Description of New and Amended Clades of the Genus Photobacterium



Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains and Culture Conditions
2.2. Phenotypic Characterization
2.3. DNA Extraction and PCR Amplification
2.4. Phylogenetic Data Analysis
3. Results and Discussion
3.1. Phenotypic Characterization
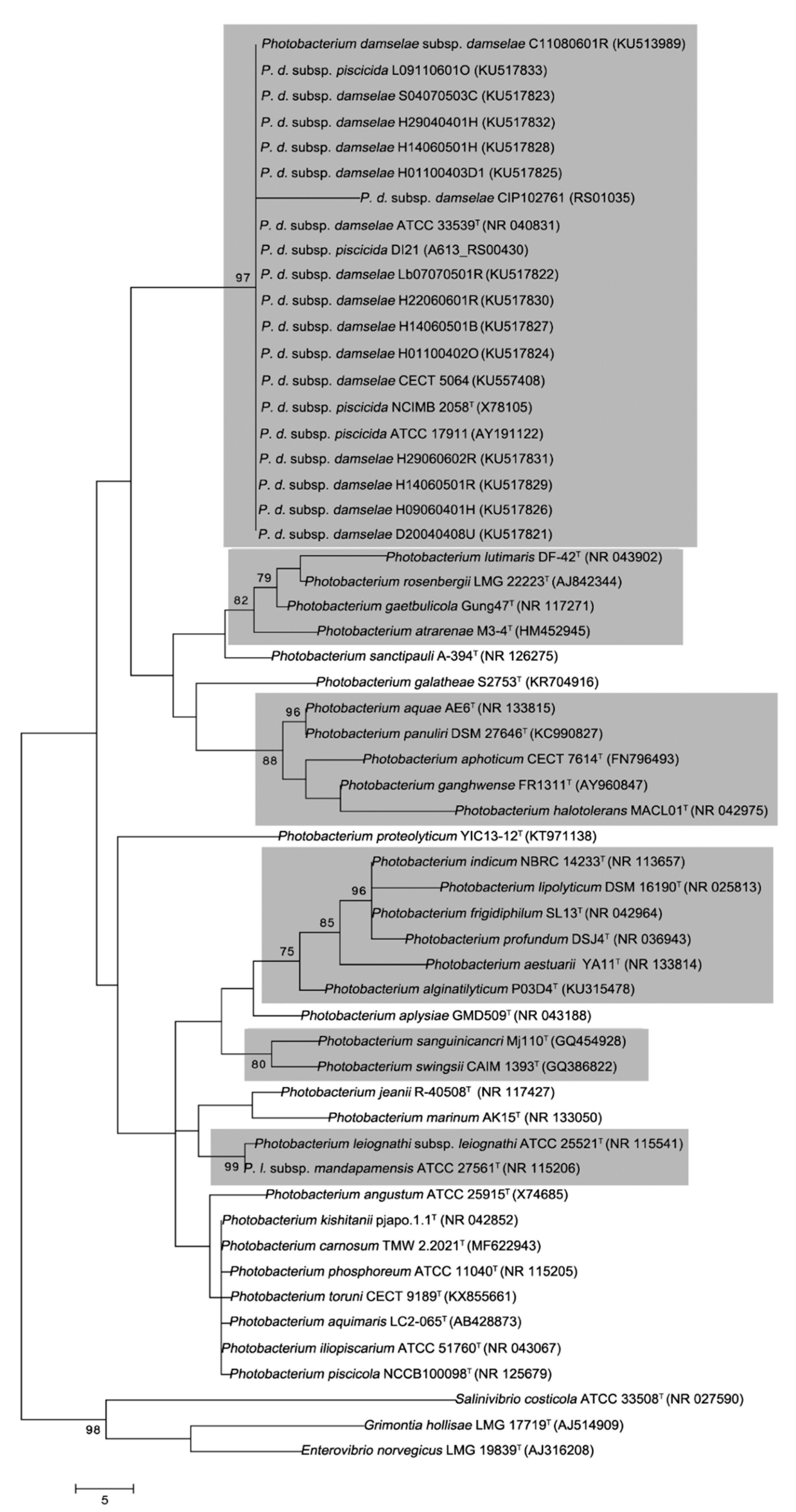
3.2. Phylogenetic Studies of the Photobacterium Genus
3.3. Intra- and Interspecies Nucleotide Sequences Variation
4. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Farmer, J.J.; Janda, J.M. Family I. Vibrionaceae. In Bergey’s Manual of Systematic Bacteriology, 2nd ed.; The Proteobacteria Part B the Gammaproteobacteria; Brenner, D.L., Krieg, N.R., Staley, J.T., Eds.; Springer: New York, NY, USA, 2005; Volume 2, pp. 491–494. [Google Scholar]
- Urbanczyk, H.; Ast, J.C.; Higgins, M.J.; Carson, J.; Dunlap, P.V. Reclassification of Vibrio fischeri, Vibrio logei, Vibrio salmonicida and Vibrio wodanis as Aliivibrio fischeri gen. nov., comb. nov., Aliivibrio logei comb. nov., Aliivibrio salmonicida comb. nov. and Aliivibrio wodanis comb. nov. Int. J. Syst. Evol. Microbiol. 2007, 57, 2823–2829. [Google Scholar] [CrossRef] [PubMed]
- LPSN–List of Prokaryotic Names with Standing in Nomenclature. Genus Photobacterium. Available online: http://www.bacterio.net/p/photobacterium.html (accessed on 5 February 2018).
- Labella, A.M.; Arahal, D.R.; Castro, D.; Lemos, M.L.; Borrego, J.J. Revisiting the genus Photobacterium: Taxonomy, ecology and pathogenesis. Int. Microbiol. 2017, 20, 1–10. [Google Scholar] [PubMed]
- Ast, J.C.; Dunlap, P.V. Phylogenetic analysis of the lux operon distinguishes two evolutionarily distinct clades of Photobacterium leiognathi. Arch. Microbiol. 2004, 181, 352–361. [Google Scholar] [CrossRef] [PubMed]
- Rivas, A.J.; Lemos, M.L.; Osorio, C.R. Photobacterium damselae subsp. damselae, a bacterium pathogenic for marine animals and humans. Front. Microbiol. 2013, 4, 283. [Google Scholar] [CrossRef] [PubMed]
- Skerman, V.B.D.; McGowan, V.; Sneath, P.H.A. Approved lists of bacterial names. Int. J. Syst. Bacteriol. 1980, 30, 225–420. [Google Scholar] [CrossRef]
- Smith, S.K.; Sutton, D.C.; Fuerst, J.A.; Reichelt, J.L. Evaluation of the genus Listonella and reassignment of Listonella damsela (Love et al.) MacDonell and Colwell to the genus Photobacterium as Photobacterium damsela comb. nov. with an emended description. Int. J. Syst. Bacteriol. 1991, 41, 529–534. [Google Scholar] [CrossRef] [PubMed]
- Gauthier, G.; Lafay, B.; Ruimy, R.; Breittmayer, V.; Nicolas, J.L.; Gauthier, M.; Christen, R. Small-subunit rRNA sequences and whole DNA relatedness concur for the reassignment of Pasteurella piscicida (Snieszko et al.) Janssen and Surgalla to the genus Photobacterium as Photobacterium damsela subsp. piscicida comb. nov. Int. J. Syst. Bacteriol. 1995, 45, 139–144. [Google Scholar] [CrossRef] [PubMed]
- Kimura, B.; Hokimoto, S.; Takahashi, H.; Fujii, T. Photobacterium histaminum Okuzumi et al. 1994 is a later subjective synonym of Photobacterium damselae subsp. damselae (Love et al. 1981) Smith et al. 1991. Int. J. Syst. Evol. Microbiol. 2000, 50, 1339–1342. [Google Scholar]
- Stackebrandt, E.; Frederiksen, W.; Garrity, G.M.; Grimont, P.A.D.; Kampfer, P.; Maiden, M.C.J.; Nesme, X.; Rossello-Mora, R.; Swings, J.; Truper, H.G.; et al. Report of the ad hoc committee for the re-evaluation of the species definition in bacteriology. Int. J. Syst. Evol. Microbiol. 2002, 52, 1043–1047. [Google Scholar] [PubMed]
- Maiden, M.C.J. Multilocus sequence typing of bacteria. Annu. Rev. Microbiol. 2006, 60, 561–588. [Google Scholar] [CrossRef] [PubMed]
- Ast, J.C.; Urbanczyk, H.; Dunlap, P.V. Multi-gene analysis reveals previously unrecognized phylogenetic diversity in Aliivibrio. Syst. Appl. Microbiol. 2009, 32, 379–386. [Google Scholar] [CrossRef] [PubMed]
- Zeigler, D.R. Gene sequences useful for predicting relatedness of whole genomes in bacteria. Int. J. Syst. Evol. Microbiol. 2003, 53, 1893–1900. [Google Scholar] [CrossRef] [PubMed]
- Park, Y.D.; Baik, K.S.; Seong, C.N.; Bae, K.S.; Kim, S.; Chun, J. Photobacterium ganghwense sp. nov., a halophilic bacterium isolated from sea water. Int. J. Syst. Evol. Microbiol. 2006, 56, 745–749. [Google Scholar] [CrossRef] [PubMed]
- Rivas, R.; García-Fraile, P.; Mateos, P.F.; Martínez-Molina, E.; Velásquez, E. Photobacterium halotolerans sp. nov., isolated from Lake Martel in Spain. Int. J. Syst. Evol. Microbiol. 2006, 56, 1067–1071. [Google Scholar] [CrossRef] [PubMed]
- Jung, S.Y.; Jung, Y.T.; Oh, T.K.; Yoon, J.H. Photobacterium lutimaris sp. nov., isolated from a tidal flat sediment in Korea. Int. J. Syst. Evol. Microbiol. 2007, 57, 332–336. [Google Scholar] [CrossRef] [PubMed]
- Yoshizawa, S.; Wada, M.; Kita-Tsukamoto, K.; Yokota, A.; Kogure, K. Photobacterium aquimaris sp. nov., a luminous marine bacterium isolated from seawater. Int. J. Syst. Evol. Microbiol. 2009, 59, 1438–1442. [Google Scholar] [CrossRef] [PubMed]
- Urbanczyk, H.; Ogura, Y.; Hendry, T.A.; Gould, A.L.; Kiwaki, N.; Atkinson, J.T.; Hayashi, T.; Dunlap, P.V. Genome sequence of Photobacterium mandapamensis strain svers.1.1, the bioluminescent symbiont of the cardinal fish Siphamia versicolor. J. Bacteriol. 2011, 193, 3144–3145. [Google Scholar] [CrossRef] [PubMed]
- Sawabe, T.; Ogura, Y.; Matsumura, Y.; Feng, G.; Rohul Amin, A.K.M.; Mino, S.; Nakagawa, S.; Sawabe, T.; Kumar, R.; Fukui, Y.; et al. Updating the Vibrio clades defined by multilocus sequence phylogeny: Proposal of eight new clades and the description of Vibrio tritonius sp. nov. Front. Microbiol. 2013, 4, 414. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.-O.; Kim, K.-K.; Park, S.; Kang, S.-J.; Lee, J.-H.; Lee, S.-J.; Oh, T.-K.; Yoon, J.-H. Photobacterium gaetbulicola sp. nov., a lipolytic bacterium isolated from a tidal flat sediment. Int. J. Syst. Evol. Microbiol. 2010, 60, 2587–2591. [Google Scholar] [CrossRef] [PubMed]
- Chimetto, L.A.; Cleenwerck, I.; Thompson, C.C.; Brocchi, M.; Willems, A.; De Vos, P.; Thompson, F.L. Photobacterium jeanii sp. nov., isolated from corals and zoanthids. Int. J. Syst. Evol. Microbiol. 2010, 60, 2843–2848. [Google Scholar] [CrossRef] [PubMed]
- Lucena, T.; Rovira, M.A.; Pascual, J.; Garay, E.; Macian, M.C.; Arahal, D.R.; Pujalte, M.J. Photobacterium aphoticum sp. nov., isolated from coastal water. Int. J. Syst. Evol. Microbiol. 2011, 61, 1579–1584. [Google Scholar] [CrossRef] [PubMed]
- Kim, B.-C.; Poo, H.; Kim, M.N.; Lee, K.H.; Lee, J.; Rhee, M.-S.; Shin, K.-S. Photobacterium atrarenae sp. nov. a novel bacterium isolated from sea sand. Curr. Microbiol. 2011, 63, 433–438. [Google Scholar] [CrossRef] [PubMed]
- Gomez-Gil, B.; Roque, A.; Rotllant, G.; Peinado, L.; Romalde, J.L.; Doce, A.; Cabanillas-Beltran, H.; Chimetto, L.; Thompson, F.L. Photobacterium swingsii sp. nov. isolated from marine organisms. Int. J. Syst. Evol. Microbiol. 2011, 61, 315–319. [Google Scholar] [CrossRef] [PubMed]
- Srinivas, T.N.R.; Vijaya Bhaskar, Y.; Bhumika, V.; Anil Kumar, P. Photobacterium marinum sp. nov., a marine bacterium isolated from a sediment sample from Palk Bay, India. Syst. Appl. Microbiol. 2013, 36, 160–165. [Google Scholar] [CrossRef] [PubMed]
- Lo, N.; Jin, H.M.; Jeon, C.O. Photobacterium aestuarii sp. nov., a marine bacterium isolated from a tidal flat. Int. J. Syst. Evol. Microbiol. 2014, 64, 625–630. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Liu, L.-Z.; Song, L.; Zhou, Y.-G.; Qi, F.-J.; Liu, Z.-P. Photobacterium aquae sp. nov., isolated from a recirculating mariculture system. Int. J. Syst. Evol. Microbiol. 2014, 64, 475–480. [Google Scholar] [CrossRef] [PubMed]
- Deep, K.; Poddar, A.; Das, S.K. Photobacterium panuliri sp. nov., an alkalitolerant marine bacterium isolated from eggs of spiny lobster, Panulirus penicillatus from Andaman Sea. Curr. Microbiol. 2014, 69, 660–668. [Google Scholar] [CrossRef] [PubMed]
- Figge, M.J.; Cleenwerck, I.; Van Uijenc, A.; De Vosb, P.; Huys, G.; Robertson, L. Photobacterium piscicola sp. nov., isolated from marine fish and spoiled packed cod. Syst. Appl. Microbiol. 2014, 37, 329–335. [Google Scholar] [CrossRef] [PubMed]
- Moreira, A.P.B.; Duytschaever, G.; Chimetto Tonon, L.A.; Froes, A.M.; De Oliveira, M.L.; Amado-Filho, G.M.; Francini-Filho, R.B.; De Vos, P.; Swings, J.; Thompson, C.C.; et al. Photobacterium sanctipauli sp. nov. isolated from bleached Madracis decactis (Scleractinia) in the St Peter & St Paul Archipelago, Mid-Atlantic Ridge, Brazil. PeerJ 2014, 2, e427. [Google Scholar] [PubMed]
- Machado, H.; Giubergia, S.; Mateiu, R.V.; Gram, L. Photobacterium galatheae sp. nov., a bioactive bacterium isolated from a mussel in the Solomon Sea. Int. J. Syst. Evol. Microbiol. 2015, 65, 4503–4507. [Google Scholar] [CrossRef] [PubMed]
- Gomez-Gil, B.; Roque, A.; Rotllant, G.; Romalde, J.L.; Doce, A.; Eggermont, M.; Defoirdt, T. Photobacterium sanguinicancri sp. nov. isolated from marine animals. Antonie Van Leeuwenhoek 2016, 109, 817–825. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zhou, M.; Wang, F.; Wang, E.T.; Du, Z.; Wu, C.; Zhang, Z.; Liu, W.; Xie, Z. Photobacterium proteolyticum sp. nov., a protease-producing bacterium isolated from ocean sediments of Laizhou Bay. Int. J. Syst. Evol. Microbiol. 2017, 67, 1835–1840. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Wang, Y.; Yang, X.; Sun, H.; Li, B.; Zhang, X.-H. Photobacterium alginatilyticum sp. nov., a marine bacterium isolated from bottom seawater. Int. J. Syst. Evol. Microbiol. 2017, 67, 1912–1917. [Google Scholar] [CrossRef] [PubMed]
- Labella, A.M.; Arahal, D.R.; Lucena, T.; Manchado, M.; Castro, D.; Borrego, J.J. Photobacterium toruni sp. nov., a bacterium isolated from diseased farmed fish. Int. J. Syst. Evol. Microbiol. 2017, 67, 4518–4525. [Google Scholar] [CrossRef] [PubMed]
- Hilgarth, M.; Fuertes, S.; Ehrmann, M.; Vogel, R.F. Photobacterium carnosum sp. nov., isolated from spoiled modified atmosphere packaged poultry meat. Syst. Appl. Microbiol. 2018, 41, 44–50. [Google Scholar] [CrossRef] [PubMed]
- Labella, A.; Vida, M.; Alonso, M.C.; Infante, C.; Cardenas, S.; Lopez-Romalde, S.; Manchado, M.; Borrego, J.J. First isolation of Photobacterium damselae ssp. damselae from cultured redbanded seabream, Pagrus auriga Valenciennes, in Spain. J. Fish Dis. 2006, 29, 175–179. [Google Scholar] [CrossRef] [PubMed]
- Labella, A.; Manchado, M.; Alonso, M.C.; Castro, D.; Romalde, J.L.; Borrego, J.J. Molecular intraspecific characterization of Photobacterium damselae ssp. damselae strains affecting cultured marine fish. J. Appl. Microbiol. 2010, 108, 2122–2132. [Google Scholar] [PubMed]
- Labella, A.; Berbel, C.; Manchado, M.; Castro, D.; Borrego, J.J. Photobacterium damselae subsp. damselae, an emerging pathogen affecting new cultured marine fish species in Southern Spain. In Recent Advances in Fish Farms; Aral, F., Doggu, Z., Eds.; InTech: Rijeka, Croatia, 2011; pp. 135–152. [Google Scholar]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [PubMed]
- Villesen, P. FaBox: An online toolbox for fasta sequences. Mol. Ecol. Notes 2007, 7, 965–968. [Google Scholar] [CrossRef]
- Martin, D.; Rybicki, E. RDP: Detection of recombination amongst aligned sequences. Bioinformatics 2000, 16, 562–563. [Google Scholar] [CrossRef] [PubMed]
- Woegerbaue, M.; Kuffner, M.; Domingues, S.; Nielsen, K.M. Involvement of aph(3’)-IIa in the formation of mosaic aminoglycoside resistance genes in natural environments. Front. Microbiol. 2015, 6, 1–12. [Google Scholar]
- James, D.; Sanderson, D.; Varga, A.; Sheveleva, A.; Chirkov, S. Genome sequence analysis of new isolates of the Winona strain of Plum pox virus and the first definitive evidence of intrastrain recombination events. Phytopathology 2016, 106, 407–416. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular Evolutionary Genetics Analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [PubMed]
- Nei, M.; Kumar, S. Molecular Evolution and Phylogenetics; Oxford University Press: New York, NY, USA, 2000. [Google Scholar]
- Jukes, T.H.; Cantor, C.R. Evolution of Protein Molecules. In Mammalian Protein Metabolism; Munro, H.N., Ed.; Academic Press: New York, NY, USA, 1969; pp. 21–132. [Google Scholar]
- Thompson, F.L.; Thompson, C.C.; Naser, S.; Hoste, B.; Vandemeulebroecke, K.; Munn, C.; Bourne, D.; Swings, J. Photobacterium rosenbergii sp. nov. and Enterovibrio coralii sp. nov., vibrios associated with coral bleaching. Int. J. Syst. Evol. Microbiol. 2005, 55, 913–917. [Google Scholar] [CrossRef] [PubMed]
- Sawabe, T.; Kita-Tsukamoto, K.; Thompson, F.L. Inferring the evolutionary history of vibrios by means of multilocus sequence analysis. J. Bacteriol. 2007, 189, 7932–7936. [Google Scholar] [CrossRef] [PubMed]
- Pascual, J.; Macian, M.C.; Arahal, D.R.; Garay, E.; Pujalte, M.J. Multilocus sequence analysis of the central clade of the genus Vibrio by using the 16S rRNA, recA, pyrH, rpoD, gyrB, rctB and toxR genes. Int. J. Syst. Evol. Microbiol. 2010, 60, 154–165. [Google Scholar] [CrossRef] [PubMed]
- Urbanczyk, H.; Ast, J.C.; Dunlap, P.V. Phylogeny, genomics and symbiosis of Photobacterium. FEMS Microbiol. Rev. 2011, 35, 324–342. [Google Scholar] [CrossRef] [PubMed]
- Thyssen, A.; Ollevier, F. Genus II. Photobacterium. In Bergey’s Manual of Systematic Bacteriology, 2nd ed.; The Proteobacteria, Part B The Gammaproteobacteria; Brenner, D.L., Krieg, N.R., Staley, J.T., Eds.; Springer: New York, NY, USA, 2005; Volume 2, pp. 546–562. [Google Scholar]
- Seo, H.J.; Bae, S.S.; Lee, J.H.; Kim, S.J. Photobacterium frigidiphilum sp. nov., a psychrophilic, lipolytic bacterium isolated from deep-sea sediments of Edison Seamount. Int. J. Syst. Evol. Microbiol. 2005, 55, 1661–1666. [Google Scholar] [CrossRef] [PubMed]
- Nogi, Y.; Masui, N.; Kato, C. Photobacterium profundum sp. nov., a new, moderately barophilic bacterial species isolated from a deep-sea sediment. Extremophiles 1998, 2, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Yoon, J.H.; Lee, J.K.; Kim, Y.O.; Oh, T.K. Photobacterium lipolyticum sp. nov., a bacterium with lipolytic activity isolated from the Yellow Sea in Korea. Int. J. Syst. Evol. Microbiol. 2005, 55, 335–339. [Google Scholar] [CrossRef] [PubMed]
- Ast, J.C.; Cleenwerck, I.; Engelbeen, K.; Urbanczyk, H.; Thompson, F.L.; De Vos, P.; Dunlap, P.V. Photobacterium kishitanii sp. nov., a luminous marine bacterium symbiotic with deep-sea fishes. Int. J. Syst. Evol. Microbiol. 2007, 57, 2073–2078. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Species | Habitats | Geographic Sources |
|---|---|---|
| P. aestuarii | Tidal flat sediment | Yeongam Bay (R. Korea) |
| P. alginatilyticum | Bottom seawater | East China Sea |
| P. angustum | Seawater | North Pacific Ocean (20°30′ N 157°30′ E) |
| P. aphoticum | Seawater | Malvarrosa beach, Valencia (Spain) |
| P. aplysiae | Eggs of sea hare (Aplysia kurodai) | Mogiyeo (R. Korea) |
| P. aquae | Malabar grouper (Epinephelus malabaricus) in mariculture system | Tianjin (China) |
| P. aquimaris | Seawater | Sagami Bay (Japan) |
| P. carnosum | Packaged poultry meat | Germany |
| P. damselae | Damselfish (Chromis punctipinnis) skin ulcer a, white perch (Roccus americanus) | California, Chesapeake Bay (USA) |
| P. frigidiphilum | Deep-sea sediments (1450 m) | Edison Seamount (western Pacific Ocean) |
| P. gaetbulicola | Tidal flat | Gung harbour (R. Korea) |
| P. galatheae | Mussel | Solomon Sea (Solomon Islands) |
| P. ganghwense | Seawater | Ganghwa Island (R. Korea) |
| P. halotolerans | Water from a subterranean saline lake | Lake Martel, Mallorca (Spain) |
| P. iliopiscarium | Intestines of fish (herring, coal fish, cod and salmon) living in cold seawater | Norway |
| P. indicum | Marine mud (400 m depth) | Indian Ocean |
| P. jeanii | Healthy corals (Palythoa caribaeorum, Phyllogorgia dilatata and Merulina ampliata) | Brazil and Australia |
| P. kishitanii | Light organs and skin of several marine fish species | Japan, Cape Verde, Hawaii, Florida, South Africa |
| P. leiognathi | Light organ of teleostean fish (Leiognathus) | Gulf of Thailand (Thailand) |
| P. lipolyticum | Intertidal sediment | Yellow Sea (R. Korea) |
| P. lutimaris | Tidal flat sediment | Saemankum (R. Korea) |
| P. panuliri | Eggs of spiny lobster (Panulirus penicillatus) | Andaman Sea (India) |
| P. phosphoreum | Skin of marine animals, intestines of marine fish, luminous organs, seawater | Hawaii (USA), Japan and other locations |
| P. piscicola | Skin and intestine of marine fish, spoiled packed cod | North Sea (The Netherlands), Denmark, Aberdeen Bay (UK) |
| P. profundum | Deep-sea sediment (5110 m) | Ryukyu Trench (24°15.23′ N 126°47.30′ E) |
| P. proteolyticum | Ocean sediment | Laizhou Bay (China) |
| P. rosenbergii | Tissue and water extracts of coral species | Magnetic Island (Australia) |
| P. sanctipauli | Coral (Madracis decactis) | St. Peter & St. Paul Archipelago (Brazil) |
| P. sanguinicancri | Crab (Maja brachydactyla) haemolymph, mussels (Mytilus edulis) | Spain, Netherlands |
| P. swingsii | Pacific oysters (Crassostrea gigas), crab (Maja brachydactyla) haemolymph | Mexico, Spain |
| P. toruni | Diseased redbanded seabream (Pagrus auriga) | Spain |
| Genes | D | Ph | P | G | L |
|---|---|---|---|---|---|
| 16S rRNA | |||||
| D | 99.6 | ||||
| Ph | 97.0 | 99.9 | |||
| P | 96.3 | 97.6 | 98.4 | ||
| G | 96.3 | 97.1 | 96.2 | 97.8 | |
| L | 96.9 | 98.3 | 97.1 | 97.3 | 99.2 |
| gyrB | |||||
| D | 83.2 | ||||
| Ph | 70.7 | 91.7 | |||
| P | 68.5 | 77.7 | 81.0 | ||
| G | 65.9 | 73.8 | 72.7 | 79.1 | |
| L | 69.3 | 80.2 | 70.6 | 73.6 | 87.5 |
| gapA | |||||
| D | 91.4 | ||||
| Ph | 87.8 | 98.1 | |||
| P | 86.1 | 91.0 | 88.4 | ||
| G | NA | NA | NA | 75.0 | |
| L | 87.4 | 90.3 | 90.9 | NA | 96.9 |
| topA | |||||
| D | 84.5 | ||||
| Ph | 72.0 | 89.1 | |||
| P | 69.9 | 60.9 | 79.4 | ||
| G | NA | NA | NA | 70.5 | |
| L | 40.2 | 45.9 | 46.7 | NA | 15.8 |
| ftsZ | |||||
| D | 99.4 | ||||
| Ph | 52.6 | 44.0 | |||
| P | 82.3 | 52.9 | 80.1 | ||
| G | 2.5 | 14.0 | 4.9 | 15.2 | |
| L | 58.1 | 44.0 | 58.5 | 12.0 | 38.8 |
| mreB | |||||
| D | 98.2 | ||||
| Ph | 47.0 | NA | |||
| P | 84.9 | 43.2 | 83.3 | ||
| G | 25.1 | 20.5 | 22.9 | NA | |
| L | 84.4 | 43.1 | 86.5 | 22.1 | 90.5 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Labella, A.M.; Castro, M.D.; Manchado, M.; Borrego, J.J. Description of New and Amended Clades of the Genus Photobacterium. Microorganisms 2018, 6, 24. https://doi.org/10.3390/microorganisms6010024
Labella AM, Castro MD, Manchado M, Borrego JJ. Description of New and Amended Clades of the Genus Photobacterium. Microorganisms. 2018; 6(1):24. https://doi.org/10.3390/microorganisms6010024
Chicago/Turabian StyleLabella, Alejandro M., M. Dolores Castro, Manuel Manchado, and Juan J. Borrego. 2018. "Description of New and Amended Clades of the Genus Photobacterium" Microorganisms 6, no. 1: 24. https://doi.org/10.3390/microorganisms6010024
APA StyleLabella, A. M., Castro, M. D., Manchado, M., & Borrego, J. J. (2018). Description of New and Amended Clades of the Genus Photobacterium. Microorganisms, 6(1), 24. https://doi.org/10.3390/microorganisms6010024