Zymobacter palmae Pyruvate Decarboxylase is Less Effective Than That of Zymomonas mobilis for Ethanol Production in Metabolically Engineered Synechocystis sp. PCC6803


 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains
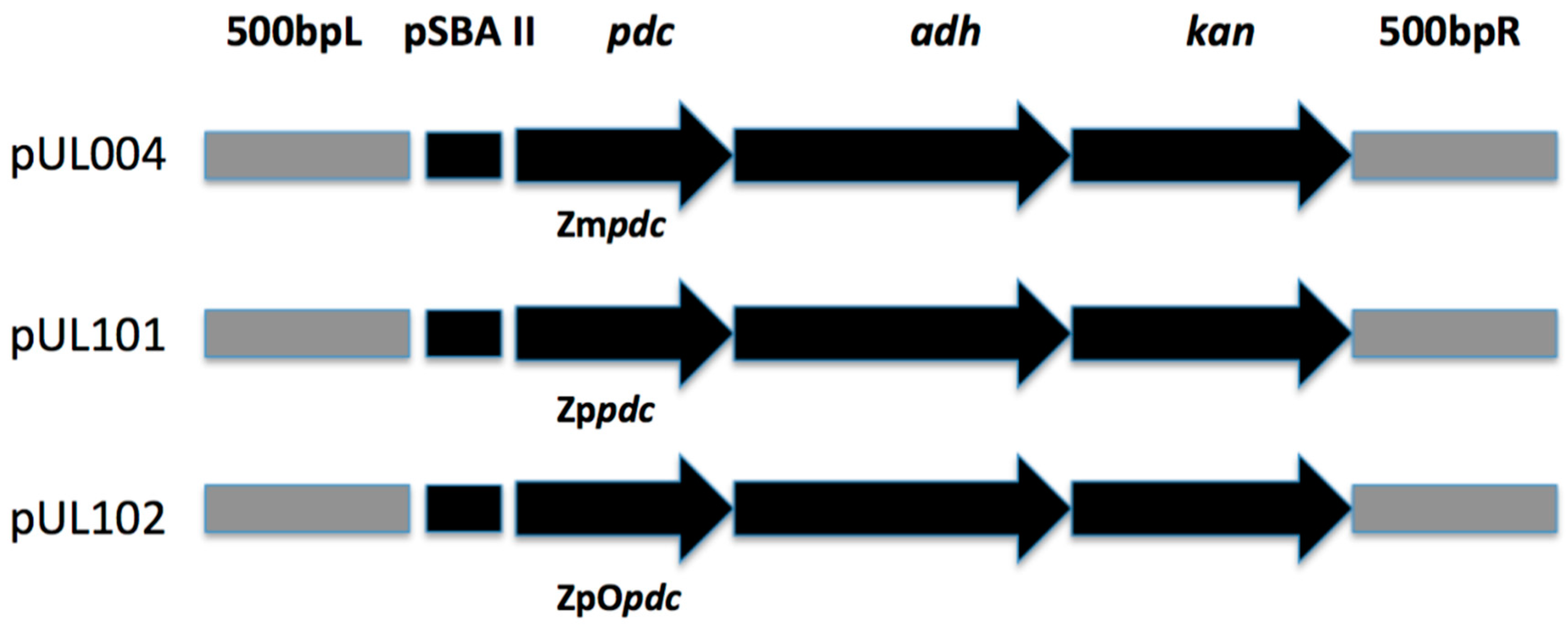
2.2. Gene Cloning and Strain Construction
2.3. Growth Measurements
2.4. Ethanol Determination
2.5. Assay for PDC from Crude Recombinant Extracts of Engineered Synechocystis sp PCC6803
2.6. Assays for Acetaldehyde in Recombinant Strains of Synechocystis sp PCC6803
3. Results and Discussion
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Pembroke, J.T.; Armshaw, P.; Ryan, M.P. Metabolic Engineering of the Model Photoautotrophic Cyanobacterium Synechocystis for Ethanol Production: Optimization Strategies and Challenges. In Fuel Ethanol Production from Sugarcane; Basso, T.P., Basso, L.C., Eds.; Intech Open: London, UK, 2019; pp. 199–219. [Google Scholar] [CrossRef]
- Bender, M.H. Potential conservation of biomass in the production of synthetic organics. Resour. Conserv. Recycl. 2000, 30, 49–58. [Google Scholar] [CrossRef]
- Demirbas, M.F. Current technologies for biomass conversion into chemicals and fuels. Energy Sources Part A Recover. Util. Environ. Eff. 2006, 28, 1181–1188. [Google Scholar] [CrossRef]
- Osamu, K.; Carl, H. Biomass Handbook; Gordon and Breach Science Publisher: New York, NY, USA, 1989. [Google Scholar]
- Touloupakis, E.; Cicchi, B.; Benavides, A.M.S.; Torzillo, G. Effect of high pH on growth of Synechocystis sp. PCC 6803 cultures and their contamination by golden algae (Poterioochromonas sp.). Appl. Microbiol. Biotechnol. 2016, 100, 1333–1341. [Google Scholar] [CrossRef] [PubMed]
- Rippka, R.; Deruelles, J.; Waterbury, J.B.; Herdman, M.; Stanier, R.Y. Generic assignments, strain histories and properties of pure cultures of cyano-bacteria. J. Gen. Microbiol. 1979, 111, 1–61. [Google Scholar]
- Dexter, J.; Armshaw, P.; Sheahan, C.; Pembroke, J.T. The state of autotrophic ethanol production in Cyanobacteria. J. Appl. Microbiol. 2015, 119, 11–24. [Google Scholar] [CrossRef]
- Ingram, L.O.; Conway, T.; Clark, D.P.; Sewell, G.W.; Preston, J.F. Genetic engineering of ethanol production in Escherichia coli. Appl. Environ. Microbiol. 1987, 53, 2420–2425. [Google Scholar]
- Deng, M.D.; Coleman, J.R. Ethanol synthesis by genetic engineering in cyanobacteria. Appl. Environ. Microbiol. 1999, 65, 523–528. [Google Scholar]
- Dexter, J.; Fu, P. Metabolic engineering of cyanobacteria for ethanol production. Energy Environ. Sci. 2009, 2, 857. [Google Scholar] [CrossRef]
- Gao, Z.; Zhao, H.; Li, Z.; Tan, X.; Lu, X. Photosynthetic production of ethanol from carbon dioxide in genetically engineered cyanobacteria. Energy Environ. Sci. 2012, 5, 9857–9865. [Google Scholar] [CrossRef]
- Armshaw, P.; Carey, D.; Sheahan, C.; Pembroke, J.T. Utilising the native plasmid, pCA2.4, from the cyanobacterium Synechocystis sp. strain PCC6803 as a cloning site for enhanced product production. Biotechnol. Biofuels 2015, 8. [Google Scholar] [CrossRef]
- Angermayr, S.A.; Paszota, M.; Hellingwerf, K.J. Engineering a cyanobacterial cell factory for production of lactic acid. Appl. Environ. Microbiol. 2012, 78, 7098–7106. [Google Scholar] [CrossRef] [PubMed]
- Angermayr, S.A.; van der Woude, A.D.; Correddu, D.; Vreugdenhil, A.; Verrone, V.; Hellingwerf, K.J. Exploring metabolic engineering design principles for the photosynthetic production of lactic acid by Synechocystis sp. PCC6803. Biotechnol. Biofuels 2014, 7. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.-H.; Lindblad, P. Wide-dynamic-range promoters engineered for cyanobacteria. J. Biol. Eng. 2013, 7, 10. [Google Scholar] [CrossRef] [PubMed]
- Lewicka, A.J.; Lyczakowski, J.J.; Blackhurst, G.; Pashkuleva, C.; Rothschild-Mancinelli, K.; Tautvaišas, D.; Thornton, H.; Villanueva, H.; Xiao, W.; Slikas, J.; et al. Fusion of pyruvate decarboxylase and alcohol dehydrogenase increases ethanol production in Escherichia coli. ACS Synth. Biol. 2014, 3, 976–978. [Google Scholar] [CrossRef]
- Murphy, K.C. Use of bacteriophage λ recombination functions to promote gene replacement in Escherichia coli. J. Bacteriol. 1998, 180, 2063–2071. [Google Scholar]
- Böltner, D.; MacMahon, C.; Pembroke, J.T.; Strike, P.; Osborn, A.M. R391: A conjugative integrating mosaic comprised of phage, plasmid, and transposon elements. J. Bacteriol. 2002, 184, 5158–5169. [Google Scholar] [CrossRef]
- Lopes da Silva, T.; Passarinho, P.C.; Galriça, R.; Zenóglio, A.; Armshaw, P.; Pembroke, J.T.; Sheahan, C.; Reis, A.; Gírio, F. Evaluation of the ethanol tolerance for wild and mutant Synechocystis strains by flow cytometry. Biotechnol. Rep. 2018, 17, 137–147. [Google Scholar] [CrossRef]
- Raj, K.C.; Ingram, L.O.; Maupin-Furlow, J.A. Pyruvate decarboxylase: A key enzyme for the oxidative metabolism of lactic acid by Acetobacter pasteurianus. Arch. Microbiol. 2001, 176, 443–451. [Google Scholar]
- Takahashi, H.; Uchimiya, H.; Hihara, Y. Difference in metabolite levels between photoautotrophic and photomixotrophic cultures of Synechocystis sp. PCC 6803 examined by capillary electrophoresis electrospray ionization mass spectrometry. J. Exp. Botany. 2008, 59, 3009–3018. [Google Scholar] [CrossRef]
- Raj, K.C.; Talarico, L.A.; Ingram, L.O.; Maupin-Furlow, J.A. Cloning and characterization of the Zymobacter palmae pyruvate decarboxylase gene (PDC) and comparison to bacterial homologues. Appl. Environ. Microbiol. 2002, 68, 2869–2876. [Google Scholar] [CrossRef]
- Buddrus, L.; Andrews, E.S.V.; Leak, D.J.; Danson, M.J.; Arcus, V.L.; Crennell, S.J. Crystal structure of pyruvate decarboxylase from Zymobacter palmae. Acta Crystallogr. Sect. F Struct. Biol. Commun. 2016, 72, 700–706. [Google Scholar] [CrossRef] [PubMed]
- Van Zyl, L.J.; Schubert, W.D.; Tuffin, M.I.; Cowan, D.A. Structure and functional characterization of pyruvate decarboxylase from Gluconacetobacter diazotrophicus. BMC Struct. Biol. 2014, 14. [Google Scholar] [CrossRef] [PubMed]
- Dobritzsch, D.; König, S.; Schneider, G.; Lu, G. High resolution crystal structure of pyruvate decarboxylase from Zymomonas mobilis. Implications for substrate activation in pyruvate decarboxylases. J. Biol. Chem. 1998, 273, 20196–20204. [Google Scholar] [CrossRef] [PubMed]
- Gocke, D.; Berthold, C.L.; Schneider, G.; Pohl, H. Holostructure of pyruvate decarboxylase from Acetobacter pasteurianus. Protein Data Bank (PDB) PDB ID: 2VBI 2008. Available online: http://www.rcsb.org/pdb/explore.do?structureId=2vbi (accessed on 6 July 2019).
- Van Zyl, L.J.; Taylor, M.P.; Eley, K.; Tuffin, M.; Cowan, D.A. Engineering pyruvate decarboxylase-mediated ethanol production in the thermophilic host Geobacillus thermoglucosidasius. Appl. Microbiol. Biotechnol. 2014, 98, 1247–1259. [Google Scholar] [CrossRef]
- Liu, S.; Dien, B.S.; Cotta, M.A. Functional expression of bacterial Zymobacter palmae pyruvate decarboxylase gene in Lactococcus lactis. Curr. Microbiol. 2005, 50, 324–328. [Google Scholar] [CrossRef]
- Kunjapur, A.M.; Prather, K.L.J. Microbial engineering for aldehyde synthesis. Appl. Environ. Microbiol. 2015, 81, 1892–1901. [Google Scholar] [CrossRef]
- Alcover, N.; Carceller, A.; Álvaro, G.; Guillén, M. Zymobacter palmae pyruvate decarboxylase production process development: Cloning in Escherichia coli, fed-batch culture and purification. Eng. Life Sci. 2019, 19, 502–512. [Google Scholar] [CrossRef]
- Eriksson, P.C.J. Measurement of acetaldehyde: What levels occur naturally and in response to alcohol? In Acetaldehyde Related Pathology: Bridging the Transdisciplinary Divide: Novartis Foundation Symposium; Chadwick, D.J., Goode, J., Eds.; Wiley: Hoboken, NJ, USA, 2007; pp. 247–255. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Strain | Genotype/phenotype | Source |
| BL21 (DE3) * | E. coli B F– ompT, gal, dcm, lon, hsdSB(rB–mB–), λ(DE3 [lacI lacUV5-T7p07 ind1, sam7, nin5]) [malB+]K-12(λS) | Thermo Fisher Scientific Ballycoolin, Dublin 15, Ireland |
| DSM-10491 | Zymobacter palmae | DSMZ—German Collection of Microorganisms and Cell Cultures GmbH |
| AA314 | Wild-type Synechocystis PCC6803 strain | K. Hellingwerf, UvA, Amsterdam, The Netherlands |
| UL004 | PCC6803 transconjugant with ZmPDC, slr1192- adhA, psbA2 locus, KanR | [19] |
| UL059 | PCC6803 transconjugant with Zymomonas mobilis adh, ptrc promoter | This study |
| UL070 | PCC6803 transconjugant with ZmPDC, slr1192- adhA, psbA2 locus, KanR | This study |
| UL071 | PCC6803 transconjugant with ZpPDC, slr1192- adhA, psbA2 locus, KanR | This study |
| UL072 | PCC6803 transconjugant with ZpOpdc, slr1192- adhA, psbA2 locus, KanR | This study |
| Plasmid | Genotype/phenotype | Source |
| pUC18 | AmpR backbone plasmid | Sigma-Aldrich, Arklow, Wicklow, Ireland |
| pUL004 | pUC18 backbone, PpsbA2 promoter, ZmPDC, slr1192- adhA, psbA2 integration site, KanR | [19] |
| pUL101 | pUC18 backbone PpsbA2 promoter, ZpPDC, slr1192- adhA, psbA2 integration site, KanR | This study |
| pUL102 | pUC18 backbone PpsbA2 promoter, ZpOpdc, slr1192- adhA, psbA2 integration site, KanR | This study |
| Primers | Sequence (5′–3′) |
|---|---|
| Zymobacter palmae pdc primers for the psbA2 vector to create pUL101 | |
| ZppdcF1 | AGGAATTATAACCATATGTATACCGTTGGTATGTACTTGG |
| ZppdcR1 | GATCCCCAAAAACTACGCTTGTGGTTTGCGAGAGTTGG |
| Codon optimized Zymobacter palmae pdc for the psbA2 vector to create pUL101 | |
| coZppdcF | AGGAATTATAACCATATGTATACCGTTGGTATGTATTTGG |
| coZppdcR | GATCCCCAAAAACTATGCCTGGGGCTTCCGGGAATTGG |
| Linearize the psbA2 vector to create pUL101 and pUL102 | |
| PSBAII F | TAGTTTTTGGGGATCAATTC |
| PSBAII R | ATGGTTATAATTCCTTATGTATTTG |
| Sequencing and screening primers for the ethanol cassette (psbA2 promoter, pdc, slr1192 adhA, kan) in the psbA2 vector | |
| P9F | GTCAGTTCCAATCTGAACATCGA |
| P35F | CTCTACACAGCCCAGAACTATGG |
| P13R | CAATTTGCAGATTATTCAGTTGGCAT |
| PDC | ZmPDC | ZpPDC | ApPDC | SvPDC | GoPDC | GdPDC |
|---|---|---|---|---|---|---|
| PDB entry | 1ZPD | 5EUJ | 2VBI | N/A | N/A | 4COK |
| Bacterial species | Zymomonas mobilis | Zymobacter palmae | Acetobacter pasteurianus | Sarcina ventriculi | Gluconobacter oxydans | Gluconacetobacter diazotrophicus |
| Gram | Negative | Negative | Negative | Positive | Negative | Negative |
| Gene | M15393 | AF474145 | AF368435 | AAL18557 | KF650839 | KJ746104 |
| Protein | AAA27696 | AAM49566 | AAM21208 | AF354297 | AHB37781 | AIG13066 |
| Amino acid identity % | *62/63 | Reference | 73 | 31 | 67 | 71 |
| Kinetics | * M–M | * M–M | * M–M | * Sigmoidal | * M–M | * M–M |
| Km mM (pH) | * 0.43(6.0) 0.94 (7.0) | * 0.24 (6.0) 0.71 (7.0) | * 0.39 (5.0) 5.10 (7.0) | * 5.7 (6.5) 4.0 (7.0) | # 0.12 (5.0) 1.20 (6.5) 2.80 (7.0) | # 0.06 (5.0) 0.60 (6.0) 1.20 (7.0) |
| Optimum Temperature °C | * 60 | * 55 | * 65 | N/A | #53 | # 45–50 |
| Optimum pH | * 6.0 | * 5.5–6.0 | * 5.0–5.5 | * 6.3–6.7 | # 4.5–5.0 | # 5.0–5.5 |
| Reference | [24,25] | [23] | [26] | [22] | [27] | [24] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Quinn, L.; Armshaw, P.; Soulimane, T.; Sheehan, C.; Ryan, M.P.; Pembroke, J.T. Zymobacter palmae Pyruvate Decarboxylase is Less Effective Than That of Zymomonas mobilis for Ethanol Production in Metabolically Engineered Synechocystis sp. PCC6803. Microorganisms 2019, 7, 494. https://doi.org/10.3390/microorganisms7110494
Quinn L, Armshaw P, Soulimane T, Sheehan C, Ryan MP, Pembroke JT. Zymobacter palmae Pyruvate Decarboxylase is Less Effective Than That of Zymomonas mobilis for Ethanol Production in Metabolically Engineered Synechocystis sp. PCC6803. Microorganisms. 2019; 7(11):494. https://doi.org/10.3390/microorganisms7110494
Chicago/Turabian StyleQuinn, Lorraine, Patricia Armshaw, Tewfik Soulimane, Con Sheehan, Michael P. Ryan, and J. Tony Pembroke. 2019. "Zymobacter palmae Pyruvate Decarboxylase is Less Effective Than That of Zymomonas mobilis for Ethanol Production in Metabolically Engineered Synechocystis sp. PCC6803" Microorganisms 7, no. 11: 494. https://doi.org/10.3390/microorganisms7110494
APA StyleQuinn, L., Armshaw, P., Soulimane, T., Sheehan, C., Ryan, M. P., & Pembroke, J. T. (2019). Zymobacter palmae Pyruvate Decarboxylase is Less Effective Than That of Zymomonas mobilis for Ethanol Production in Metabolically Engineered Synechocystis sp. PCC6803. Microorganisms, 7(11), 494. https://doi.org/10.3390/microorganisms7110494