Characteristics of Carbapenem-Resistant and Colistin-Resistant Escherichia coli Co-Producing NDM-1 and MCR-1 from Pig Farms in China

 , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Antimicrobial Susceptibility Testing
2.2. Detection of the Carbapenem-Resistant and Colistin-Resistant Genes
2.3. DNA Extraction and Whole Genome Sequencing and Bioinformatic Analysis
2.4. Plasmid Conjugation
3. Results
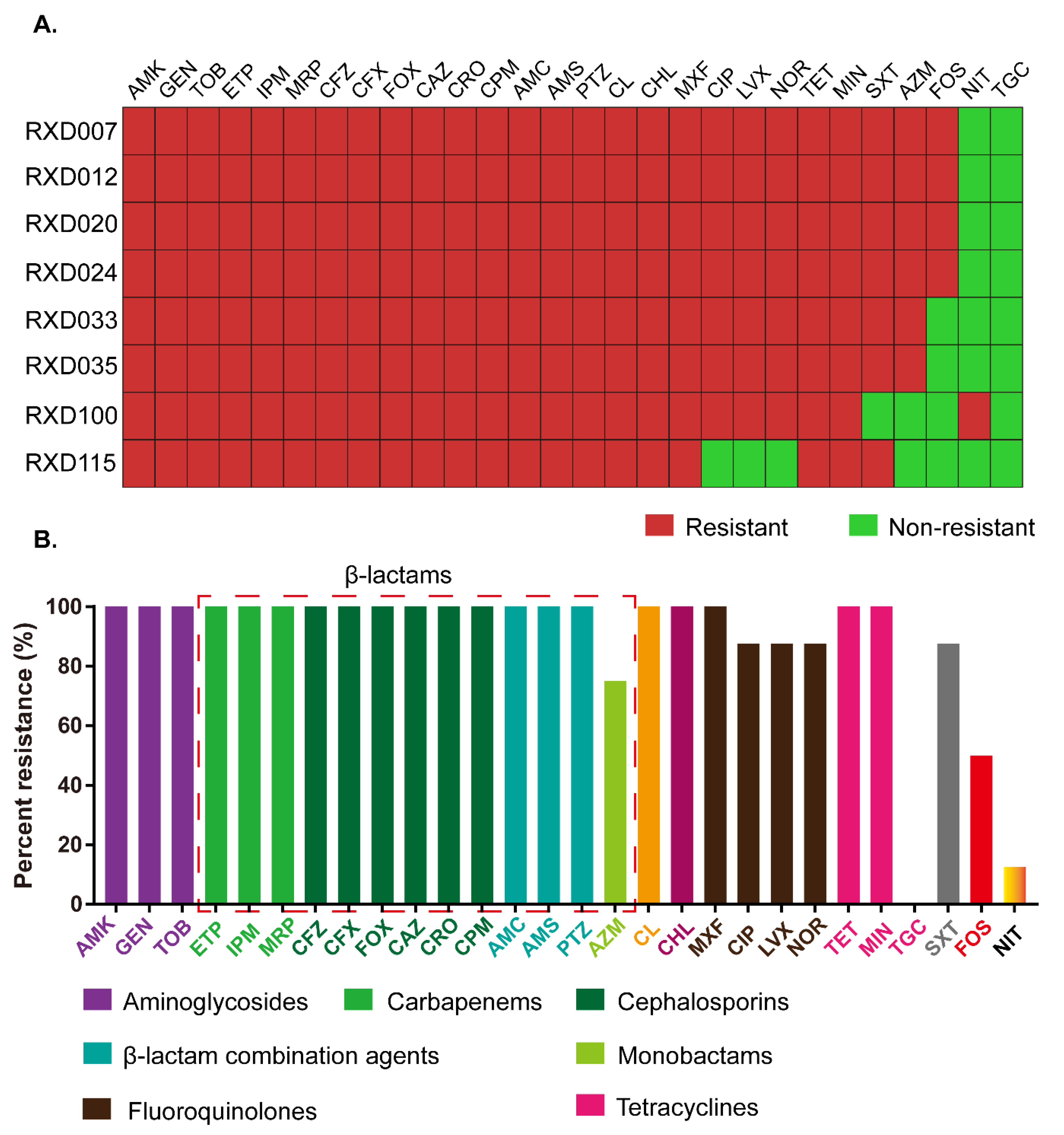
3.1. Antimicrobial Susceptibility Profile of the Eight E. coli Isolates
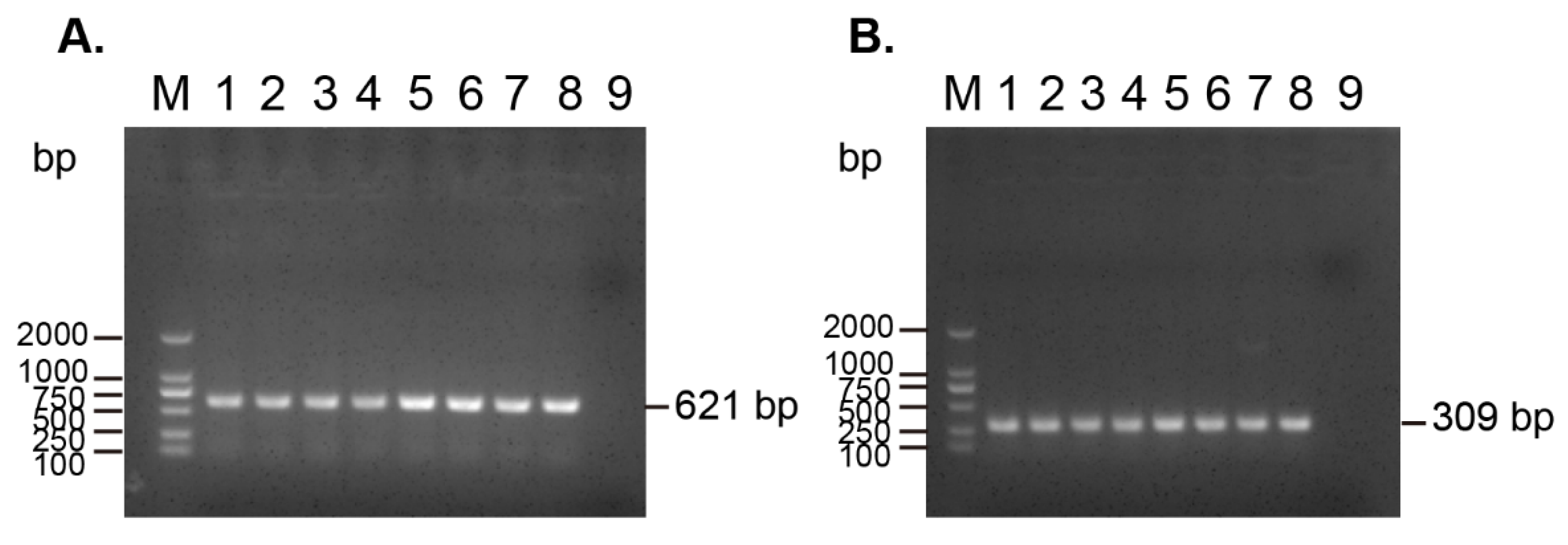
3.2. Detection of the Carbapenem-Resistant Gene blaNDM-1 and Colistin-Resistant Gene mcr-1
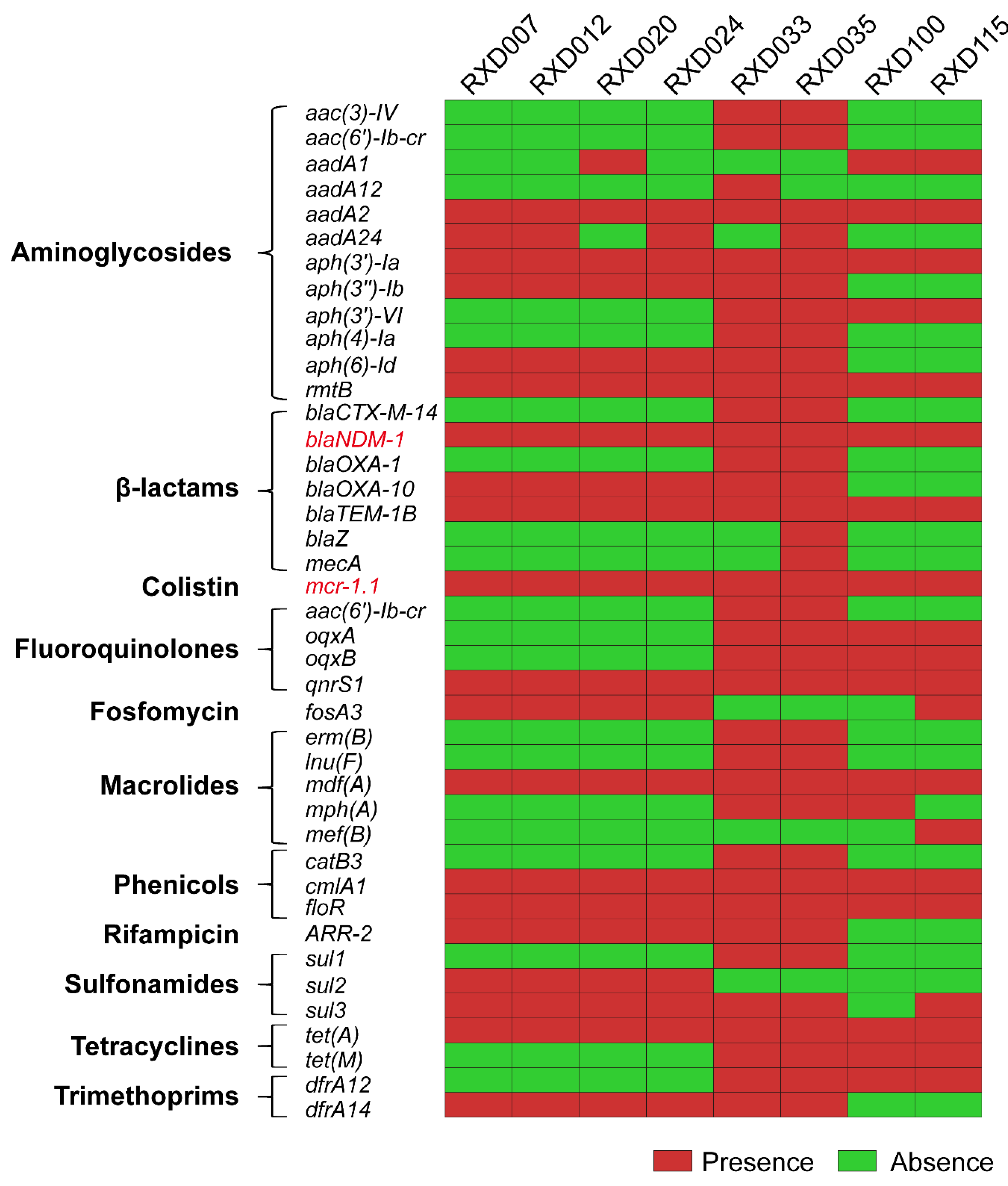
3.3. Acquired Antibiotic Resistance Genes in the Eight Carbapenem-Resistant and Colistin-Resistant E. coli
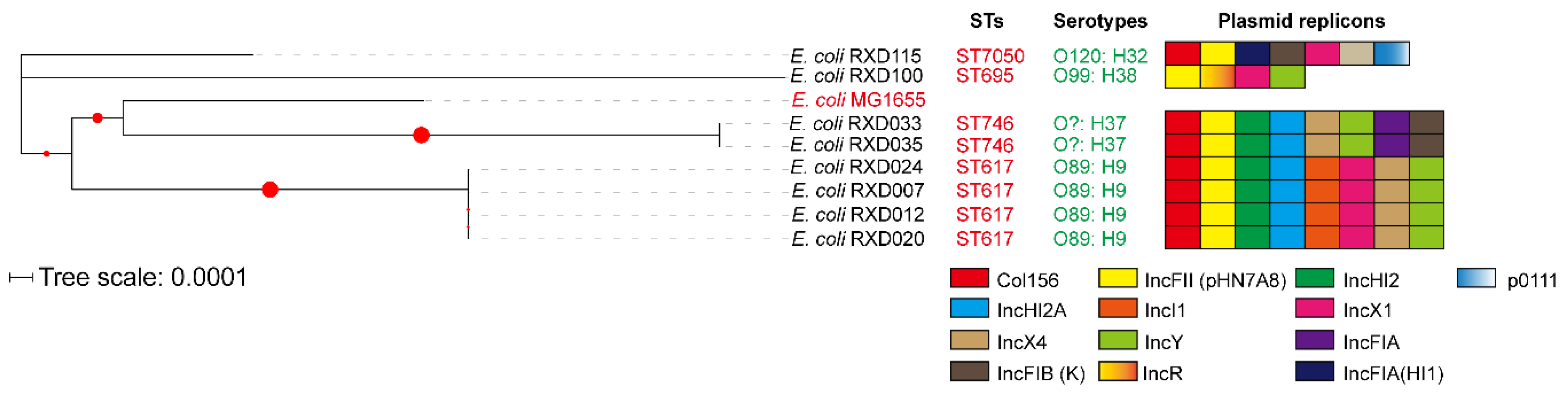
3.4. Genetic Characteristics
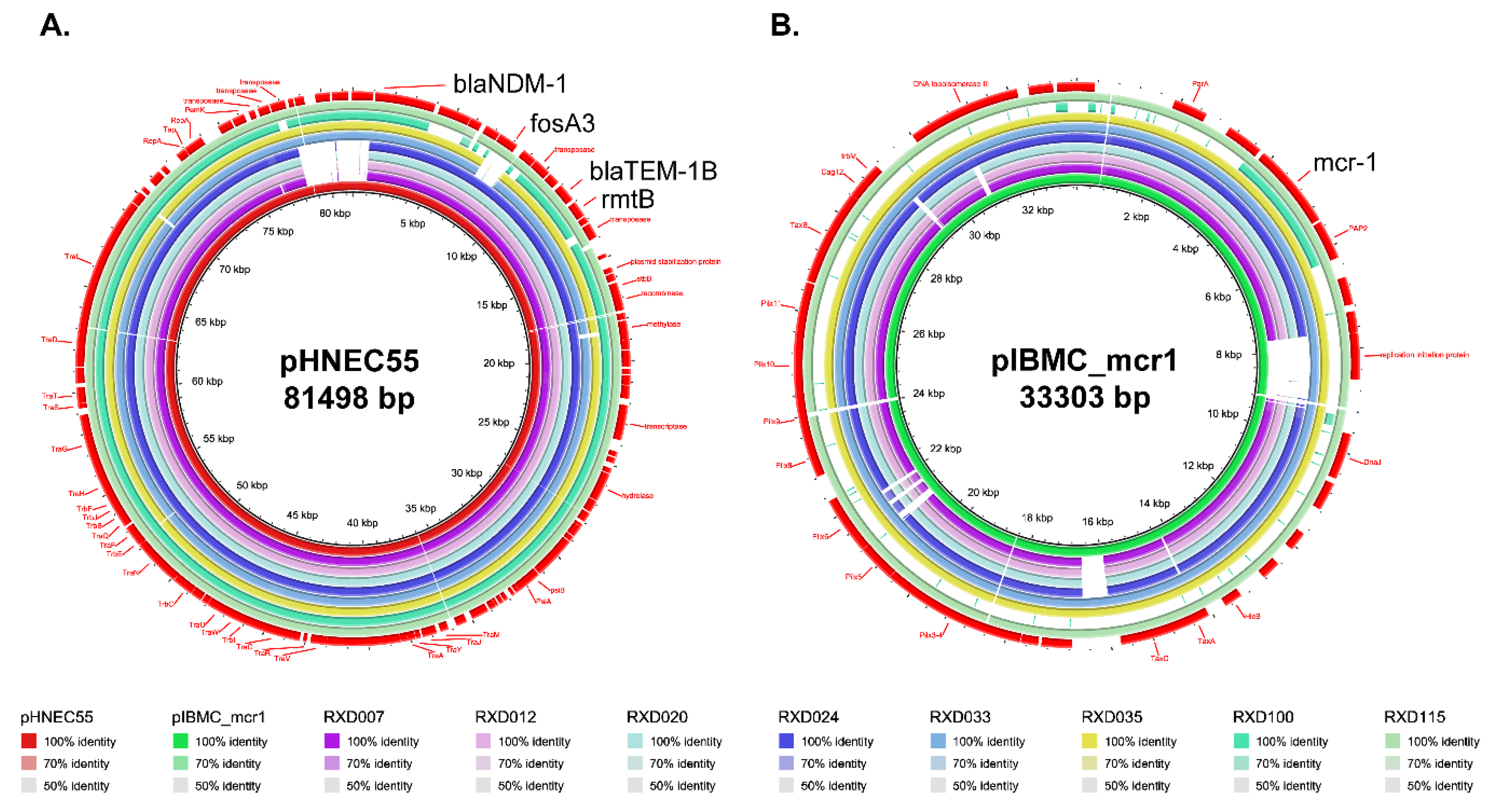
3.5. Plasmid Determination
3.6. Plasmid Conjugation
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Bielaszewska, M.; Mellmann, A.; Zhang, W.; Kock, R.; Fruth, A.; Bauwens, A.; Peters, G.; Karch, H. Characterisation of the Escherichia coli strain associated with an outbreak of haemolytic uraemic syndrome in Germany, 2011: A microbiological study. Lancet Infect. Dis. 2011, 11, 671–676. [Google Scholar] [CrossRef]
- Vidovic, S.; Korber, D.R. Escherichia coli O157: Insights into the adaptive stress physiology and the influence of stressors on epidemiology and ecology of this human pathogen. Crit. Rev. Microbiol. 2016, 42, 83–93. [Google Scholar] [CrossRef] [PubMed]
- Chekabab, S.M.; Paquin-Veillette, J.; Dozois, C.M.; Harel, J. The ecological habitat and transmission of Escherichia coli O157:H7. FEMS Microbiol. Lett. 2013, 341, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Navarro-Garcia, F. Escherichia coli O104:H4 Pathogenesis: An Enteroaggregative E. coli/Shiga Toxin-Producing E. coli Explosive Cocktail of High Virulence. Microbiol. Spectr. 2014, 2. [Google Scholar] [CrossRef] [PubMed]
- Poirel, L.; Madec, J.Y.; Lupo, A.; Schink, A.K.; Kieffer, N.; Nordmann, P.; Schwarz, S. Antimicrobial Resistance in Escherichia coli. Microbiol. Spectr. 2018, 6. [Google Scholar] [CrossRef] [PubMed]
- Wailan, A.M.; Paterson, D.L. The spread and acquisition of NDM-1: A multifactorial problem. Expert Rev. Anti Infect. Ther. 2014, 12, 91–115. [Google Scholar] [CrossRef] [PubMed]
- Paterson, D.L.; Harris, P.N. Colistin resistance: A major breach in our last line of defence. Lancet Infect. Dis. 2016, 16, 132–133. [Google Scholar] [CrossRef]
- Sun, J.; Yang, R.S.; Zhang, Q.; Feng, Y.; Fang, L.X.; Xia, J.; Li, L.; Lv, X.Y.; Duan, J.H.; Liao, X.P.; et al. Co-transfer of blaNDM-5 and mcr-1 by an IncX3-X4 hybrid plasmid in Escherichia coli. Nat. Microbiol. 2016, 1, 16176. [Google Scholar] [CrossRef]
- Liu, Y.Y.; Wang, Y.; Walsh, T.R.; Yi, L.X.; Zhang, R.; Spencer, J.; Doi, Y.; Tian, G.; Dong, B.; Huang, X.; et al. Emergence of plasmid-mediated colistin resistance mechanism MCR-1 in animals and human beings in China: A microbiological and molecular biological study. Lancet Infect. Dis. 2016, 16, 161–168. [Google Scholar] [CrossRef]
- Lai, C.C.; Chuang, Y.C.; Chen, C.C.; Tang, H.J. Coexistence of MCR-1 and NDM-9 in a clinical carbapenem-resistant Escherichia coli isolate. Int. J. Antimicrob. Agents 2017, 49, 517–518. [Google Scholar] [CrossRef]
- Delgado-Blas, J.F.; Ovejero, C.M.; Abadia-Patino, L.; Gonzalez-Zorn, B. Coexistence of mcr-1 and blaNDM-1 in Escherichia coli from Venezuela. Antimicrob. Agents Chemother. 2016, 60, 6356–6358. [Google Scholar] [CrossRef] [PubMed]
- Yang, R.S.; Feng, Y.; Lv, X.Y.; Duan, J.H.; Chen, J.; Fang, L.X.; Xia, J.; Liao, X.P.; Sun, J.; Liu, Y.H. Emergence of NDM-5- and MCR-1-Producing Escherichia coli Clones ST648 and ST156 from a Single Muscovy Duck (Cairina moschata). Antimicrob. Agents Chemother. 2016, 60, 6899–6902. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, R.; Liu, Y.; Zhang, Q.; Jin, L.; Wang, Q.; Zhang, Y.; Wang, X.; Hu, M.; Li, L.; Qi, J.; et al. The prevalence of colistin resistance in Escherichia coli and Klebsiella pneumoniae isolated from food animals in China: Coexistence of mcr-1 and blaNDM with low fitness cost. Int. J. Antimicrob. Agents 2018, 51, 739–744. [Google Scholar] [CrossRef] [PubMed]
- Yao, X.; Doi, Y.; Zeng, L.; Lv, L.; Liu, J.H. Carbapenem-resistant and colistin-resistant Escherichia coli co-producing NDM-9 and MCR-1. Lancet Infect. Dis. 2016, 16, 288–289. [Google Scholar] [CrossRef] [Green Version]
- Wirth, T.; Falush, D.; Lan, R.; Colles, F.; Mensa, P.; Wieler, L.H.; Karch, H.; Reeves, P.R.; Maiden, M.C.; Ochman, H.; et al. Sex and virulence in Escherichia coli: An evolutionary perspective. Mol. Microbiol. 2006, 60, 1136–1151. [Google Scholar] [CrossRef]
- El-Shazly, S.; Dashti, A.; Vali, L.; Bolaris, M.; Ibrahim, A.S. Molecular epidemiology and characterization of multiple drug-resistant (MDR) clinical isolates of Acinetobacter baumannii. Int. J. Infect. Dis. 2015, 41, 42–49. [Google Scholar] [CrossRef] [Green Version]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A new genome assembly algorithm and its applications to single-cell sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef]
- Joensen, K.G.; Tetzschner, A.M.; Iguchi, A.; Aarestrup, F.M.; Scheutz, F. Rapid and Easy in Silico Serotyping of Escherichia coli Isolates by Use of Whole-Genome Sequencing Data. J. Clin. Microbiol. 2015, 53, 2410–2426. [Google Scholar] [CrossRef]
- Larsen, M.V.; Cosentino, S.; Rasmussen, S.; Friis, C.; Hasman, H.; Marvig, R.L.; Jelsbak, L.; Sicheritz-Ponten, T.; Ussery, D.W.; Aarestrup, F.M.; et al. Multilocus sequence typing of total-genome-sequenced bacteria. J. Clin. Microbiol. 2012, 50, 1355–1361. [Google Scholar] [CrossRef]
- Zankari, E.; Hasman, H.; Cosentino, S.; Vestergaard, M.; Rasmussen, S.; Lund, O.; Aarestrup, F.M.; Larsen, M.V. Identification of acquired antimicrobial resistance genes. J. Antimicrob. Chemother. 2012, 67, 2640–2644. [Google Scholar] [CrossRef]
- Carattoli, A.; Zankari, E.; Garcia-Fernandez, A.; Voldby Larsen, M.; Lund, O.; Villa, L.; Moller Aarestrup, F.; Hasman, H. In silico detection and typing of plasmids using PlasmidFinder and plasmid multilocus sequence typing. Antimicrob. Agents Chemother. 2014, 58, 3895–3903. [Google Scholar] [CrossRef] [PubMed]
- Goris, J.; Konstantinidis, K.T.; Klappenbach, J.A.; Coenye, T.; Vandamme, P.; Tiedje, J.M. DNA-DNA hybridization values and their relationship to whole-genome sequence similarities. Int. J. Syst. Evol. Microbiol. 2007, 57, 81–91. [Google Scholar] [CrossRef] [PubMed]
- Alikhan, N.F.; Petty, N.K.; Ben Zakour, N.L.; Beatson, S.A. BLAST Ring Image Generator (BRIG): Simple prokaryote genome comparisons. BMC Genom. 2011, 12, 402. [Google Scholar] [CrossRef] [PubMed]
- Sullivan, M.J.; Petty, N.K.; Beatson, S.A. Easyfig: A genome comparison visualizer. Bioinformatics 2011, 27, 1009–1010. [Google Scholar] [CrossRef]
- Tamura, K.; Nei, M. Estimation of the number of nucleotide substitutions in the control region of mitochondrial DNA in humans and chimpanzees. Mol. Biol. Evol. 1993, 10, 512–526. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Rogolsky, M.; Beall, B.W.; Wiley, B.B. Transfer of the plasmid for exfoliative toxin B synthesis in mixed cultures on nitrocellulose membranes. Infect. Immun. 1986, 54, 265–268. [Google Scholar] [Green Version]
- Shaheen, B.W.; Nayak, R.; Boothe, D.M. Emergence of a New Delhi metallo-beta-lactamase (NDM-1)-encoding gene in clinical Escherichia coli isolates recovered from companion animals in the United States. Antimicrob. Agents Chemother. 2013, 57, 2902–2903. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, R.; Li, J.; Wu, Z.; Yin, W.; Schwarz, S.; Tyrrell, J.M.; Zheng, Y.; Wang, S.; Shen, Z.; et al. Comprehensive resistome analysis reveals the prevalence of NDM and MCR-1 in Chinese poultry production. Nat. Microbiol. 2017, 2, 16260. [Google Scholar] [CrossRef]
- Li, J.; Nation, R.L.; Turnidge, J.D.; Milne, R.W.; Coulthard, K.; Rayner, C.R.; Paterson, D.L. Colistin: The re-emerging antibiotic for multidrug-resistant Gram-negative bacterial infections. Lancet Infect. Dis. 2006, 6, 589–601. [Google Scholar] [CrossRef]
- Moellering, R.C., Jr. NDM-1—A cause for worldwide concern. N. Engl. J. Med. 2010, 363, 2377–2379. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Chen, J.; Wang, L.; Wu, Z. Whole genome sequence of an MCR-1-carrying, extended-spectrum beta-lactamase (ESBL)-producing Escherichia coli ST746 isolate recovered from a community-acquired urinary tract infection. J. Glob. Antimicrob. Resist. 2018, 13, 171–173. [Google Scholar] [CrossRef] [PubMed]
- Cai, Y.; Chen, C.; Zhao, M.; Yu, X.; Lan, K.; Liao, K.; Guo, P.; Zhang, W.; Ma, X.; He, Y.; et al. High Prevalence of Metallo-beta-Lactamase-Producing Enterobacter cloacae from Three Tertiary Hospitals in China. Front. Microbiol. 2019, 10, 1610. [Google Scholar] [CrossRef] [PubMed]
- Paskova, V.; Medvecky, M.; Skalova, A.; Chudejova, K.; Bitar, I.; Jakubu, V.; Bergerova, T.; Zemlickova, H.; Papagiannitsis, C.C.; Hrabak, J. Characterization of NDM-Encoding Plasmids from Enterobacteriaceae Recovered From Czech Hospitals. Front. Microbiol. 2018, 9, 1549. [Google Scholar] [CrossRef]
- Pedersen, T.; Sekyere, J.O.; Govinden, U.; Moodley, K.; Sivertsen, A.; Samuelsen, O.; Essack, S.Y.; Sundsfjord, A. Spread of Plasmid-Encoded NDM-1 and GES-5 Carbapenemases among Extensively Drug-Resistant and Pandrug-Resistant Clinical Enterobacteriaceae in Durban, South Africa. Antimicrob. Agents Chemother. 2018, 62. [Google Scholar] [CrossRef] [Green Version]
- Baraniak, A.; Machulska, M.; Zabicka, D.; Literacka, E.; Izdebski, R.; Urbanowicz, P.; Bojarska, K.; Herda, M.; Kozinska, A.; Hryniewicz, W.; et al. Towards endemicity: Large-scale expansion of the NDM-1-producing Klebsiella pneumoniae ST11 lineage in Poland, 2015–2016. J. Antimicrob. Chemother. 2019. [Google Scholar] [CrossRef]
- Yang, B.; Feng, Y.; McNally, A.; Zong, Z. Occurrence of Enterobacter hormaechei carrying blaNDM-1 and blaKPC-2 in China. Diagn. Microbiol. Infect. Dis. 2018, 90, 139–142. [Google Scholar] [CrossRef]
- Manageiro, V.; Clemente, L.; Romao, R.; Silva, C.; Vieira, L.; Ferreira, E.; Canica, M. IncX4 Plasmid Carrying the New mcr-1.9 Gene Variant in a CTX-M-8-Producing Escherichia coli Isolate Recovered from Swine. Front. Microbiol. 2019, 10, 367. [Google Scholar] [CrossRef]
- Bai, F.; Li, X.; Niu, B.; Zhang, Z.; Malakar, P.K.; Liu, H.; Pan, Y.; Zhao, Y. A mcr-1-Carrying Conjugative IncX4 Plasmid in Colistin-Resistant Escherichia coli ST278 Strain Isolated from Dairy Cow Feces in Shanghai, China. Front. Microbiol. 2018, 9, 2833. [Google Scholar] [CrossRef]
- Dalmolin, T.V.; Martins, A.F.; Zavascki, A.P.; de Lima-Morales, D.; Barth, A.L. Acquisition of the mcr-1 gene by a high-risk clone of KPC-2-producing Klebsiella pneumoniae ST437/CC258, Brazil. Diagn. Microbiol. Infect. Dis. 2018, 90, 132–133. [Google Scholar] [CrossRef]
- Xu, L.; Wang, P.; Cheng, J.; Qin, S.; Xie, W. Characterization of a novel bla NDM-5-harboring IncFII plasmid and an mcr-1-bearing IncI2 plasmid in a single Escherichia coli ST167 clinical isolate. Infect. Drug Resist. 2019, 12, 511–519. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Feng, Y.; Liu, F.; Jiang, H.; Qu, Z.; Lei, M.; Wang, J.; Zhang, B.; Hu, Y.; Ding, J.; et al. A Phage-Like IncY Plasmid Carrying the mcr-1 Gene in Escherichia coli from a Pig Farm in China. Antimicrob. Agents Chemother. 2017, 61, e02035-16. [Google Scholar] [CrossRef] [PubMed]
- Lin, D.; Xie, M.; Li, R.; Chen, K.; Chan, E.W.; Chen, S. IncFII Conjugative Plasmid-Mediated Transmission of blaNDM-1 Elements among Animal-Borne Escherichia coli Strains. Antimicrob. Agents Chemother. 2017, 61. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antibiotics tested | Transconjugants selected by rifampin plus imipenem plus colistin | E. coli C600 | ||||||
|---|---|---|---|---|---|---|---|---|
| Minimum inhibitory concentration (μg/ml) | ||||||||
| Cir007 1 | Cir012 | Cir020 | Cir033 | Cir035 | Cir100 | Cir115 | ||
| Amikacin | > 32 (R1) | > 32 (R) | > 32 (R) | > 32 (R) | > 32 (R) | > 32 (R) | > 32 (R) | ≤ 8 (S) |
| Gentamicin | > 8 (R) | > 8 (R) | > 8 (R) | > 8 (R) | > 8 (R) | > 8 (R) | > 8 (R) | ≤ 2 (S) |
| Tobramycin | > 8 (R) | > 8 (R) | > 8 (R) | > 8 (R) | > 8 (R) | > 8 (R) | > 8 (R) | ≤ 2 (S) |
| Ertapenem | > 2 (R) | > 2 (R) | > 2 (R) | > 2 (R) | > 2 (R) | > 2 (R) | 2 (R) | ≤ 0.25 (S) |
| Imipenem | > 8 (R) | > 8 (R) | > 8 (R) | > 8 (R) | > 8 (R) | 1 (S) | 8 (R) | 0.5 (S) |
| Meropenem | > 8 (R) | > 8 (R) | > 8 (R) | > 8 (R) | > 8 (R) | 1 (S) | 4 (R) | ≤ 0.13 (S) |
| Cefazolin | > 16 (R) | > 16 (R) | > 16 (R) | > 16 (R) | > 16 (R) | > 16 (R) | > 16 (R) | 4 (S) |
| Cefuroxime | > 16 (R) | > 16 (R) | > 16 (R) | > 16 (R) | > 16 (R) | > 16 (R) | > 16 (R) | 16 (R) |
| Cefoxitin | > 16 (R) | > 16 (R) | > 16 (R) | > 16 (R) | > 16 (R) | > 16 (R) | > 16 (R) | 8 (S) |
| Ceftazidime | > 32 (R) | > 32 (R) | > 32 (R) | > 32 (R) | > 32 (R) | > 32 (R) | > 32 (R) | ≤ 1 (S) |
| Ceftriaxone | > 32 (R) | > 32 (R) | > 32 (R) | > 32 (R) | > 32 (R) | > 32 (R) | > 32 (R) | ≤ 1 (S) |
| Cefepime | > 16 (R) | > 16 (R) | > 16 (R) | 16 (R) | > 16 (R) | 16 (R) | 8 (R) | ≤ 1 (S) |
| Amoxicillin/clavulanate | > 32/16 (R) | > 32/16 (R) | > 32/16 (R) | > 32/16 (R) | > 32/16 (R) | 32/16 (R) | 32/16 (R) | ≤ 8/4 (S) |
| Ampicillin/sulbactam | >16/8 (R) | >16/8 (R) | >16/8 (R) | >16/8 (R) | > 16/8 (R) | > 16/8 (R) | >16/8 (R) | 8/4 (S) |
| Piperacillin/tazobactam | >64/4 (R) | >64/4 (R) | >64/4 (R) | >64/4 (R) | >64/4 (R) | >64/4 (R) | >64/4 (R) | ≤ 4/4 (S) |
| Colistin | > 4 (R) | > 4 (R) | 4 (R) | 4 (R) | 4 (R) | > 4 (R) | > 4 (R) | ≤ 1 (S) |
| Chloramphenicol | > 16 (R) | > 16 (R) | > 16 (R) | ≤ 4 (S) | ≤ 4 (S) | > 16 (R) | ≤ 4 (S) | ≤ 4 (S) |
| Moxifloxacin | > 2 (R) | > 2 (R) | > 2 (R) | ≤ 0.5 (S) | ≤ 0.5 (S) | > 2 (R) | ≤ 0.5 (S) | ≤ 0.5 (S) |
| Ciprofloxacin | > 4 (R) | > 4 (R) | > 4 (R) | ≤ 0.5 (S) | ≤ 0.5 (S) | 2 (I) | ≤ 0.5 (S) | ≤ 0.5 (S) |
| Levofloxacin | > 8 (R) | > 8 (R) | > 8 (R) | ≤ 1 (S) | ≤ 1 (S) | 2 (S) | ≤ 1 (S) | ≤ 1 (S) |
| Norfloxacin | > 8 (R) | > 8 (R) | > 8 (R) | ≤ 2 (S) | ≤ 2 (S) | 4 (S) | ≤ 2 (S) | ≤ 2 (S) |
| Tetracycline | > 8 (R) | > 8 (R) | > 8 (R) | ≤ 2 (S) | ≤ 2 (S) | > 8 (R) | ≤ 2 (S) | ≤ 2 (S) |
| Minocycline | 4 (S) | 4 (S) | 4 (S) | ≤ 1 (S) | 2 (S) | 16 (R) | 2 (S) | ≤ 1 (S) |
| Trimethoprim/sulfamethoxazole | > 4/76 (R) | > 4/76 (R) | > 4/76 (R) | ≤1/19 (S) | ≤ 1/19 (S) | ≤ 1/19 (S) | ≤1/19 (S) | ≤ 1/19 (S) |
| Aztreonam | ≤ 2 (S) | ≤ 2 (S) | ≤ 2 (S) | ≤ 2 (S) | ≤ 2 (S) | ≤ 2 (S) | ≤ 2 (S) | ≤ 2 (S) |
| Fosfomycin | > 128 (R) | > 128 (R) | > 128 (R) | ≤ 16 (S) | ≤ 16 (S) | ≤ 16 (S) | > 128 (R) | ≤ 16 (S) |
| Nitrofurantoin | ≤ 16 (S) | 32 (S) | ≤ 16 (S) | ≤ 16 (S) | ≤ 16 (S) | > 64 (R) | ≤ 16 (S) | ≤ 16 (S) |
| Tigecycline | ≤ 1 (S) | ≤ 1 (S) | ≤ 1 (S) | ≤ 1 (S) | ≤ 1 (S) | ≤ 1 (S) | ≤ 1 (S) | ≤ 1 (S) |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Peng, Z.; Li, X.; Hu, Z.; Li, Z.; Lv, Y.; Lei, M.; Wu, B.; Chen, H.; Wang, X. Characteristics of Carbapenem-Resistant and Colistin-Resistant Escherichia coli Co-Producing NDM-1 and MCR-1 from Pig Farms in China. Microorganisms 2019, 7, 482. https://doi.org/10.3390/microorganisms7110482
Peng Z, Li X, Hu Z, Li Z, Lv Y, Lei M, Wu B, Chen H, Wang X. Characteristics of Carbapenem-Resistant and Colistin-Resistant Escherichia coli Co-Producing NDM-1 and MCR-1 from Pig Farms in China. Microorganisms. 2019; 7(11):482. https://doi.org/10.3390/microorganisms7110482
Chicago/Turabian StylePeng, Zhong, Xiaosong Li, Zizhe Hu, Zugang Li, Yujin Lv, Minggang Lei, Bin Wu, Huanchun Chen, and Xiangru Wang. 2019. "Characteristics of Carbapenem-Resistant and Colistin-Resistant Escherichia coli Co-Producing NDM-1 and MCR-1 from Pig Farms in China" Microorganisms 7, no. 11: 482. https://doi.org/10.3390/microorganisms7110482
APA StylePeng, Z., Li, X., Hu, Z., Li, Z., Lv, Y., Lei, M., Wu, B., Chen, H., & Wang, X. (2019). Characteristics of Carbapenem-Resistant and Colistin-Resistant Escherichia coli Co-Producing NDM-1 and MCR-1 from Pig Farms in China. Microorganisms, 7(11), 482. https://doi.org/10.3390/microorganisms7110482