. https://doi.org/10.3390/microorganisms6010019
APA StylePavankumar, T. L. (2018). Inteins: Localized Distribution, Gene Regulation, and Protein Engineering for Biological Applications. Microorganisms, 6(1), 19. https://doi.org/10.3390/microorganisms6010019