Changes in the Bacterioplankton Community Structure from Southern Gulf of Mexico During a Simulated Crude Oil Spill at Mesocosm Scale

 , , ,
, , ,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling and Set-Up of the Mesocosm System
2.2. Quantification of Hydrocarbons in the Water Column
2.3. Estimation of Culturable Hydrocarbonoclastic Bacteria and Quantification of the alkB Gene
2.4. Bacterial Community Analyses
2.5. Bioinformatics and Statistics
3. Results
3.1. Physicochemical Analyses
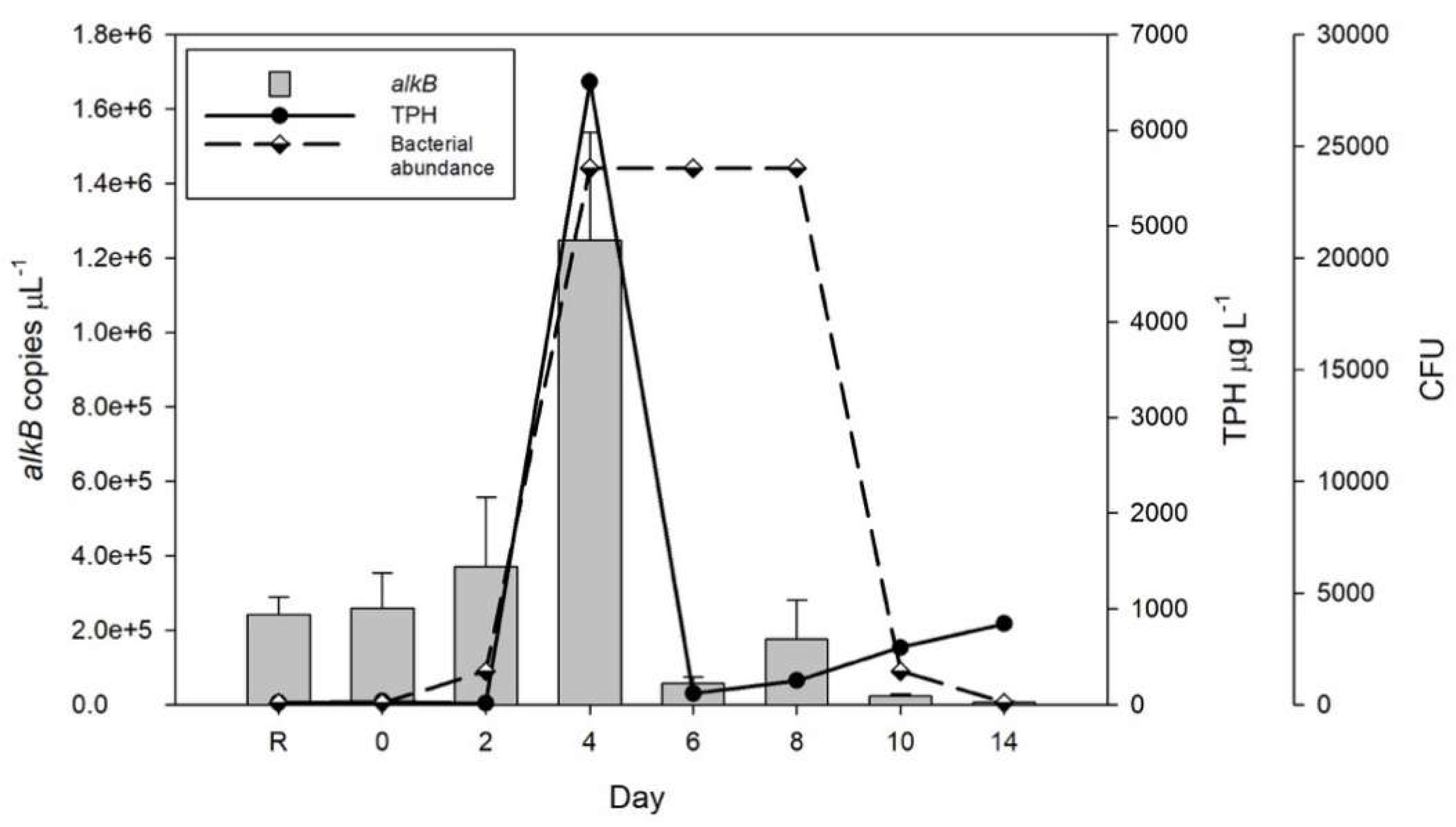
3.2. Estimation of the Bacterial Hydrocarbon Degrading Activity
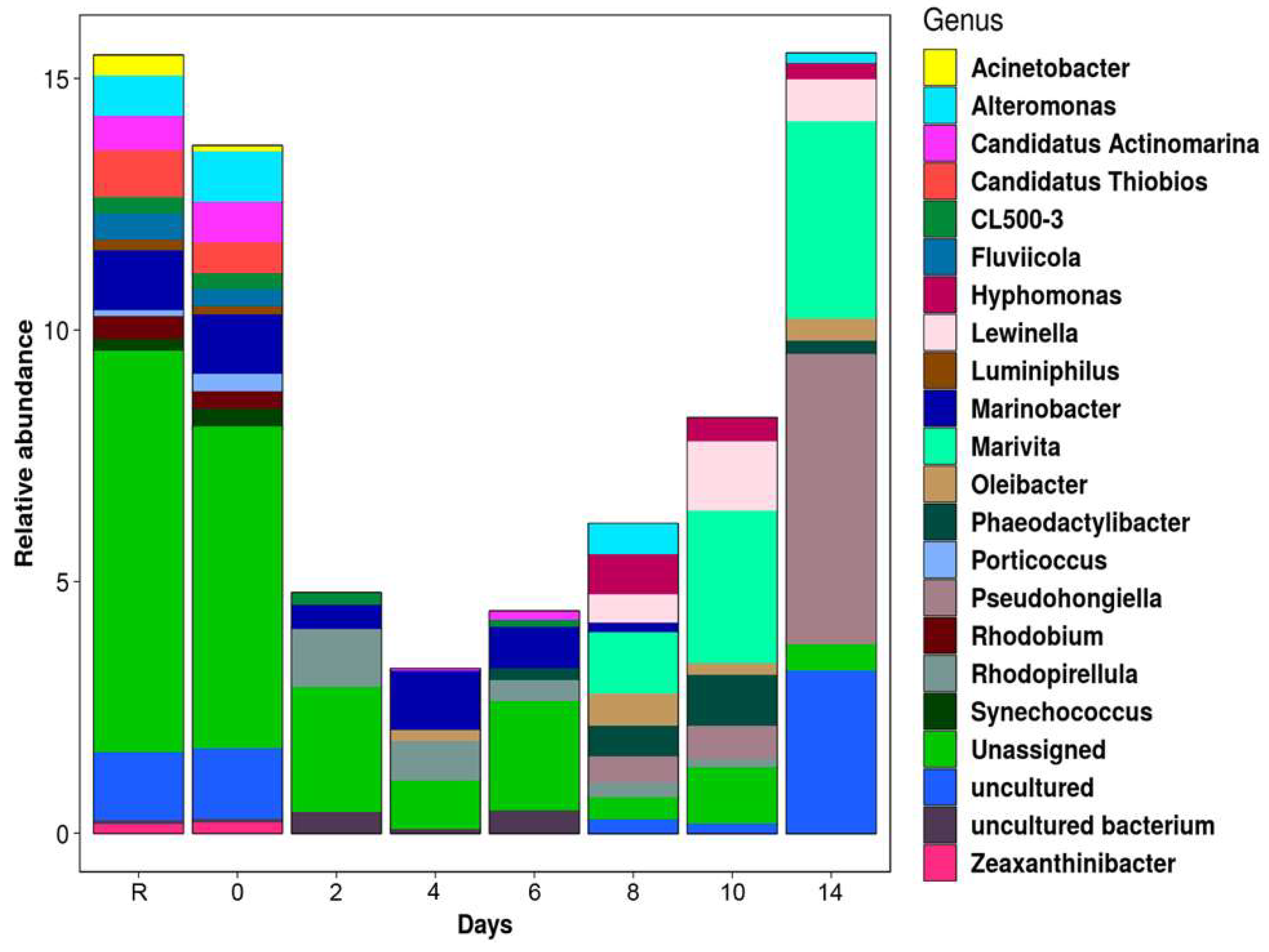
3.3. Bacterial Community Structure and Composition
3.4. Differential Abundance Analysis
4. Discussion
4.1. Shifts in the Bacterioplankton Community after the Simulated Oil Spill
4.2. Hydrocarbonoclastic Bacterial Activity
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Joye, S.B.; Bracco, A.; Özgökmen, T.M.; Chanton, J.P.; Grosell, M.; MacDonald, I.R.; Cordes, E.E.; Montoya, J.P.; Passow, U. The Gulf of Mexico Ecosystem, Six Years after the Macondo Oil Well Blowout. Deep Sea Res. Part II Top. Stud. Oceanogr. 2016, 129, 4–19. [Google Scholar] [CrossRef]
- García-Cruz, N.U.; Aguirre-Macedo, M.L. Biodegradación de Petróleo Por Bacterias: Algunos Casos de Estudio En El Golfo de Mexico. In Golfo de México: Contaminación e Impacto Ambiental, Diagnóstico y Tendencias; Botello, A.V., Rendón von Osten, J., Benítez, J.A., Gold-Bouchot, G., Eds.; UAC, UNAM-ICMYL, CINVESTAV-Unidad Mérida: Mérida, Mexico, 2014; pp. 641–652. [Google Scholar]
- Botello, A.V.; Soto, L.A.; Ponce-Vélez, G.; Susana Villanueva, F. Baseline for PAHs and Metals in NW Gulf of Mexico Related to the Deepwater Horizon Oil Spill. Estuar. Coast. Shelf Sci. 2015, 156, 124–133. [Google Scholar] [CrossRef]
- Mapelli, F.; Scoma, A.; Michoud, G.; Aulenta, F.; Boon, N.; Borin, S.; Kalogerakis, N.; Daffonchio, D. Biotechnologies for Marine Oil Spill Cleanup: Indissoluble Ties with Microorganisms. Trends Biotechnol. 2017, 35, 860–870. [Google Scholar] [CrossRef] [PubMed]
- Soto, L.A.; Botello, A.V.; Licea-Durán, S.; Lizárraga-Partida, M.L.; Yáñez-Arancibia, A. The Environmental Legacy of the Ixtoc-I Oil Spill in Campeche Sound, Southwestern Gulf of Mexico. Front. Mar. Sci. 2014, 1, 57. [Google Scholar] [CrossRef]
- Lindh, M.V.; Pinhassi, J. Sensitivity of Bacterioplankton to Environmental Disturbance: A Review of Baltic Sea Field Studies and Experiments. Front. Mar. Sci. 2018, 5, 361. [Google Scholar] [CrossRef]
- Shade, A.; Peter, H.; Allison, S.D.; Baho, D.L.; Berga, M.; Bürgmann, H.; Huber, D.H.; Langenheder, S.; Lennon, J.T.; Martiny, J.B.H.; et al. Fundamentals of Microbial Community Resistance and Resilience. Front. Microbiol. 2012, 3, 417. [Google Scholar] [CrossRef] [Green Version]
- Hazen, T.C.; Dubinsky, E.A.; DeSantis, T.Z.; Andersen, G.L.; Piceno, Y.M.; Singh, N.; Jansson, J.K.; Probst, A.; Borglin, S.E.; Fortney, J.L.; et al. Deep-Sea Oil Plume Enriches Indigenous Oil-Degrading Bacteria. Science 2010, 330, 204–208. [Google Scholar] [CrossRef] [Green Version]
- Kimes, N.E.; Callaghan, A.V.; Aktas, D.F.; Smith, W.L.; Sunner, J.; Golding, B.T.; Drozdowska, M.; Hazen, T.C.; Suflita, J.M.; Morris, P.J. Metagenomic Analysis and Metabolite Profiling of Deep-Sea Sediments from the Gulf of Mexico Following the Deepwater Horizon Oil Spill. Front. Microbiol. 2013, 4, 50. [Google Scholar] [CrossRef]
- Kimes, N.E.; Callaghan, A.V.; Suflita, J.M.; Morris, P.J. Microbial Transformation of the Deepwater Horizon Oil Spill—Past, Present, and Future Perspectives. Front. Microbiol. 2014, 5, 603. [Google Scholar] [CrossRef]
- King, G.M.; Kostka, J.E.; Hazen, T.C.; Sobecky, P.A. Microbial Responses to the Deepwater Horizon Oil Spill: From Coastal Wetlands to the Deep Sea. Ann. Rev. Mar. Sci. 2015, 7, 377–401. [Google Scholar] [CrossRef]
- Hazen, T.C. Lessons from the 2010 Deepwater Horizon Accident in the Gulf of Mexico. In Hydrocarbons, Oils and Lipids: Diversity, Origin, Chemistry and Fate; Wilkes, H., Ed.; Springer International Publishing: Cham, Switzerland, 2018; pp. 1–19. [Google Scholar] [CrossRef]
- Marietou, A.; Chastain, R.; Beulig, F.; Scoma, A.; Hazen, T.C.; Bartlett, D.H. The Effect of Hydrostatic Pressure on Enrichments of Hydrocarbon Degrading Microbes from the Gulf of Mexico Following the Deepwater Horizon Oil Spill. Front. Microbiol. 2018, 9, 808. [Google Scholar] [CrossRef] [PubMed]
- Kessler, J.D.; Valentine, D.L.; Redmond, M.C.; Du, M.; Chan, E.W.; Mendes, S.D.; Quiroz, E.W.; Villanueva, C.J.; Shusta, S.S.; Werra, L.M.; et al. A Persistent Oxygen Anomaly Reveals the Fate of Spilled Methane in the Deep Gulf of Mexico. Science 2011, 331, 312–315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cappello, S.; Yakimov, M.M. Mesocosms for Oil Spill Simulation. In Handbook of Hydrocarbon and Lipid Microbiology; Timmis, K.N., Ed.; Springer: Berlin/Heidelberg, Germany, 2010; pp. 3513–3521. [Google Scholar] [CrossRef]
- Wozniak, A.S.; Prem, P.M.; Obeid, W.; Waggoner, D.C.; Quigg, A.; Xu, C.; Santschi, P.H.; Schwehr, K.A.; Hatcher, P.G. Rapid Degradation of Oil in Mesocosm Simulations of Marine Oil Snow Events. Environ. Sci. Technol. 2019, 53, 3441–3450. [Google Scholar] [CrossRef] [PubMed]
- Yamada, M.; Takada, H.; Toyoda, K.; Yoshida, A.; Shibata, A.; Nomura, H.; Wada, M.; Nishimura, M.; Okamoto, K.; Ohwada, K. Study on the Fate of Petroleum-Derived Polycyclic Aromatic Hydrocarbons (PAHs) and the Effect of Chemical Dispersant Using an Enclosed Ecosystem, Mesocosm. Mar. Pollut. Bull. 2003, 47, 105–113. [Google Scholar] [CrossRef]
- Teira, E.; Lekunberri, I.; Gasol, J.M.; Nieto-Cid, M.; Álvarez-Salgado, X.A.; Figueiras, F.G. Dynamics of the Hydrocarbon-Degrading Cycloclasticus Bacteria during Mesocosm-Simulated Oil Spills. Environ. Microbiol. 2007, 9, 2551–2562. [Google Scholar] [CrossRef] [PubMed]
- Cappello, S.; Denaro, R.; Genovese, M.; Giuliano, L.; Yakimov, M.M. Predominant Growth of Alcanivorax during Experiments on “Oil Spill Bioremediation” in Mesocosms. Microbiol. Res. 2007, 162, 185–190. [Google Scholar] [CrossRef] [PubMed]
- Gertler, C.; Gerdts, G.; Timmis, K.N.; Golyshin, P.N. Microbial Consortia in Mesocosm Bioremediation Trial Using Oil Sorbents, Slow-Release Fertilizer and Bioaugmentation. FEMS Microbiol. Ecol. 2009, 69, 288–300. [Google Scholar] [CrossRef] [PubMed]
- Hassanshahian, M.; Emtiazi, G.; Caruso, G.; Cappello, S. Bioremediation (Bioaugmentation/Biostimulation) Trials of Oil Polluted Seawater: A Mesocosm Simulation Study. Mar. Environ. Res. 2014, 95, 28–38. [Google Scholar] [CrossRef] [PubMed]
- Doyle, S.M.; Whitaker, E.A.; De Pascuale, V.; Wade, T.L.; Knap, A.H.; Santschi, P.H.; Quigg, A.; Sylvan, J.B. Rapid Formation of Microbe-Oil Aggregates and Changes in Community Composition in Coastal Surface Water Following Exposure to Oil and the Dispersant Corexit. Front. Microbiol. 2018, 9, 689. [Google Scholar] [CrossRef] [Green Version]
- García-Cruz, N.U.; Sánchez-Avila, J.I.; Valdés-Lozano, D.; Gold-Bouchot, G.; Aguirre-Macedo, L. Biodegradation of Hexadecane Using Sediments from Rivers and Lagoons of the Southern Gulf of Mexico. Mar. Pollut. Bull. 2018, 128, 202–207. [Google Scholar] [CrossRef]
- Carrit, D.E.; Carpenter, J.H. Recommendation procedure for Winkler analyses of sea water for dissolved oxygen. J. Mar. Res. 1966, 24, 313–318. [Google Scholar]
- Bryan, J.R.; Riley, J.P.; Williams, P.J. A Winkler Procedure for Making Precise Measurements Studies. J. Exp. Mar. Biol. Ecol. 1976, 21, 191–197. [Google Scholar] [CrossRef]
- Williams, P.J.l.B.; Jenkinson, N.W. A Transportable Microprocessor-controlled Precise Winkler Titration Suitable for Field Station and Shipboard Use. Limnol. Oceanogr. 1982, 27, 576–584. [Google Scholar] [CrossRef]
- Strickland, J.D.H.; Parsons, T.R. A Practical Hand Book of Seawater Analysis, 2nd ed.; Fisheries Research Board of Canada Bulletin: Ottawa, ON, Canada, 1972; pp. 157–310. [Google Scholar]
- Lizárraga-Partida, M.L.; Izquierdo-Vicuña, F.B.; Wong-Chang, I. Marine Bacteria on the Campeche Bank Oil Field. Mar. Pollut. Bull. 1991, 22, 401–405. [Google Scholar] [CrossRef]
- Uribe-Flores, M.M.; Cerqueda-García, D.; Hernández-Nuñez, E.; Cadena, S.; García-Cruz, N.U.; Trejo-Hernández, M.R.; Aguirre-Macedo, M.L.; García-Maldonado, J.Q. Bacterial Succession and Co-Occurrence Patterns of an Enriched Marine Microbial Community during Light Crude Oil Degradation in a Batch Reactor. J. Appl. Microbiol. 2019, 127, 495–507. [Google Scholar] [CrossRef] [PubMed]
- Klindworth, A.; Pruesse, E.; Schweer, T.; Peplies, J.; Quast, C.; Horn, M.; Glöckner, F.O. Evaluation of General 16S Ribosomal RNA Gene PCR Primers for Classical and Next-Generation Sequencing-Based Diversity Studies. Nucleic Acids Res. 2013, 41, e1. [Google Scholar] [CrossRef] [PubMed]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Gonzalez-Peña, A.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME Allows Analysis of High-Throughput Community Sequencing Data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-Resolution Sample Inference from Illumina Amplicon Data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef]
- Callahan, B.J.; McMurdie, P.J.; Holmes, S.P. Exact Sequence Variants Should Replace Operational Taxonomic Units in Marker-Gene Data Analysis. ISME J. 2017, 11, 2639–2643. [Google Scholar] [CrossRef]
- Rognes, T.; Flouri, T.; Nichols, B.; Quince, C.; Mahé, F. VSEARCH: A Versatile Open Source Tool for Metagenomics. PeerJ 2016, 4, e2584. [Google Scholar] [CrossRef]
- Price, M.N.; Dehal, P.S.; Arkin, A.P. FastTree 2—Approximately Maximum-Likelihood Trees for Large Alignments. PLoS ONE 2010, 5, e9490. [Google Scholar] [CrossRef] [PubMed]
- McMurdie, P.J.; Holmes, S. Phyloseq: An R Package for Reproducible Interactive Analysis and Graphics of Microbiome Census Data. PLoS ONE 2013, 8, e61217. [Google Scholar] [CrossRef] [PubMed]
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; Stevens, M.H.H.; Wagner, H. Vegan: Community Ecology Package. R Package Version 2.0-10. 2013. Available online: http://CRAN.R-project.org/package=vegan (accessed on 1 July 2019).
- Wickham, H. Ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2009; p. 213. [Google Scholar]
- Lozupone, C.; Knight, R. UniFrac: A New Phylogenetic Method for Comparing Microbial Communities. Appl. Environ. Microbiol. 2005, 71, 8228–8235. [Google Scholar] [CrossRef] [PubMed]
- Lozupone, C.A.; Knight, R. Global Patterns in Bacterial Diversity. Proc. Natl. Acad. Sci. USA 2007, 104, 11436–11440. [Google Scholar] [CrossRef] [PubMed]
- Herrera-Silveira, J.A.; Medina-Gomez, I.; Colli, R. Trophic Status Based on Nutrient Concentration Scales and Primary Producers Community of Tropical Coastal Lagoons Influenced by Groundwater Discharges. Hydrobiologia 2002, 475, 91–98. [Google Scholar] [CrossRef]
- Herrera-Silveira, J.A.; Comin, F.A.; Aranda-Cirerol, N.; Troccoli, L.; Capurro, L. Coastal Water Quality Assessment in the Yucatan Peninsula: Management Implications. Ocean Coast. Manag. 2004, 47, 625–639. [Google Scholar] [CrossRef]
- Overmann, J.; Lepleux, C. Marine Bacteria and Archaea: Diversity, Adaptations, and Culturability. In the Marine Microbiome; Stal, L.J., Cretoiu, M.S., Eds.; Springer: Cham, Switzerland, 2016; pp. 21–55. [Google Scholar]
- Lanfranconi, M.P.; Bosch, R.; Nogales, B. Short-Term Changes in the Composition of Active Marine Bacterial Assemblages in Response to Diesel Oil Pollution. Microb. Biotechnol. 2010, 3, 607–621. [Google Scholar] [CrossRef] [PubMed]
- Vila, J.; Nieto, J.M.; Mertens, J.; Springael, D.; Grifoll, M. Microbial Community Structure of a Heavy Fuel Oil-Degrading Marine Consortium: Linking Microbial Dynamics with Polycyclic Aromatic Hydrocarbon Utilization. FEMS Microbiol. Ecol. 2010, 73, 349–362. [Google Scholar] [CrossRef]
- Morris, L.; O’Brien, A.; Natera, S.H.A.; Lutz, A.; Roessner, U.; Long, S.M. Structural and Functional Measures of Marine Microbial Communities: An Experiment to Assess Implications for Oil Spill Management. Mar. Pollut. Bull. 2018, 131, 525–529. [Google Scholar] [CrossRef]
- Gertler, C.; Näther, D.J.; Cappello, S.; Gerdts, G.; Quilliam, R.S.; Yakimov, M.M.; Golyshin, P.N. Composition and Dynamics of Biostimulated Indigenous Oil-Degrading Microbial Consortia from the Irish, North and Mediterranean Seas: A Mesocosm Study. FEMS Microbiol. Ecol. 2012, 81, 520–536. [Google Scholar] [CrossRef]
- Dellagnezze, B.M.; Vasconcellos, S.P.; Angelim, A.L.; Melo, V.M.M.; Santisi, S.; Cappello, S.; Oliveira, V.M. Bioaugmentation Strategy Employing a Microbial Consortium Immobilized in Chitosan Beads for Oil Degradation in Mesocosm Scale. Mar. Pollut. Bull. 2016, 107, 107–117. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-R, L.M.; Overholt, W.A.; Hagan, C.; Huettel, M.; Kostka, J.E.; Konstantinidis, K.T. Microbial Community Successional Patterns in Beach Sands Impacted by the Deepwater Horizon Oil Spill. ISME J. 2015, 9, 1928–1940. [Google Scholar] [CrossRef] [PubMed]
- Röling, W.F.M.; Milner, M.G.; Jones, D.M.; Lee, K.; Daniel, F.; Swannell, R.J.P.; Head, I.M. Robust Hydrocarbon Degradation and Dynamics of Bacterial Communities during Nutrient-Enhanced Oil Spill Bioremediation. Appl. Environ. Microbiol. 2002, 68, 5537–5548. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herzyk, A.; Fillinger, L.; Larentis, M.; Qiu, S.; Maloszewski, P.; Hünniger, M.; Schmidt, S.I.; Stumpp, C.; Marozava, S.; Knappett, P.S.K.; et al. Response and Recovery of a Pristine Groundwater Ecosystem Impacted by Toluene Contamination—A Meso-Scale Indoor Aquifer Experiment. J. Contam. Hydrol. 2017, 207, 17–30. [Google Scholar] [CrossRef]
- Coclet, C.; Garnier, C.; Durrieu, G.; Omanović, D.; D’Onofrio, S.; Le Poupon, C.; Mullot, J.U.; Briand, J.F.; Misson, B. Changes in Bacterioplankton Communities Resulting from Direct and Indirect Interactions with Trace Metal Gradients in an Urbanized Marine Coastal Area. Front. Microbiol. 2019, 10, 257. [Google Scholar] [CrossRef]
- Paissé, S.; Coulon, F.; Goñi-Urriza, M.; Peperzak, L.; McGenity, T.J.; Duran, R. Structure of Bacterial Communities along a Hydrocarbon Contamination Gradient in a Coastal Sediment. FEMS Microbiol. Ecol. 2008, 66, 295–305. [Google Scholar] [CrossRef]
- Torstensson, A.; Dinasquet, J.; Chierici, M.; Fransson, A.; Riemann, L.; Wulff, A. Physicochemical Control of Bacterial and Protist Community Composition and Diversity in Antarctic Sea Ice. Environ. Microbiol. 2015, 17, 3869–3881. [Google Scholar] [CrossRef]
- Hazen, T.C.; Prince, R.C.; Mahmoudi, N. Marine Oil Biodegradation. Environ. Sci. Technol. 2016, 50, 2121–2129. [Google Scholar] [CrossRef]
- Wammer, K.H.; Peters, C.A. Polycyclic Aromatic Hydrocarbon Biodegradation Rates: A Structure-Based Study. Environ. Sci. Technol. 2005, 39, 2571–2578. [Google Scholar] [CrossRef]
- Wang, B.; Lai, Q.; Cui, Z.; Tan, T.; Shao, Z. A Pyrene-Degrading Consortium from Deep-Sea Sediment of the West Pacific and Its Key Member Cycloclasticus Sp. P1. Environ. Microbiol. 2008, 10, 1948–1963. [Google Scholar] [CrossRef]
- Sun, L.; Chiu, M.H.; Xu, C.; Lin, P.; Schwehr, K.A.; Bacosa, H.; Kamalanathan, M.; Quigg, A.; Chin, W.C.; Santschi, P.H. The Effects of Sunlight on the Composition of Exopolymeric Substances and Subsequent Aggregate Formation During Oil Spills. Mar. Chem. 2018, 203, 49–54. [Google Scholar] [CrossRef]
- Cappello, S.; Caruso, G.; Zampino, D.; Monticelli, L.S.; Maimone, G.; Denaro, R.; Tripodo, B.; Troussellier, M.; Yakimov, M.; Giuliano, L. Microbial Community Dynamics during Assays of Harbour Oil Spill Bioremediation: A Microscale Simulation Study. J. Appl. Microbiol. 2007, 102, 184–194. [Google Scholar] [CrossRef] [PubMed]
- Brakstad, O.G.; Daling, P.S.; Faksness, L.G.; Almås, I.K.; Vang, S.H.; Syslak, L.; Leirvik, F. Depletion and Biodegradation of Hydrocarbons in Dispersions and Emulsions of the Macondo 252 Oil Generated in an Oil-on-Seawater Mesocosm Flume Basin. Mar. Pollut. Bull. 2014, 84, 125–134. [Google Scholar] [CrossRef] [PubMed]
- Gutierrez, T. Marine, Aerobic Hydrocarbon-Degrading Gammaproteobacteria: Overview. In Taxonomy, Genomics and Ecophysiology of Hydrocarbon-Degrading Microbes. Handbook of Hydrocarbon and Lipid Microbiology; Mcgenity, T.J., Ed.; Springer: Cham, Switzerland, 2017; pp. 1–10. [Google Scholar]
- Yakimov, M.M.; Timmis, K.N.; Golyshin, P.N. Obligate Oil-Degrading Marine Bacteria. Curr. Opin. Biotechnol. 2007, 18, 257–266. [Google Scholar] [CrossRef] [PubMed]
- Dombrowski, N.; Donaho, J.A.; Gutierrez, T.; Seitz, K.W.; Teske, A.P.; Baker, B.J. Reconstructing Metabolic Pathways of Hydrocarbon-Degrading Bacteria from the Deepwater Horizon Oil Spill. Nat. Microbiol. 2016, 1, 16057. [Google Scholar] [CrossRef] [PubMed]
- Buchan, A.; González, J.M.; Chua, M.J. Aerobic Hydrocarbon-Degrading Alphaproteobacteria: Rhodobacteraceae (Roseobacter). In Taxonomy, Genomics and Ecophysiology of Hydrocarbon-Degrading Microbes; Mcgenity, T.J., Ed.; Springer: Cham, Switzerland, 2019; pp. 1–13. [Google Scholar] [CrossRef]
- Guibert, L.M.; Loviso, C.L.; Borglin, S.; Jansson, J.K.; Dionisi, H.M.; Lozada, M. Diverse Bacterial Groups Contribute to the Alkane Degradation Potential of Chronically Polluted Subantarctic Coastal Sediments. Microb. Ecol. 2016, 71, 100–112. [Google Scholar] [CrossRef] [PubMed]
- Varjani, S.J. Microbial Degradation of Petroleum Hydrocarbons. Bioresour. Technol. 2017, 223, 277–286. [Google Scholar] [CrossRef]
- Rochman, F.F. Aerobic Hydrocarbon-Degrading Microbial Communities in Oilsands Tailings Ponds. Ph.D. Thesis, University of Calgary, Calgary, AB, Canada, 2016. [Google Scholar]
- Chikere, C.B.; Mordi, I.J.; Chikere, B.O.; Selvarajan, R.; Ashafa, T.O.; Obieze, C.C. Comparative Metagenomics and Functional Profiling of Crude Oil-Polluted Soils in Bodo West Community, Ogoni, with Other Sites of Varying Pollution History. Ann. Microbiol. 2019, 69, 495–513. [Google Scholar] [CrossRef]
- Woźniak-Karczewska, M.; Lisiecki, P.; Białas, W.; Owsianiak, M.; Piotrowska-Cyplik, A.; Wolko, Ł.; Ławniczak, Ł.; Heipieper, H.J.; Gutierrez, T.; Chrzanowski, Ł. Effect of Bioaugmentation on Long-Term Biodegradation of Diesel/Biodiesel Blends in Soil Microcosms. Sci. Total Environ. 2019, 671, 948–958. [Google Scholar] [CrossRef]
- Hwang, C.Y.; Bae, G.D.; Yih, W.; Cho, B.C. Marivita Cryptomonadis Gen. Nov., Sp. Nov. and Marivita Litorea Sp. Nov., of the Family Rhodobacteraceae, Isolated from Marine Habitats. Int. J. Syst. Evolut. Microbiol. 2009, 59, 1568–1575. [Google Scholar] [CrossRef]
- Daniel, R.; Simon, M.; Wemheuer, B. Editorial: Molecular Ecology and Genetic Diversity of the Roseobacter Clade. Front. Microbiol. 2018, 9, 1185. [Google Scholar] [CrossRef] [PubMed]
- Lofthus, S.; Netzer, R.; Lewin, A.S.; Heggeset, T.M.B.; Haugen, T.; Brakstad, O.G. Biodegradation of N-Alkanes on Oil–Seawater Interfaces at Different Temperatures and Microbial Communities Associated with the Degradation. Biodegradation 2018, 29, 141–157. [Google Scholar] [CrossRef] [PubMed]
- Teramoto, M.; Queck, S.Y.; Ohnishi, K. Specialized Hydrocarbonoclastic Bacteria Prevailing in Seawater around a Port in the Strait of Malacca. PLoS ONE 2013, 8, e66594. [Google Scholar] [CrossRef]
- Teramoto, M.; Suzuki, M.; Okazaki, F.; Hatmanti, A.; Harayama, S. Oceanobacter-Related Bacteria Are Important for the Degradation of Petroleum Aliphatic Hydrocarbons in the Tropical Marine Environment. Microbiology 2009, 155, 3362–3370. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | Reference | 0 | 2 | 4 | 6 | 8 | 10 | 14 |
|---|---|---|---|---|---|---|---|---|
| Total_C (%) | 0.00 ±0.008 | 0.01 ±0.01 | 0.00 ±0.02 | 0.00 ±0.02 | 0.02 ±0.01 | 0.01 ±0.003 | 0.01 ±0.04 | 0.04 ±0.02 |
| Total_N (%) | 0.39 ±0.13 | 0.33 ±0.03 | 0.39 ±0.01 | 0.48 ±0.01 | 0.41 ±0.06 | 0.51 ±0.07 | 0.64 ±0.11 | 1.55 ±0.08 |
| Temperature (°C) | 28.3 | 27.00 | 26.2 | 27.10 | 26.30 | 26.60 | 27.40 | 27.80 |
| O2 (mg L−1) | 1.90 ±0.06 | 3.84 ±0.22 | 2.88 ±0.17 | 3.94 ±0.77 | 3.51 ±0.26 | 3.85 ±0.67 | 4.19 ±0.22 | 4.54 ±0.14 |
| Salinity | 36.20 | 36.13 | 35.7 | 35.93 | 35.90 | 35.53 | 36.07 | 36.40 |
| PO4 (µmol L−1) | 0.89 ±0.07 | 0.61 ±0.02 | 0.75 ±0.02 | 1.18 ±0.09 | 0.91 ±0.07 | 1.20 ±0.05 | 1.09 ±0.01 | 1.04 ±0.14 |
| NO2 (µmol L−1) | 0.08 ±0.01 | 0.12 ±0.002 | 0.10 ±0.009 | 0.09 ±0.09 | 0.10 ±0.006 | 0.11 ±0.01 | 0.07 ±0.02 | 0.13 ±0.001 |
| NO3 (µmol L−1) | 4.34 ±0.14 | 0.77 ±0.03 | 0.43 ±0.10 | 0.38 ±0.06 | 0.41 ±0.17 | 0.23 ±0.17 | 0.17 ±0.03 | 0.49 ±0.04 |
| SiO4 (µmol L−1) | 3.26 ±0.23 | 0.09 ±0.02 | 0.38 ±0.11 | 0.16 ±0.16 | 0.25 ±0.59 | 0.12 ±0.01 | 0.60 ±0.28 | 0.25 ±0.11 |
| NH4 (µmol L−1) | 1.04 ±0.04 | 1.33 ±0.01 | 1.42 ±0.01 | 0.99 ±0.01 | 0.83 ±0.02 | 0.69 ±0.005 | 0.76 ±0.06 | 1.16 ±0.51 |
| Day | Input Reads | Clean Reads | Observed ASVs | Shannon |
|---|---|---|---|---|
| R | 150,285 | 45,912 | 263 | 5.18 |
| 0 | 133,342 | 39,671 | 236 | 5.02 |
| 2 | 115,598 | 32,610 | 178 | 4.66 |
| 4 | 113,562 | 34,066 | 196 | 4.74 |
| 6 | 126,258 | 37,726 | 231 | 4.94 |
| 8 | 135,164 | 39,719 | 234 | 5.08 |
| 10 | 145,363 | 42,706 | 255 | 5.06 |
| 14 | 293,188 | 64,015 | 317 | 5.08 |
| Variable | R2 | Pr(>F) |
|---|---|---|
| Total Nitrogen | 0.22 | 0.030 |
| PAHs fraction | 0.25 | 0.048 |
| Naphthalene | 0.32 | 0.003 |
| 1-metilnaphtalene | 0.025 | 0.041 |
| 2-metilnaphtalene | 0.33 | 0.002 |
| Fluoranthene | 0.29 | 0.013 |
| Pyrene | 0.28 | 0.026 |
| Chrysene | 0.33 | 0.022 |
| Benzo(b)fluoranthene | 0.29 | 0.013 |
| Benzo(e)pyrene | 0.30 | 0.025 |
| Benzo(a)pyrene | 0.35 | 0.005 |
| Benzo(ghi)perylene | 0.30 | 0.020 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Valencia-Agami, S.S.; Cerqueda-García, D.; Putzeys, S.; Uribe-Flores, M.M.; García-Cruz, N.U.; Pech, D.; Herrera-Silveira, J.; Aguirre-Macedo, M.L.; García-Maldonado, J.Q. Changes in the Bacterioplankton Community Structure from Southern Gulf of Mexico During a Simulated Crude Oil Spill at Mesocosm Scale. Microorganisms 2019, 7, 441. https://doi.org/10.3390/microorganisms7100441
Valencia-Agami SS, Cerqueda-García D, Putzeys S, Uribe-Flores MM, García-Cruz NU, Pech D, Herrera-Silveira J, Aguirre-Macedo ML, García-Maldonado JQ. Changes in the Bacterioplankton Community Structure from Southern Gulf of Mexico During a Simulated Crude Oil Spill at Mesocosm Scale. Microorganisms. 2019; 7(10):441. https://doi.org/10.3390/microorganisms7100441
Chicago/Turabian StyleValencia-Agami, Sonia S., Daniel Cerqueda-García, Sébastien Putzeys, María Magdalena Uribe-Flores, Norberto Ulises García-Cruz, Daniel Pech, Jorge Herrera-Silveira, M. Leopoldina Aguirre-Macedo, and José Q. García-Maldonado. 2019. "Changes in the Bacterioplankton Community Structure from Southern Gulf of Mexico During a Simulated Crude Oil Spill at Mesocosm Scale" Microorganisms 7, no. 10: 441. https://doi.org/10.3390/microorganisms7100441
APA StyleValencia-Agami, S. S., Cerqueda-García, D., Putzeys, S., Uribe-Flores, M. M., García-Cruz, N. U., Pech, D., Herrera-Silveira, J., Aguirre-Macedo, M. L., & García-Maldonado, J. Q. (2019). Changes in the Bacterioplankton Community Structure from Southern Gulf of Mexico During a Simulated Crude Oil Spill at Mesocosm Scale. Microorganisms, 7(10), 441. https://doi.org/10.3390/microorganisms7100441