Phoma Infections: Classification, Potential Food Sources, and Their Clinical Impact

Abstract
:1. Introduction
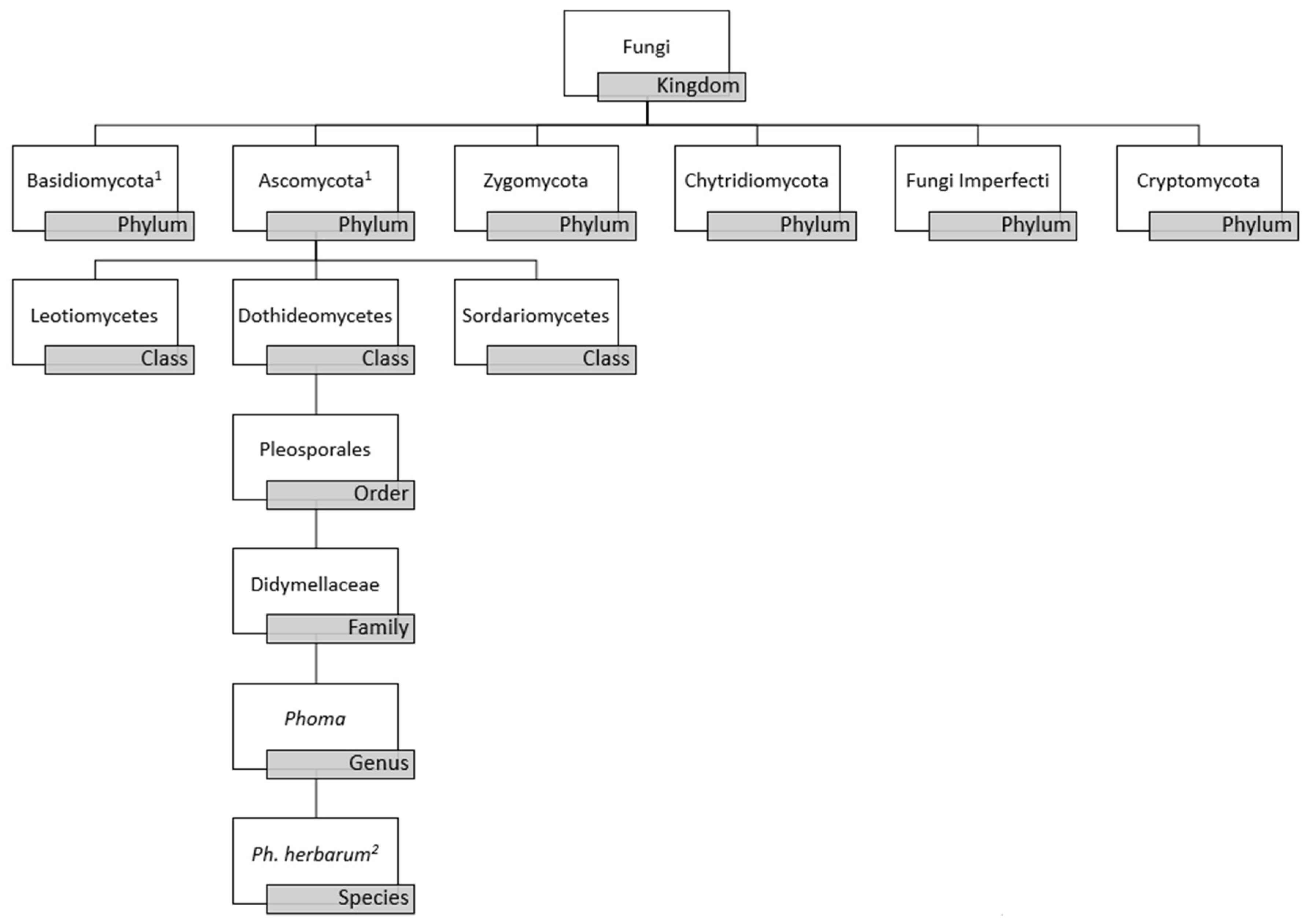
2. Morphology and Molecular-Based Taxonomy
3. Ecological Distribution
4. Clinical Significance
5. Management
6. Discussion
Author Contributions
Funding
Conflicts of Interest
References
- Parr, C.S.; Wilson, N.; Leary, P.; Schulz, K.S.; Lans, K.; Walley, L.; Hammock, J.A.; Goddard, A.; Rice, J.; Studer, M.; et al. The Encyclopedia of Life Vol.2: Providing Global Access to Knowledge about Life on Earth. Biodivers. Data J. 2014, 2, e1079. [Google Scholar] [CrossRef] [PubMed]
- Valenzuela-Lopez, N.; Cano-Lira, J.F.; Guarro, J.; Sutton, D.A.; Wiederhold, N.; Crous, P.W.; Stchigel, A.M. Coelomycetous Dothideomycetes with emphasis on the families Cucurbitariaceae and Didymellaceae. Stud. Mycol. 2018, 90, 1–69. [Google Scholar] [CrossRef] [PubMed]
- Jones, M.D.M.; Forn, I.; Gadelha, C.; Egan, M.J.; Bass, D.; Massana, R.; Richards, T.A. Discovery of novel intermediate forms redefines the fungal tree of life. Nature 2011, 474, 200–203. [Google Scholar] [CrossRef] [PubMed]
- Kwong-Chung, K.J.; Bennett, J.E. Medical Mycology; Lea & Febiger: Philadelphia, PA, USA, 1992; pp. 661–662. ISBN 0812114639. [Google Scholar]
- Boerema, G.H.; Bollen, G.J. Conidiogenesis and conidial septation as differentiating criteria between Phoma and Ascochyta. Persoonia 1975, 8, 111–144. [Google Scholar]
- Boerema, G.H.; de Gruyter, J.; Noordeloos, M.E.; Hamers, M.E.C. Phoma Identification Manual. Differentiation of Specific and Infra-Specific Taxa in Culture; CABI Publishing: Wallingford, UK, 2004; pp. 1–480. ISBN 9780851997438. [Google Scholar]
- Guégan, S.; Garcia-Hermoso, D.; Sitbon, K.; Ahmed, S.; Moguelet, P.; Dromer, F.; Lortholary, O.; French Mycosis Study Group. Ten-Year Experience of Cutaneous and/or Subcutaneous Infections Due to Coelomycetes in France. Open Forum Infect. Dis. 2016, 3, ofw106. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.J.; Kim, S.H. A genome tree of life for the fungi kingdom. Proc. Natl. Acad. Sci. USA 2017, 114, 9391–9396. [Google Scholar] [CrossRef] [PubMed]
- Aveskamp, M.M.; de Gruyter, J.; Crous, P.W. Biology and recent developments in the systematics of Phoma, a complex genus of major quarantine significance. Fungal Divers. 2008, 31, 1–18. [Google Scholar]
- Young, N.A.; Kwon-chung, K.J.; Freeman, J. Subcutaneous abscess caused by Phoma sp. resembling Pyrenochaeta romeroi: Unique fungal infection occurring in immunosuppressed recipient of renal allograft. Am. J. Clin. Pathol. 1973, 59, 810–816. [Google Scholar] [CrossRef] [PubMed]
- Nucci, M.; Queiroz-Telles, F.; Tobón, A.M.; Restrepo, A.; Colombo, A.L. Epidemiology of opportunistic fungal infections in Latin America. Clin. Infect. Dis. 2010, 51, 561–570. [Google Scholar] [CrossRef] [PubMed]
- Weidenbörner, M.; Hindorf, H. Fungi isolated from protein enriched seeds and pods with special emphasis on the genus Aspergillus. Sci. Technol. 1989, 17, 383–390. [Google Scholar]
- Pitt, J.I.; Hocking, A.D.; Bhudhasamai, K.; Miscamble, B.F.; Wheeler, K.A.; Tanboon-Ek, P. The normal mycoflora of commodities from Thailand. 1. Nuts and oilseeds. Int. J. Food Microbiol. 1993, 20, 211–226. [Google Scholar] [CrossRef]
- Huang, L.H.; Hanlin, R.T. Fungi occurring in freshly harvested and in-market pecans. Mycologia 1975, 67, 689–700. [Google Scholar] [CrossRef] [PubMed]
- Joffe, A.Z. The mycoflora of fresh and stored groundnut kernels in Israel. Mycopathol. Mycol. Appl. 1969, 39, 255–264. [Google Scholar] [CrossRef]
- Ko¨vics, G.J.; de Gruyter, J.; van der Aa, H.A. Phoma sojicola comb. nov. and other hyaline-spored coelomycetes pathogenic on soybean. Mycol. Res. 1999, 103, 1065–1070. [Google Scholar] [CrossRef]
- Temorshuizen, A.J. Fungal and fungus-like pathogens of potato. In Potato Biology and Biotechnology; Elsevier Science B.V.: Amsterdam, The Netherlands, 2007; pp. 643–665. [Google Scholar]
- Paterson, R.R.M.; Lima, N. Filamentous Fungal Human Pathogens from Food Emphasising Aspergillus, Fusarium and Mucor. Microorganisms 2017, 5, 44. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, R.C.; Goncalves, S.S.; Oliveira, M.S.; Dilkin, P.; Mallmann, C.A.; Freitas, R.S.; Bianchi, P.; Correa, B. Natural occurrence of tenuazonic acid and Phoma sorghina in Brazilian sorghum grains at different maturity stages. Food Chem. 2017, 230, 491–496. [Google Scholar] [CrossRef] [PubMed]
- Do Amaral, A.L.; de Carli, M.L.; Neto, J.F.B.; Dal Soglio, F.K. Phoma sorghina, a new pathogen associated with Phaeosphaeria leaf spot on maize in Brazil. Plant Pathol. J. 2004, 53, 259. [Google Scholar] [CrossRef]
- Adekoya, I.; Obadina, A.; Adaku, C.C.; De Boevre, M.; Okoth, S.; De Saeger, S.; Njobeh, P. Mycobiota and co-occurrence of mycotoxins in South African maize-based opaque beer. Int. J. Food Microbiol. 2018, 270, 22–30. [Google Scholar] [CrossRef] [PubMed]
- Kim, G.H.; Kim, D.R.; Park, S.; Lee, Y.S.; Jung, J.S.; Koh, Y.J. Incidence rates of major diseases of kiwiberry in 2015 and 2016. Plant Pathol. J. 2017, 33, 434–439. [Google Scholar] [CrossRef] [PubMed]
- Migheli, Q.; Cacciola, S.O.; Balmas, V.; Pane, A.; Ezra, D.; Magnano di San Lio, G. Mal secco disease caused by Phoma tracheiphila: A potential threat to lemon production worldwide. Plant Dis. 2009, 93, 852–867. [Google Scholar] [CrossRef]
- Kubota, M.; Kishi, K.; Abiko, K. Phoma leaf spot, stem and fruit rot of tomato caused by Phoma lycopersici Cooke in Japan. Jpn. J. Phytopathol. 2000, 66, 12–17. [Google Scholar] [CrossRef]
- Laundon, G.F. Records of fungal plant disease in New Zealand. N. Z. J. Bot. 1971, 9, 610–624. [Google Scholar] [CrossRef]
- Palavouzis, S.; Tzamos, S.; Paplomatas, E.; Thomidis, T. First report of Phoma Aliena causing fruit rots of pomegranates in northern Greece. J. Plant Pathol. 2015, 97, 215. [Google Scholar]
- Cole, R.J.; Jarvis, B.B.; Schweikert, M.A. Handbook of Secondary Fungal Metabolites, 3rd ed.; Academic Press: Amsterdam, The Netherlands, 2003. [Google Scholar]
- Lugauskas, A.; Raila, A.; Railiene, M.; Raudoniene, V. Toxic micromycetes in grain raw material during its processing. Ann. Agric. Environ. Med. 2006, 13, 147–161. [Google Scholar] [PubMed]
- Weidenbörner, M. Pine nutes: The mycobiota and potential mycotoxins. Can. J. Microbiol. 2001, 47, 460–463. [Google Scholar] [CrossRef] [PubMed]
- Visconti, A.; Logrieco, A.; Vurro, M.; Bottalico, A. Tenuazonic acid in blackmold tomatoes: Occurrence, production by associated Alternaria species, and phytotoxic properties. Phytopathol. Mediterran. 1987, 26, 125–128. [Google Scholar]
- Yekeler, H.; Bitmis, K.; Özcelik, N.; Doymaz, M.Z.; Metin, C. Analysis of toxic effects of Alternaria toxins on esophagus of mice by light and electron microscopy. Toxicol. Pathol. 2001, 29, 492–497. [Google Scholar] [CrossRef] [PubMed]
- Sutton, D.A.; Rinaldi, M.G.; Sanche, S.E. Dematiaceous fungi. In Clinical Mycology, 2nd ed.; Anaissie, E.J., McGinnis, M.R., Pfaller, M.A., Eds.; Churchill Livingstone (Elsevier): London, UK, 2009; p. 342. ISBN 9781416056805. [Google Scholar]
- Rai, M.K.; Tiwari, V.V.; Irinyi, L.; Kövics, G.J. Advances in taxonomy of genus Phoma: Polyphyletic nature and role of phenotypic traits and molecular systematics. Indian J. Microbiol. 2014, 54, 123–128. [Google Scholar] [CrossRef] [PubMed]
- Valenzuela-Lopez, N.; Sutton, D.A.; Cano-Lira, J.F.; Paredes, K.; Wiederhold, N.; Guarro, J.; Stchigel, A.M. Coelomycetous Fungi in the Clinical Setting: Morphological Convergence and Cryptic Diversity. J. Clin. Microbiol. 2017, 55, 552–567. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Jiang, J.R.; Zhang, G.Z.; Cai, L.; Crous, P.W. Resolving the Phoma enigma. Stud. Mycol. 2015, 82, 137–217. [Google Scholar] [CrossRef] [PubMed]
- Babič, M.N.; Gunde-Cimerman, N.; Vargha, M.; Tischner, Z.; Magyar, D.; Veríssimo, C.; Sabino, R.; Viegas, C.; Meyer, W.; Brandão, J. Fungal Contaminants in Drinking Water Regulation? A Tale of Ecology, Exposure, Purification and Clinical Relevance. Int. J. Environ. Res. Public Health 2017, 14, 636. [Google Scholar] [CrossRef]
- Doggett, M.S. Characterization of Fungal Biofilms within a Municipal Water Distribution System. Appl. Environ. Microbiol. 2000, 66, 1249–1251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aho, R.; Hirn, J. A survey of fungi and some indicator bacteria in chlorinated water of indoor public swimming pools. Zentralbl. Bakteriol. Mikrobiol. Hyg. B 1981, 173, 242–249. [Google Scholar] [PubMed]
- Ekowati, Y.; van Diepeningen, A.D.; Ferrero, G.; Kennedy, M.D.; de Roda Husman, A.M.; Schets, F.M. Clinically relevant fungi in water and on surfaces in an indoor swimming pool facility. Int. J. Hyg. Environ. Health 2017, 220, 1152–1160. [Google Scholar] [CrossRef] [PubMed]
- Hamada, N.; Abe, N. Growth characteristics of four fungal species in bathrooms. Biocontrol. Sci. 2010, 15, 111–115. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Wang, N.F.; Zhang, Y.Q.; Liu, H.Y.; Yu, L.Y. Diversity and Distribution of Aquatic Fungal Communities in the Ny-Ålesund Region, Svalbard (High Arctic): Aquatic Fungi in the Arctic. Microb. Ecol. 2016, 71, 543–554. [Google Scholar] [CrossRef] [PubMed]
- Calvo, M.A.; Guarro, J.; Suarez, G.; Ramírez, C. Air-borne fungi in the air of Barcelona (Spain). IV. Various isolated genera. Mycopathologia 1980, 71, 119–123. [Google Scholar] [CrossRef] [PubMed]
- A’Hara, D. Detection and identification of Phoma pathogens of potato. Methods Mol. Biol. 2015, 1302, 17–27. [Google Scholar] [PubMed]
- Food and Agriculture Organization. Revised Guidelines for the Assessment of Microbiological Quality of Processed Foods. 2013. Available online: http://fda.gov.ph./attachments/article/17218/FC2013-010.pdf (accessed on 13 June 2018).
- Bandyopadhyay, R.; Mughogho, L.K.; Satyanarayana, M.V.; Kalisz, M.E. Occurrence of airborne spores of fungi causing grain mould over sorghum crop. Mycol. Res. 1991, 95, 1315–1320. [Google Scholar] [CrossRef]
- González, H.H.; Martínez, E.J.; Resnik, S.L. Fungi associated with sorghum grain from Argentina. Mycopathologia 1997, 46, 1420–1422. [Google Scholar]
- Hussaini, A.M.; Timothy, A.G.; Olufunmilayo, H.A.; Ezekiel, A.S.; Godwin, H.O. Fungi and some mycotoxins found in mouldy Sorghum in Niger state, Nigeria. World J. Agric. Sci. 2009, 5, 5–17. [Google Scholar]
- Food and Agriculture Organization of the United Nations. 2010–2017. Available online: https://www.fao.org/giews/countrybrief/country.jsp?code=BRA (accessed on 14 June 2018).
- Janke, D. About a human-pathogenic new species of Peyronellaea bred from lung changes. Mycopathologia 1956, 7, 229–240. [Google Scholar]
- Bakerspigel, A. The isolation of Phoma hibernica from lesions on a leg. Sabouraudia 1970, 7, 261–264. [Google Scholar] [CrossRef] [PubMed]
- Bakerspigel, A.; Lowe, D.; Rostas, A. The isolation of Phoma eupyrena from a human lesion. Arch. Dermatol. 1981, 117, 362–363. [Google Scholar] [CrossRef] [PubMed]
- Roehm, C.E.; Salazar, J.C.; Hagstrom, N.; Valdez, T.A. Phoma and Acremonium invasive fungal rhinosinusitis in congenital acute lymphocytic leukemia and literature review. Int. J. Pediatr. Otorhinolaryngol. 2012, 76, 1387–1391. [Google Scholar] [CrossRef] [PubMed]
- Gordon, M.A.; Salkin, I.F.; Stone, W.B. Phoma (Peyronellaea) as zoopathogen. Sabouraudia 1975, 13, 329–333. [Google Scholar] [CrossRef] [PubMed]
- Punithalingam, E. Sphaeropsidales in culture from humans. Nova Hedwig. 1979, 31, 119–158. [Google Scholar]
- Shukla, N.P.; Rajak, R.K.; Agarwasl, G.P.; Gupta, D. Phoma minutispora as a human pathogen. Mykosen 1984, 27, 255–258. [Google Scholar] [CrossRef] [PubMed]
- Baker, J.G.; Salkin, I.F.; Forgacs, P.; Haines, J.H.; Kemna, M.E. First report of subcutaneous phaeohyphomycosis of the foot caused by Phoma minutella. J. Clin. Microbiol. 1987, 25, 2395–2397. [Google Scholar] [PubMed]
- Rai, M.K. Phoma sorghina infection in human being. Mycopathologia 1989, 105, 167–170. [Google Scholar] [CrossRef] [PubMed]
- Hirsh, A.H.; Schiff, T.A. Subcutaneous phaeohyphomycosis caused by an unusual pathogen: Phoma species. J. Am. Acad. Dermatol. 1996, 34, 679–680. [Google Scholar] [CrossRef]
- Rosen, T.; Rinaldi, M.G.; Tschen, J.A.; Stern, J.K.; Cernoch, P. Cutaneous lesions due to Pleurophoma (Phoma) Complex. South Med. J. 1996, 89, 431–434. [Google Scholar] [CrossRef] [PubMed]
- Zaitz, C.; Heins-Vaccari, E.M.; de Freitas, R.S.; Arriagada, G.L.; Ruiz, L.; Totoli, S.A.; Marques, A.C.; Rezze, G.G.; Müller, H.; Valente, N.S.; et al. Subcutaneous phaeohyphomycosis caused by Phoma cava. Report of a case and review of the literature. Rev. Inst. Med. Trop. Sao Paulo 1997, 39, 43–48. [Google Scholar] [CrossRef] [PubMed]
- Arrese, J.E.; Piérard-Franchimont, C.; Piérard, G.E. Unusual mould infection of the human stratum corneum. J. Med. Vet. Mycol. 1997, 35, 225–227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oh, C.K.; Kwon, K.S.; Lee, J.B.; Jang, H.S.; Chung, T.A.; Suh, S.B. Subcutaneous phaeohyphomycosis caused by Phoma species. Int. J. Dermatol. 1999, 38, 874–876. [Google Scholar] [PubMed]
- Everett, J.E.; Busick, N.P.; Sielaff, T.; Wahoff, D.C.; Dunn, D.L. A deeply invasive Phoma species infection in a renal transplant recipient. Transplant. Proc. 2003, 35, 1387–1389. [Google Scholar] [CrossRef]
- Suh, M.K. Phaeohyphomycosis in Korea. Nihon Ishinkin Gakkai Zasshi 2005, 46, 67–70. [Google Scholar] [CrossRef] [PubMed]
- Vasoo, S.; Yong, L.K.; Sultania-Dudani, P.; Scorza, M.L.; Sekosan, M.; Beavis, K.G.; Huhn, G.D. Phaeomycotic cysts caused by Phoma species. Diagn. Microbiol. Infect. Dis. 2011, 70, 531–533. [Google Scholar] [CrossRef] [PubMed]
- Schieffelin, J.S.; Garcia-Diaz, J.B.; Loss, G.E., Jr.; Beckman, E.N.; Keller, R.A.; Staffeld-Coit, C.; Garces, J.C.; Pankey, G.A. Phaeohyphomycosis fungal infections in solid organ transplant recipients: Clinical presentation, pathology, and treatment. Transpl. Infect. Dis. 2014, 16, 270–278. [Google Scholar] [CrossRef] [PubMed]
- Hernández-Hernández, F.; Vargas-Arzola, J.; Ríos-Cruz, O.P.; Córdova-Martínez, E.; Manzano-Gayosso, P.; Segura-Salvador, A. First case of chromoblastomycosis due to Phoma insulana. Enferm. Infecc. Microbiol. Clin. 2018, 36, 95–99. [Google Scholar] [CrossRef] [PubMed]
- Rishi, K.; Font, R.L. Keratitis caused by an unusual fungus, Phoma species. Cornea 2003, 22, 166–168. [Google Scholar] [CrossRef] [PubMed]
- Errera, M.H.; Barale, P.O.; Nourry, H.; Zamfir, O.; Guez, A.; Warnet, J.M.; Sahel, J.A.; Chaumeil, C. Usefulness of voriconazole in treatment of Phoma glomerata after penetrating injury. J. Fr. Ophthalmol. 2008, 31, 62–66. [Google Scholar] [CrossRef]
- Kumar, P.; Thomas, S.; Papagiannuli, E.; Hardman, S.C.; Jenkins, D.; Prydal, J. A case of Phoma fungal keratitis in a contact lens user. JRSM Open 2015, 6. [Google Scholar] [CrossRef] [PubMed]
- McElnea, E.; Farrell, S.; Lynch, B.; Bishop, K.; Mullen, D.; Borman, A.; Higgins, G. A rare case of fungal keratitis: Diagnosis and management. JMM Case Rep. 2015, 2, e000091. [Google Scholar] [CrossRef]
- Morris, J.T.; Beckius, M.L.; Jeffery, S.; Longfeld, R.N.; Heaven, R.F.; Baker, W.J. Lung mass caused by Phoma species. Infect. Dis. Clin. Pract. 1995, 4, 58–59. [Google Scholar] [CrossRef]
- Balis, E.; Velegraki, A.; Fragou, A.; Pefanis, A.; Kalabokas, T.; Mountokalakis, T. Lung mass caused by Phoma exigua. Scand. J. Infect. Dis. 2006, 38, 552–555. [Google Scholar] [CrossRef] [PubMed]
- Tullio, V.; Banche, G.; Allizond, V.; Roana, J.; Mandras, N.; Scalas, D.; Panzone, M.; Cervetti, O.; Valle, S.; Carlone, N.; et al. Non-dermatophyte moulds as skin and nail foot mycosis agents: Phoma herbarum, Chaetomium globosum and Microascus cinereus. Fungal Biol. 2010, 114, 345–349. [Google Scholar] [CrossRef] [PubMed]
- Punithalingam, E. Phoma oculohominis sp. nov. from corneal ulcer. Trans. Br. Mycol. Soc. 1976, 67, 142–143. [Google Scholar] [CrossRef]
- Chowdhary, A.; Meis, J.F.; Guarro, J.; de Hoog, G.S.; Kathuria, S.; Arendrup, M.C.; Arikan-Akdagli, S.; Akova, M.; Boekhout, T.; Caira, M.; et al. ESCMID and ECMM joint clinical guidelines for the diagnosis and management of systemic phaeohyphomycosis: Diseases caused by black fungi. Clin. Microbiol. Infect. 2014, 20, 47–75. [Google Scholar] [CrossRef] [PubMed]
- Sutton, D.A. Coelomycetous fungi in human disease. A review: Clinical entities, pathogenesis, identification and therapy. Rev. Iberoam. Micol. 1999, 16, 171–191. [Google Scholar] [PubMed]
- Brenier-Pinchart, M.P.; Faure, O.; Garban, F.; Fricker-Hidalgo, H.; Mallaret, M.R.; Trens, A.; Lebeau, B.; Pelloux, H.; Grillot, R. Ten-year surveillance of fungal contamination of food within a protected haemotological unit. Mycoses 2006, 49, 421–425. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Isolated Fungus | Gender/Age | Source/History | Immunosuppression | Treatment/Outcome | Reference |
|---|---|---|---|---|---|
| Phoma spp. | N/A | Pulmonary | N/A | N/A | Janke, D. et al. 1956 [49] |
| Phoma hibernica | F/22 | Skin (deep leg) | Topical steroids | Oral griseofulvin/clinical improvement | Bakerspigel, A. 1970 [50] |
| Phoma spp. | F/42 | Skin (deep heel) | Azathioprine; prednisone; s/p renal transplant | Debridement/resolved | Young, N.A. et al. 1973 [10] |
| Phoma cava | M/4 | Skin (superficial ear) | Otherwise healthy | Oral griseofulvin; corticosteroid/resolved | Gordon, M.A. et al. 1975 [53] |
| Phoma oculo hominis | N/A | Eye (Corneal ulcer) | Otherwise healthy | N/A | Punithalingam, E. 1976 [75] |
| Phoma cruris- hominis | F/? | Subcutaneous | N/A | N/A | Punithalingam, E. 1979 [54] |
| Phoma eupyrena | M/18 mos. | Skin (perioral lesions) | Otherwise healthy | Clotrimazole; 15% zinc oxide paste; Dimethicone/resolved | Bakerspigel, A. et al. 1981 [51] |
| Phoma minutispora Phoma minutispora | M/18 M/20 | Skin (face) Skin (neck) | Typhoid fever Oral steroids | Topical clotrimazole/resolved Topical clotrimazole/resolved | Shukla, N.P. et al. 1984 [55] |
| Phoma minutella | M/75 | Skin (deep foot) Farmer from Dominican Republic | Steroid therapy Diabetes mellitus | Debridement; amputation for secondary gangrene/resolved | Baker, J.G. et al. 1987 [56] |
| Phoma sorghina Phoma sorghina | M/24 M/19 | Skin (face, neck, hands) Skin (face) | Otherwise healthy Otherwise healthy | Topical miconazole/resolved Topical miconazole/resolved | Rai, M.K. 1989 [57] |
| Phoma spp. | F/24 | Pulmonary (lung mass) | Acute Lymphocytic Leukemia; chemotherapy | Left lower lobectomy Amphotericin B/resolved | Morris, J.T. et al. 1995 [72] |
| Phoma spp. | M/45 | Skin (deep/hands) | Otherwise healthy | Itraconazole; ketoconazole/clinical improvement | Hirsh, A.H. et al. 1996 [58] |
| Phoma spp. | F/24 | Skin (deep face) | Topical steroids | Ketoconazole/resolved | Rosen, T. et al. 1996 [59] |
| Phoma cava | M/63 | Skin (deep hand) | Pulmonary sarcoidosis; oral steroids | Amphotericin B; itraconazole/resolved | Zaitz, C. et al. 1997 [60] |
| Phoma spp. Phoma spp. | M/49 M/53 | Skin (plantar; foot) Skin (plantar; foot) [Both Phoma and Scopulariopsis brevicaulis grew from the strateum corneum] | Atopic dermatitis Atopic dermatitis | Topical bifonazole and ketoconazole/No improvement; lost to follow up Topical bifonazole and ketoconazole/no improvement; lost to follow up | Arrese, J.E. et al. 1997 [61] |
| Phoma spp. | M/77 | Skin (deep) | Otherwise healthy | Itraconazole/resolved | Oh, C.K. et al. 1999 [62] |
| Phoma spp. | M/72 | Eye (keratitis) Globe trauma | Otherwise healthy | Debridement; keratectomy | Rishi, K. et al. 2003 [68] |
| Phoma spp. | F/50 | Skin (deep hand) | s/p renal transplant | Surgical debridement; amphotericin B/resolved | Everett, J.E. et al. 2003 [63] |
| Phoma spp. | M/19 | Skin (deep face) | N/A | Amphotericin B | Suh, M.K. 2005 [64] |
| Phoma exigua | M/68 | Pulmonary | Acute myeloid leukemia; Diabetes mellitus | Amphotericin B; left pneumonectomy/death | Balis, E. et al. 2006 [73] |
| Phoma glomerata | M/32 | Eye (endophthalmitis) Retinal detachment surgery after penetrating globe injury | None noted | Amphotericin (intravitreal); voriconazole (intravitreal)/resolved | Errera, M.H. et al. 2008 [69] |
| Phoma herbarum | F/36 | Nail, toe [Phoma herbarum, Chaetomium globosum, and Microascus cinereus were isolated] | Otherwise healthy | Allylamine; sertaconazole/resolved | Tullio, V. et al. 2010 [74] |
| Phoma spp. | M/69 | Skin (ganglion cysts on wrist, forearm) | Diabetes mellitus | Oral itraconazole; surgical excision/resolved | Vasoo, S. et al. 2011 [65] |
| Phoma spp. | F/1 mo. | Sinus (invasive rhinosinusitis) [Phoma and Acremonium spp. were isolated] | Acute lymphoblastic leukemia; s/p chemotherapy | Amphotericin B; posaconazole; voriconazole; debridement/death with progressive rhinocerebral extension | Roehm, C.E. et al. 2012 [52] |
| Phoma spp. Phoma spp. | M/45 M/48 | Skin (deep knee) Skin(deep knee) | Diabetes mellitus; s/p liver transplant s/p renal transplant; s/p pancreas transplant | Oral ketoconazole; surgical excision Oral itraconazole; surgical excision | Schieffelin, J.S. et al. 2014 [66] |
| Phoma spp. | F/79 | Eye (keratitis) (Risk factor: used contact lenses) | Otherwise healthy | Oral itraconazole; amphotericin eye (intravitreal); keratoplasty/resolved | Kumar, P. et al. 2015 [70] |
| Phoma spp. | F/59 | Eye (corneal ulcer and abscess) (Risk factor: used contact lenses) | Otherwise healthy | Amphotericin B; (intravitreal); keratoplasty | McElnea, E. et al. 2015 [71] |
| Phoma insulana | M/79 | Skin (deep foot) Foot laceration which evolved over 27 years compatible with chromo- blastomycosis | Chronic alcoholism, smoker | None/lost to follow up | Hernández-Hernández, F. et al. 2018 [67] |
| Organism | Value for the Drug (Microgram/mL) | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| AMB 1 | 5-FC | ITC | VRC | FLC | PSC | KTC | TRB | MFG | CFG | AFG | MON | NAT | |
| Valenzuela-Lopez, N. et al. 2017 [11] | |||||||||||||
| Phoma spp. Range 2 MIC90 | 0.03–4 0.25 | 0.5–16 4 | 0.03–2 0.5 | 0.03–2 1 | -- -- | 0.03–1 0.5 | -- -- | ≤0.03 0.03 | ≤0.03 0.03 | ≤0.03 0.03 | ≤0.03 0.03 | -- -- | -- -- |
| Ph. herbarum Range MIC90 | 0.12–2 1 | 0.5–16 16 | 0.25–4 1 | 0.06–4 1 | -- -- | 0.12–1 1 | -- -- | ≤0.03 0.03 | 0.03–0.06 0.06 | 0.03–0.12 0.12 | 0.03–0.12 0.06 | -- -- | -- -- |
| Sutton, D.A. 1999 [48] | |||||||||||||
| Phoma spp. MIC HIBreakpoints 3 | 1 <1 = S, >2 = R | 16 <16 = S, >32 = R | 16 <0.5 = S, >1 = R | -- | 32 <32 = S, >64 = R | -- | 8 <8 = S, >16 = R | -- | -- | -- | -- | 8 <8 = S, >16 = R | 32 <32 4 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bennett, A.; Ponder, M.M.; Garcia-Diaz, J. Phoma Infections: Classification, Potential Food Sources, and Their Clinical Impact. Microorganisms 2018, 6, 58. https://doi.org/10.3390/microorganisms6030058
Bennett A, Ponder MM, Garcia-Diaz J. Phoma Infections: Classification, Potential Food Sources, and Their Clinical Impact. Microorganisms. 2018; 6(3):58. https://doi.org/10.3390/microorganisms6030058
Chicago/Turabian StyleBennett, Ashely, Michelle M. Ponder, and Julia Garcia-Diaz. 2018. "Phoma Infections: Classification, Potential Food Sources, and Their Clinical Impact" Microorganisms 6, no. 3: 58. https://doi.org/10.3390/microorganisms6030058
APA StyleBennett, A., Ponder, M. M., & Garcia-Diaz, J. (2018). Phoma Infections: Classification, Potential Food Sources, and Their Clinical Impact. Microorganisms, 6(3), 58. https://doi.org/10.3390/microorganisms6030058