Comparative Transcriptomic Analysis of the Interaction between Penicillium expansum and Apple Fruit (Malus pumila Mill.) during Early Stages of Infection




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Fruit
2.2. Pathogen
2.3. Inoculation of Apples with P. expansum
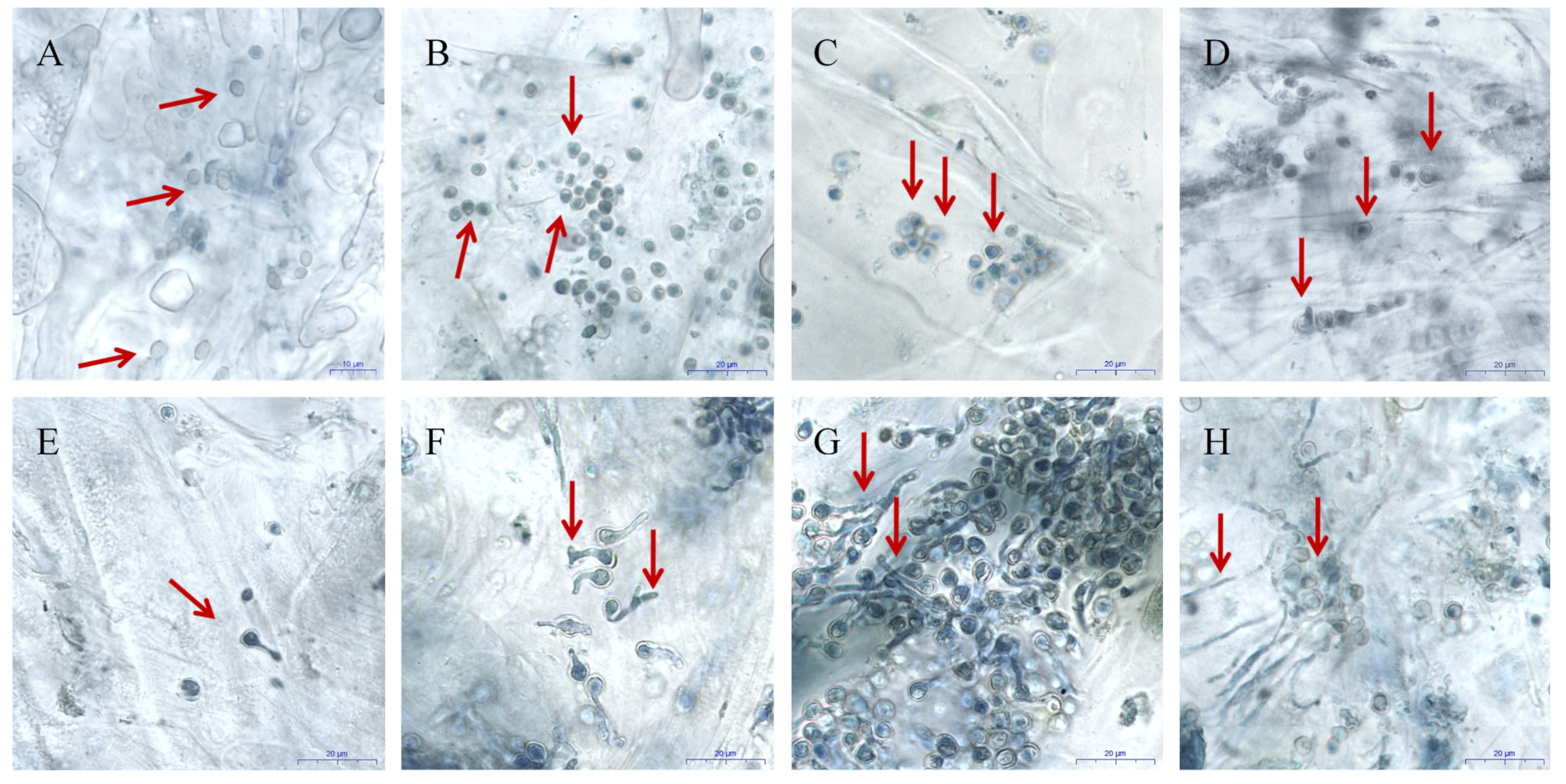
2.4. Microscopy
2.5. RNA Extraction
2.6. RNA-Seq Library Construction and Illumina Sequencing
2.7. Bioinformatic Analysis of the RNA-Seq Data
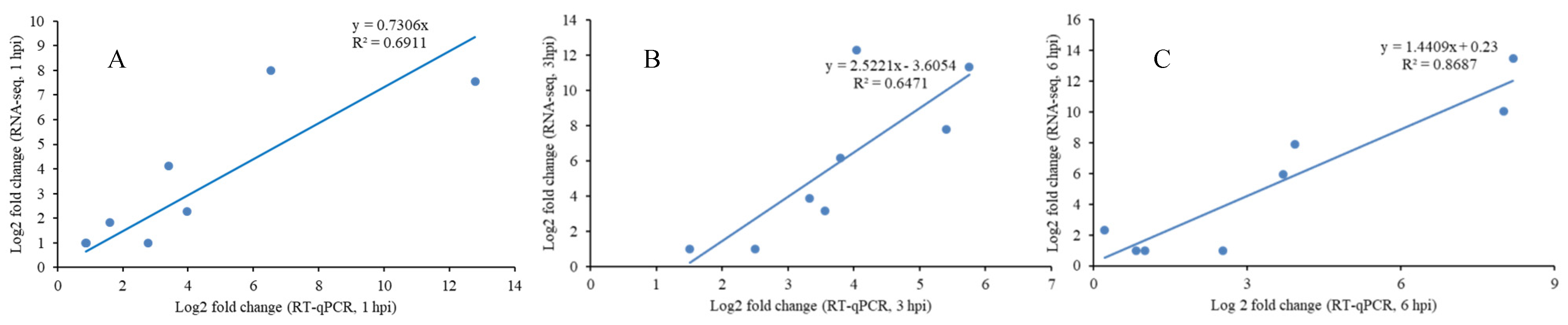
2.8. Validation of RNA-Seq Data by RT-qPCR
2.9. Statistical Analysis
3. Results
3.1. Infection of Apples by P. expansum
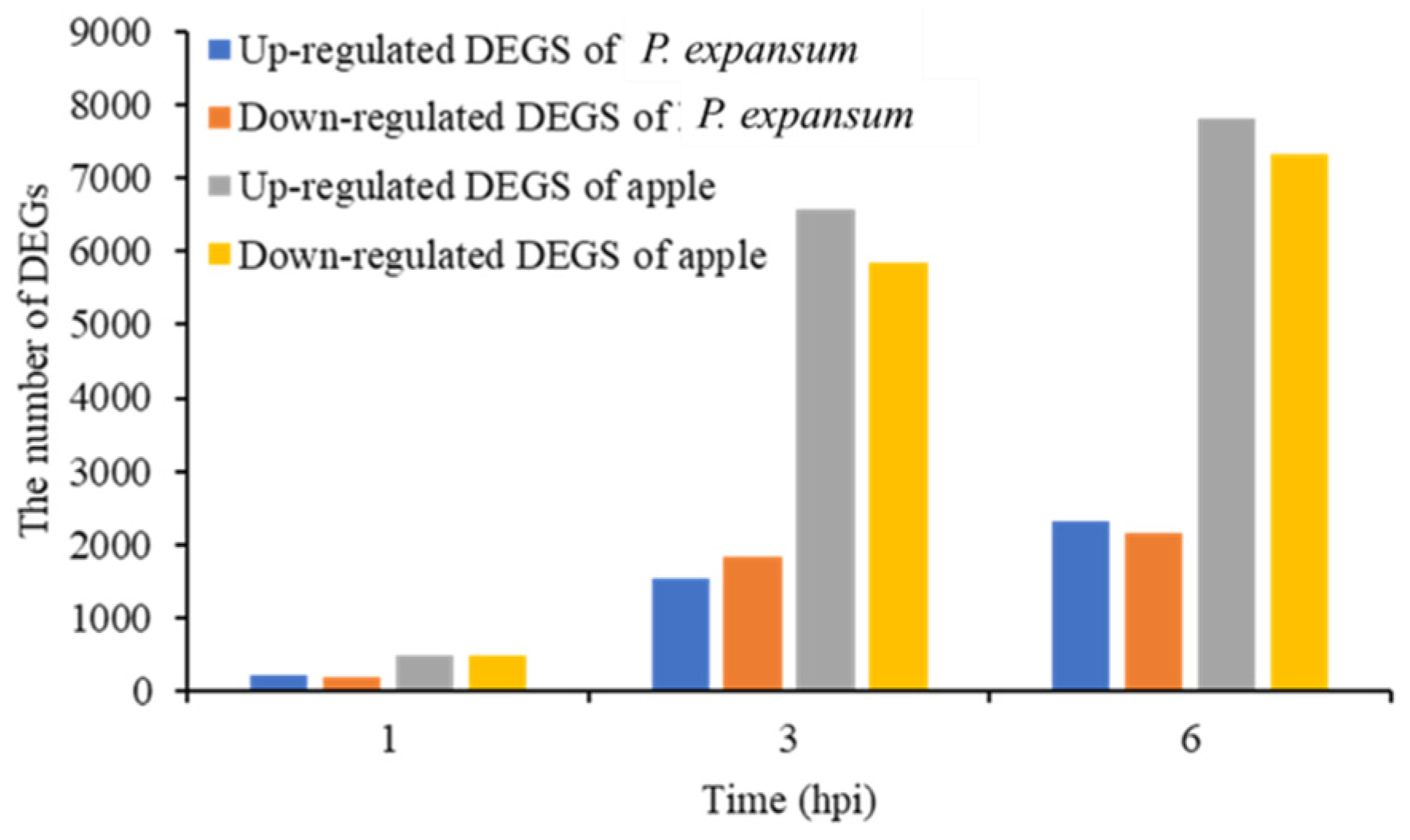
3.2. RNA-Seq Analysis and Validation
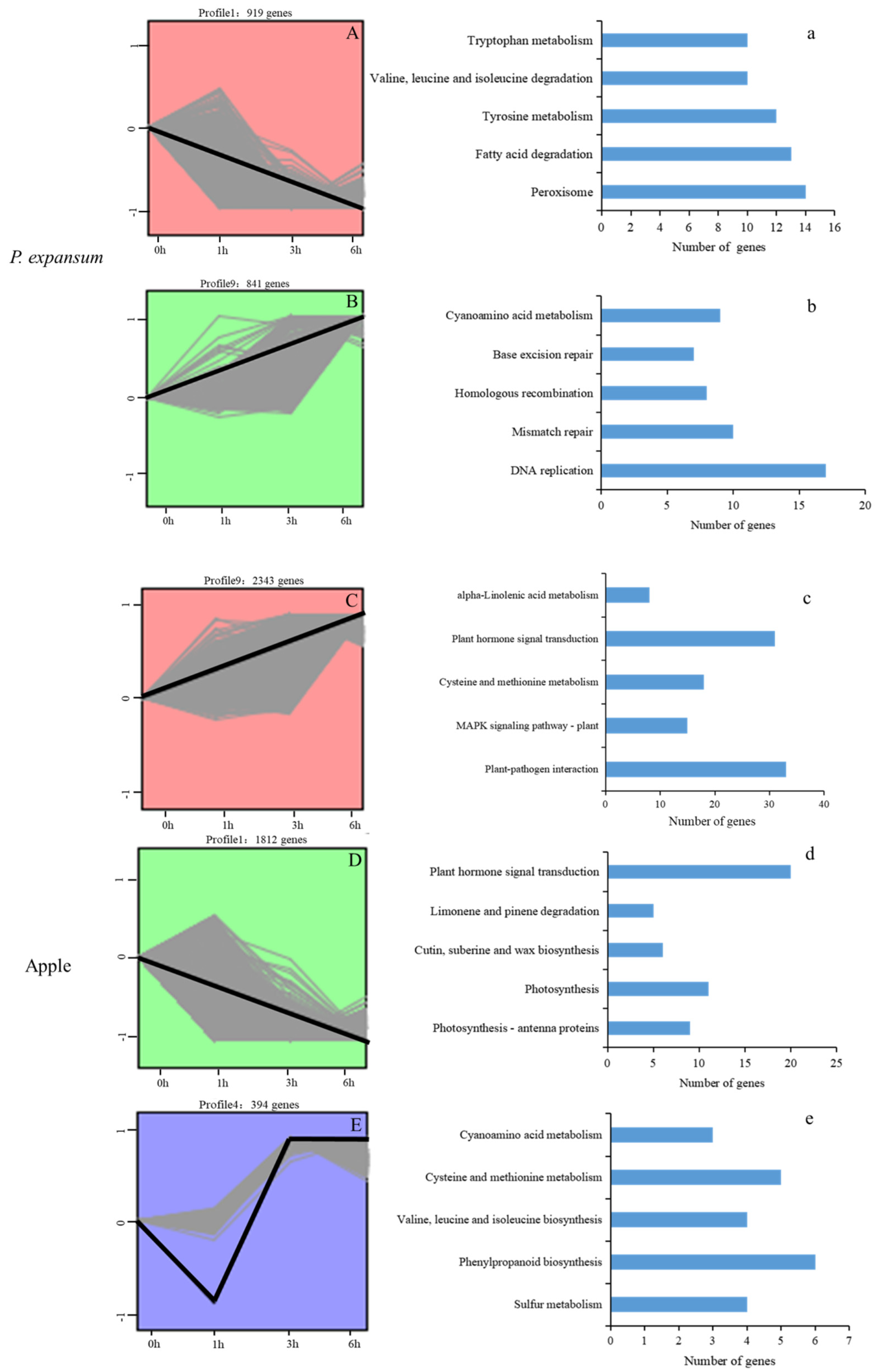
3.3. Gene Expression Pattern Analysis, Clustering and Functional Enrichment of DEGs
3.4. Major DEGs in P. expansum Identified During the Early Infection Process in Apple
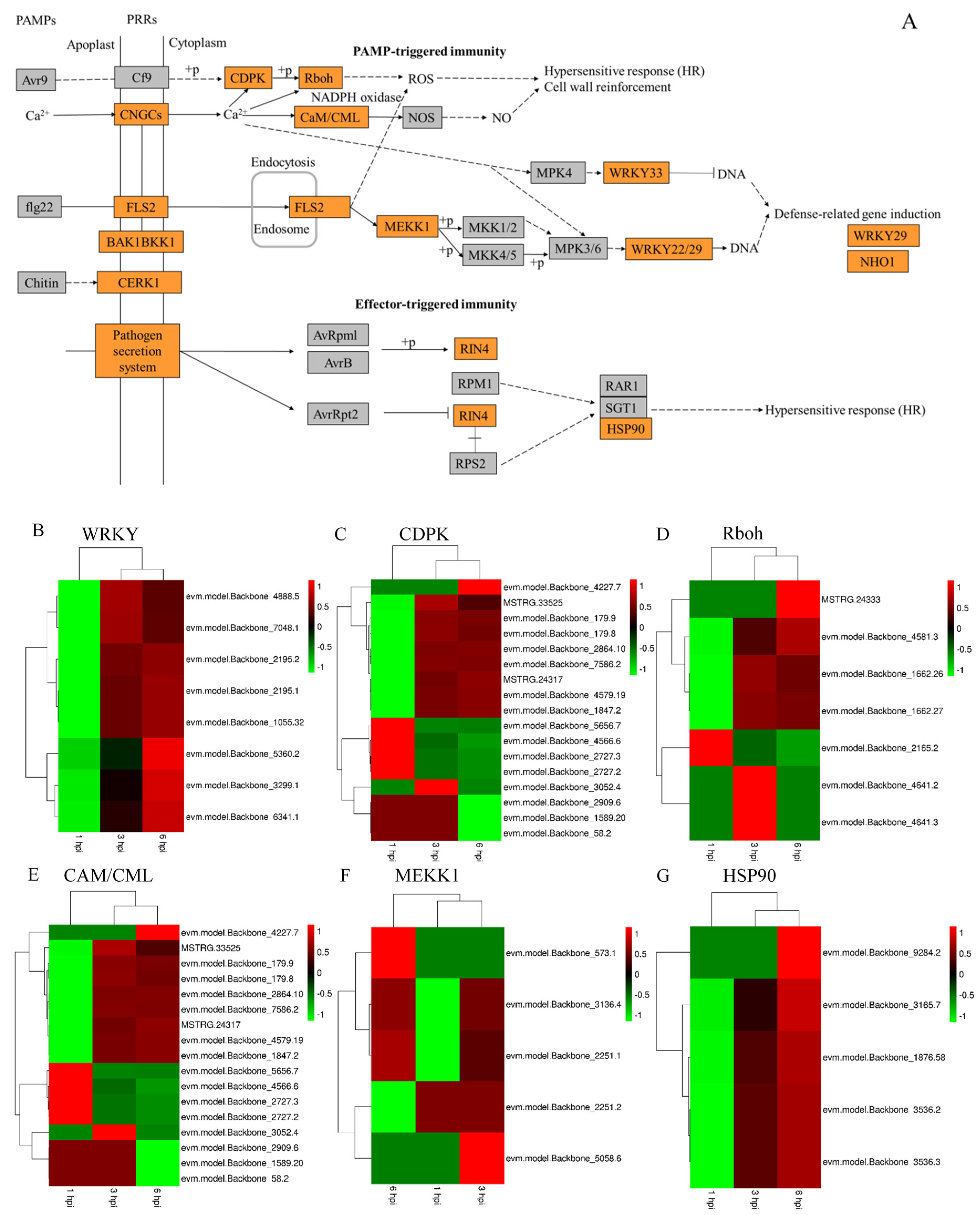
3.5. Major DEGs in Apple
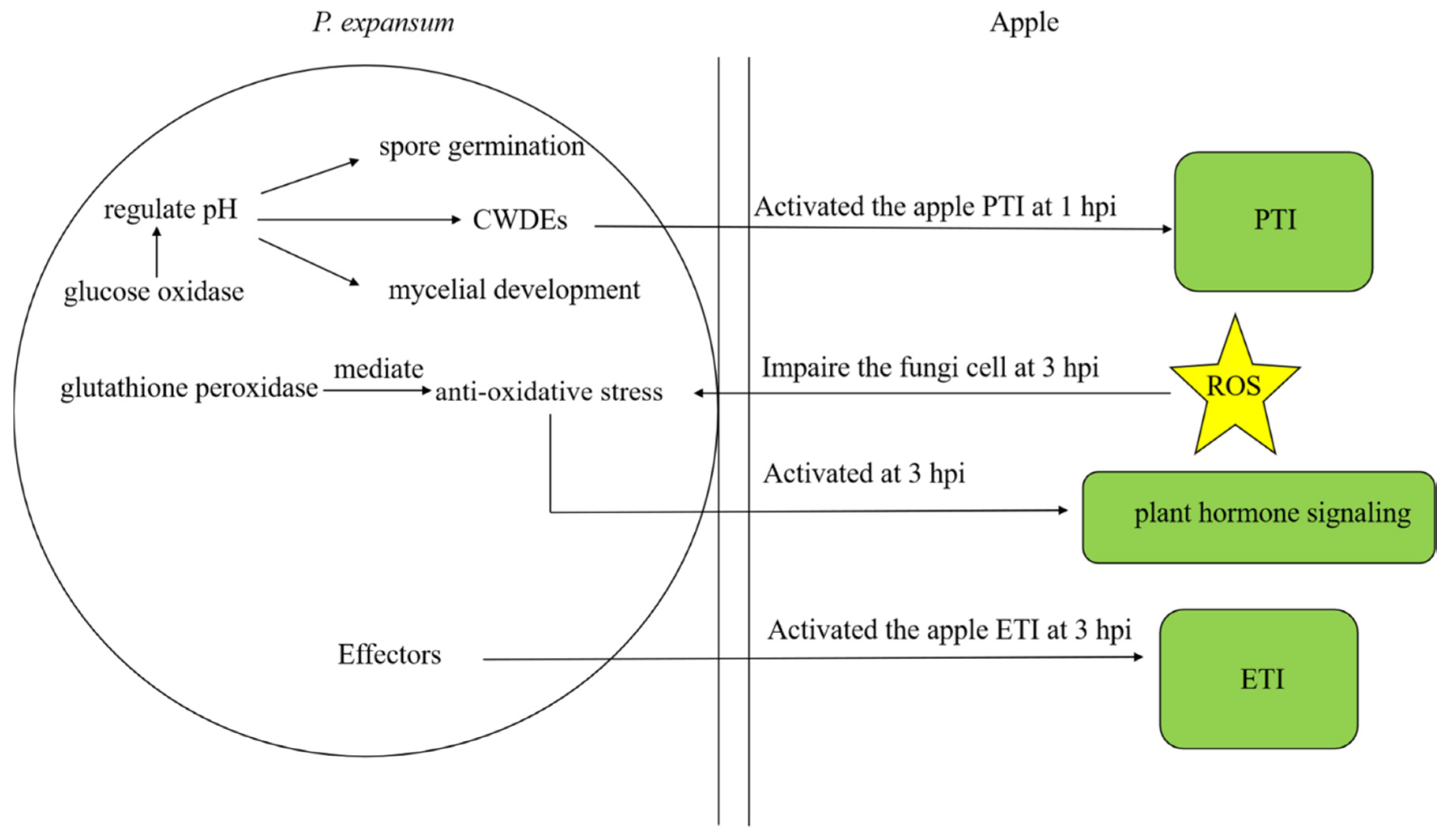
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ge, Y.; Wei, M.; Li, C.; Chen, Y.; Duan, B. Changes in the sucrose metabolism in apple fruit following postharvest acibenzolar-S-methyl treatment. J. Sci. Food Agric. 2018, 99, 1519–1524. [Google Scholar] [CrossRef] [PubMed]
- Wallace, R.L.; Hirkala, D.L.; Nelson, L.M. Postharvest biological control of blue mold of apple by Pseudomonas fluorescens during commercial storage and potential modes of action. Postharvest Biol. Technol. 2017, 33, 1–11. [Google Scholar] [CrossRef]
- Quaglia, M.; Ederli, L.; Pasqualini, S.; Zazzerini, A. Biological control agents and chemical inducers of resistance for postharvest control of Penicillium expansum Link. on apple fruit. Postharvest Biol. Technol. 2011, 59, 307–315. [Google Scholar] [CrossRef]
- Salomão, B.C.; Aragão, G.M.; Churey, J.J.; Worobo, R.W. Efficacy of sanitizing treatments against Penicillium expansum inoculated on six varieties of apples. J. Food Protect. 2008, 71, 643–647. [Google Scholar] [CrossRef] [PubMed]
- Spadaro, D.; Droby, S. Development of biocontrol products for postharvest diseases of fruit, the importance of elucidating the mechanisms of action of yeast antagonists. Trends Food Sci. Technol. 2016, 47, 39–49. [Google Scholar] [CrossRef]
- Schirra, M.; D’Aquino, S.; Cabras, P.; Angioni, A. Control of postharvest diseases of fruit by heat and fungicides, efficacy, residue levels, and residue persistence. A review. J. Agric. Food Chem. 2011, 59, 8531–8542. [Google Scholar] [CrossRef]
- Liu, C. Biological Mechanisms of the Spoilage Mould Infection of Fresh-Cut Apple and Its Prevention-Control by Ozone. Ph.D. Thesis, Shenyang Agricultural University, Shengyang, China, 2016. [Google Scholar]
- Ballester, A.R.; Marcet-Houben, M.; Levin, E.; Sela, N.; Selma-Lázaro, C. Genome, Transcriptome, and Functional Analysis of Penicillium expansum Provide New Insights Into Secondary Metabolism and Pathogenicity. Mol. Plant-Microbe Interact. 2015, 28, 232–248. [Google Scholar] [CrossRef]
- Alkan, N.; Espeso, E.A.; Prusky, D. Virulence regulation of phytopathogenic fungi by pH. Antioxid. Redox Sign. 2013, 19, 1012–1025. [Google Scholar] [CrossRef]
- Yan, Y.; Zheng, X.F.; Apaliya, M.T.; Yang, H.J.; Zhang, H.Y. Transcriptome characterization and expression profile of defense-related genes in pear induced by Meyerozyma guilliermondii. Postharvest Biol. Technol. 2018, 141, 63–70. [Google Scholar] [CrossRef]
- Tao, K. Molecular and Cellular Biology of Interactions between Phytophthora sojae and Host Soybean. Master’s Thesis, Nanjing Agricultural University, Nangjing, China, 2012. [Google Scholar]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(t)) method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Wang, K.; Lin, Z.; Zhang, H.; Zhang, X.; Zheng, X. Investigating proteome and transcriptome response of Cryptococcus podzolicus Y3 to citrinin and the mechanisms involved in its degradation. Food Chem. 2019, 283, 345–352. [Google Scholar] [CrossRef] [PubMed]
- Taheri, P. Cereal diseases caused by Fusarium graminearum: From biology of the pathogen to oxidative burst-related host defense responses. Eur. J. Plant Pathol. 2018, 152, 1–20. [Google Scholar] [CrossRef]
- Rossi, F.R.; Krapp, A.R.; Bisaro, F.; Maiale, S.J.; Pieckenstain, F.L. Reactive oxygen species generated in chloroplasts contribute to tobacco leaf infection by the necrotrophic fungus Botrytis cinerea. Plant J. 2017, 92, 761–773. [Google Scholar] [CrossRef] [PubMed]
- Møller, I.M.; Jensen, P.E.; Hansson, A. Oxidative modifications to cellular components in plants. Annu. Rev. Plant Biol. 2007, 58, 459–481. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Torres, P.; Vilanova, L.; Ballester, A.R.; Pérezc, M.L.; ópez- Teixidó, N. Unravelling the contribution of the Penicillium expansum PeSte12 transcription factor to virulence during apple fruit infection. Food Microbiol. 2018, 69, 123–135. [Google Scholar] [CrossRef]
- Prusky, D.; Barad, S.; Luria, N.; Ment, D. pH modulation of host environment, a mechanism modulating fugal attack in postharvest pathogen interactions. Postharvest Pathol. 2014, 11–25. [Google Scholar]
- Gust, A.A.; Pruitt, R.; Nürnberger, T. Sensing Danger: Key to Activating Plant Immunity. Trends Plant Sci. 2017, 22, 779–791. [Google Scholar] [CrossRef]
- Friesen, T.L.; Faris, J.D.; Solomon, P.S.; Oliver, R.P. Host-specific toxins, effectors of necrotrophic pathogenicity. Cell Microbiol. 2008, 10, 1421–1428. [Google Scholar] [CrossRef]
- Demidchik, V.; Shabala, S.; Isayenkov, S.; Cuin, T.; Pottosin, I. Calcium transport across plant membranes, mechanisms and functions. New Phytol. 2018, 220, 49–69. [Google Scholar] [CrossRef]
- Mori, I.C.; Nobukiyo, Y.; Nakahara, Y.; Shibasaka, M.; Furuichi, T. A Cyclic Nucleotide-Gated Channel, HvCNGC2-3, Is Activated by the Co-Presence of Na+ and K+ and Permeable to Na+ and K+ Non-Selectively. Plants 2018, 7, 61. [Google Scholar] [CrossRef]
- Suzuki, M.; Shibuya, M.; Shimada, H.; Motoyama, N.; Nakashima, M. Autophosphorylation of specific threonine and tyrosine residues in Arabidopsis CERK1 is essential for the activation of chitin-induced immune signaling. Plant Cell Physiol. 2016, 57, 2312. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Zhou, T.; Wu, T.; Li, X. Saccharomyces cerevisiae YE-7 reduces the risk of apple blue mold disease by inhibiting the fungal incidence and patulin biosynthesis. J. Food Process. Preserv. 2018, 42, e13360. [Google Scholar] [CrossRef]
- Zhou, T.; Wang, X.H.; Luo, J.; Ye, B.S.; Zhou, Y.Y. Identification of differentially expressed genes involved in spore germination of Penicillium expansum by comparative transcriptome and proteome approaches. Microbiol. Open 2017, 8, e654. [Google Scholar] [CrossRef] [PubMed]
- Van Leeuwen, M.R.; Van Doorn, T.M.; Golovina, E.A.; Stark, J.; Dijksterhuis, J. Water and air distributed conidia differ in sterol content and cytoplasmic microviscosity. Appl. Environ. Microbiol. 2010, 76, 366–369. [Google Scholar] [CrossRef]
- Maruyama, J.; Kitamoto, K. Expanding functional repertoires of fungal peroxisomes, contribution to growth and survival processes. Front. Physiol. 2013, 4, 177. [Google Scholar] [CrossRef]
- Tang, J.Z.; Ding, Y.X.; Nan, J.; Yang, X.Y.; Sun, L. Transcriptome sequencing and ITRAQ reveal the detoxification mechanism of Bacillus GJ1, a potential biocontrol agent for Huanglongbing. PLoS ONE 2018, 13, e0200427. [Google Scholar] [CrossRef]
- Barad, S.; Sela, N.; Verma, D.; Dubey, A.; Glam, G.; Sherman, A.; Prusky, D. Fungal and host transcriptome analysis of pH-regulated genes during colonization of apple fruits by Penicillium expansum. BMC Genom. 2016, 17, 330. [Google Scholar] [CrossRef]
- Bent, A.F.; Mackey, D. Elicitors, effectors, and R genes, the new paradigm and a lifetime supply of questions. Annu. Rev. Phytopathol. 2007, 45, 399–436. [Google Scholar] [CrossRef]
- Wu, G.; Liu, S.; Zhao, Y.; Wang, W.; Kong, Z.S. ENHANCED DISEASE RESISTANCE4 associates with CLATHRIN HEAVY CHAIN2 and modulates plant immunity by regulating relocation of EDR1 in Arabidopsis. Plant Cell 2015, 27, 857–873. [Google Scholar] [CrossRef]
- Kubicek, C.P.; Starr, T.L.; Glass, N.L. Plant Cell Wall–Degrading Enzymes and Their Secretion in Plant-Pathogenic Fungi. Annu. Rev. Phytopathol. 2014, 52, 427–451. [Google Scholar] [CrossRef]
- Minic, Z. Physiological roles of plant glycoside hydrolases. Planta 2008, 227, 723–740. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.L.; Yu, P.L.; Chung, K.R. The glutathione peroxidase-mediated reactive oxygen species resistance, fungicide sensitivity and cell wall construction in the citrus fungal pathogen alternaria alternata. Environ. Microbiol. 2016, 18, 923–935. [Google Scholar] [CrossRef] [PubMed]
- Prusky, D.; McEvoy, J.L.; Leverentz, B.; Conway, L. Local modulation of host pH by Colletotrichum species as a mechanism to increase virulence. Mol. Plant-Microbe Interact. 2001, 14, 1105–1113. [Google Scholar] [CrossRef] [PubMed]
- Vilanova, L.; Viñas, I.; Torres, R.; Usall, J.; Buron-Moles, G. Acidification of apple and orange hosts by Penicillium digitatum and Penicillium expansum. Int. J. Food Microbiol. 2014, 178, 39–49. [Google Scholar] [CrossRef]
- Barad, S.; Horowitz, S.B.; Kobiler, I.; Sherman, A.; Prusky, D. Accumulation of the mycotoxin patulin in the presence of gluconic acid contributes to pathogenicity of Penicillium expansum. Mol. Plant-Microbe Interact. 2014, 27, 66–77. [Google Scholar] [CrossRef]
- Lee, C.G.; Da Silva, C.A.; Lee, J.Y.; Hartl, D.; Elias, J.A. Chitin regulation of immune responses, an old molecule with new roles. Curr. Opin. Immunol. 2008, 20, 684–689. [Google Scholar] [CrossRef]
- de Jonge, R.; van Esse, H.P.; Kombrink, A.; Shinya, T.; Desaki, Y. Conserved fungal LysM effector Ecp6 prevents chitin-triggered immunity in plants. Science 2010, 329, 953–955. [Google Scholar] [CrossRef]






© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, K.; Zheng, X.; Zhang, X.; Zhao, L.; Yang, Q.; Boateng, N.A.S.; Ahima, J.; Liu, J.; Zhang, H. Comparative Transcriptomic Analysis of the Interaction between Penicillium expansum and Apple Fruit (Malus pumila Mill.) during Early Stages of Infection. Microorganisms 2019, 7, 495. https://doi.org/10.3390/microorganisms7110495
Wang K, Zheng X, Zhang X, Zhao L, Yang Q, Boateng NAS, Ahima J, Liu J, Zhang H. Comparative Transcriptomic Analysis of the Interaction between Penicillium expansum and Apple Fruit (Malus pumila Mill.) during Early Stages of Infection. Microorganisms. 2019; 7(11):495. https://doi.org/10.3390/microorganisms7110495
Chicago/Turabian StyleWang, Kaili, Xiangfeng Zheng, Xiaoyun Zhang, Lina Zhao, Qiya Yang, Nana Adwoa Serwah Boateng, Joseph Ahima, Jia Liu, and Hongyin Zhang. 2019. "Comparative Transcriptomic Analysis of the Interaction between Penicillium expansum and Apple Fruit (Malus pumila Mill.) during Early Stages of Infection" Microorganisms 7, no. 11: 495. https://doi.org/10.3390/microorganisms7110495
APA StyleWang, K., Zheng, X., Zhang, X., Zhao, L., Yang, Q., Boateng, N. A. S., Ahima, J., Liu, J., & Zhang, H. (2019). Comparative Transcriptomic Analysis of the Interaction between Penicillium expansum and Apple Fruit (Malus pumila Mill.) during Early Stages of Infection. Microorganisms, 7(11), 495. https://doi.org/10.3390/microorganisms7110495