A Mesophilic Aeromonas salmonicida Strain Isolated from an Unsuspected Host, the Migratory Bird Pied Avocet



Abstract
:
1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Growth Conditions
2.2. DNA Extraction and Sequencing
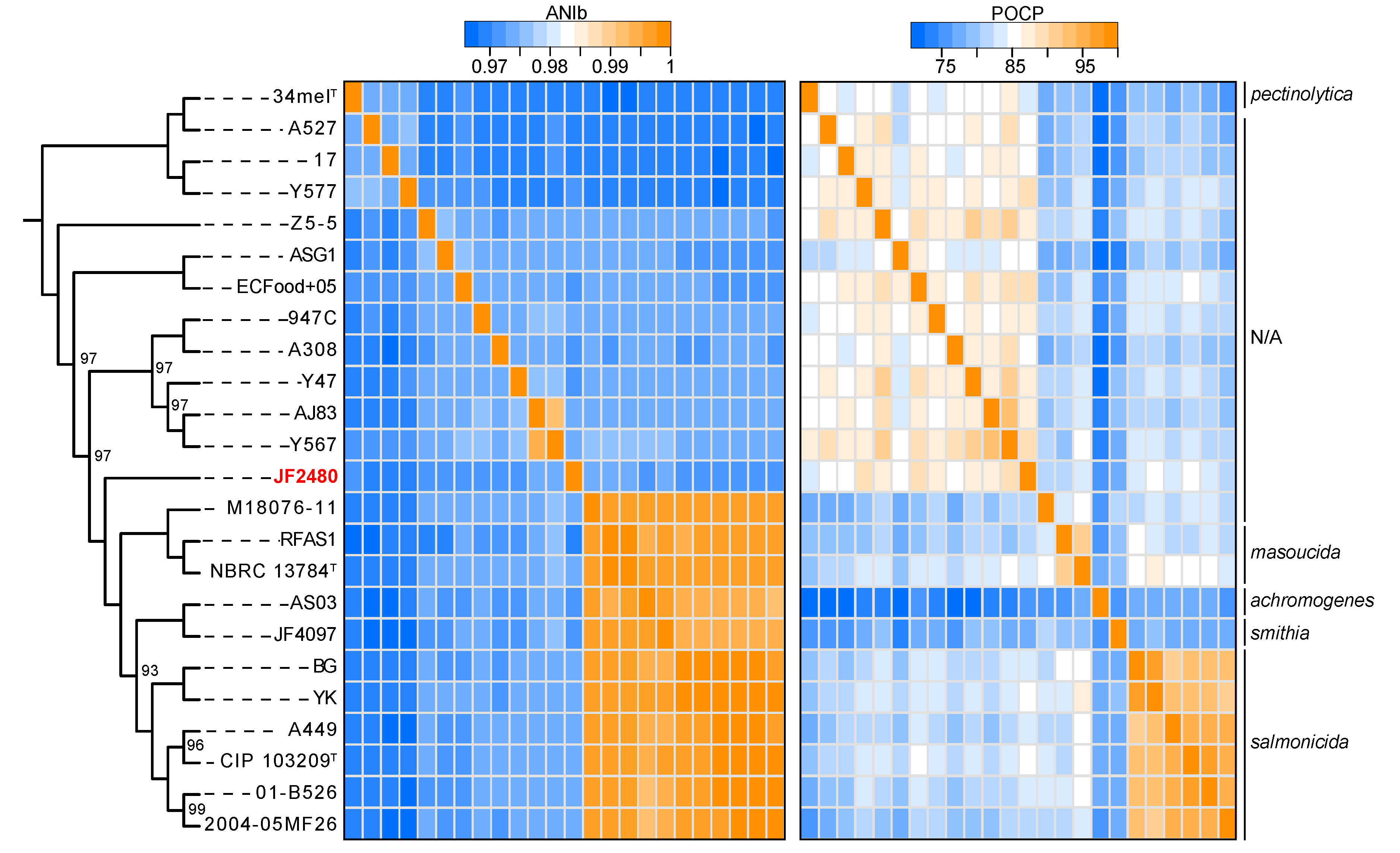
2.3. Sequence Assembly and Analyses
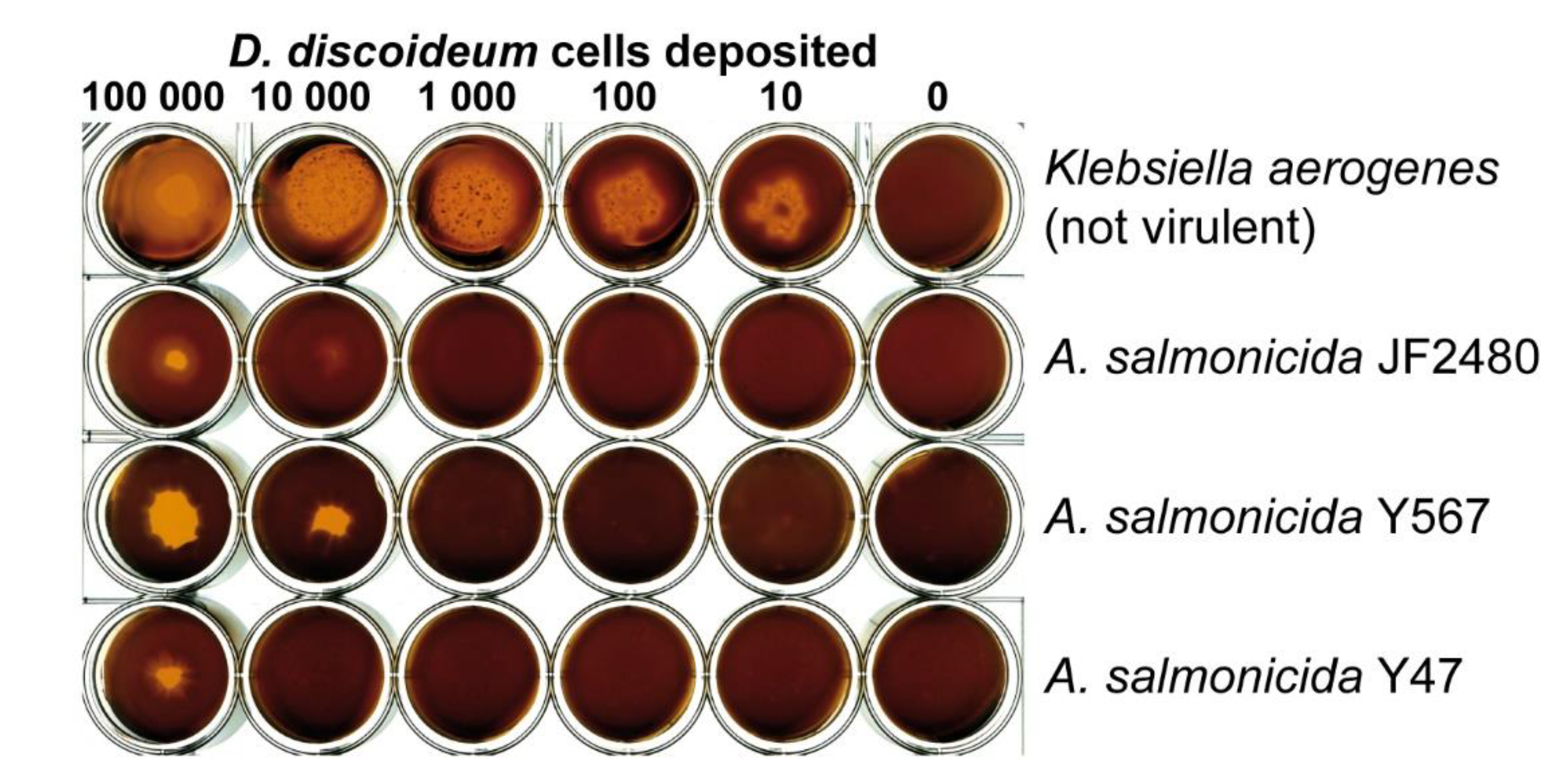
2.4. Virulence Assay
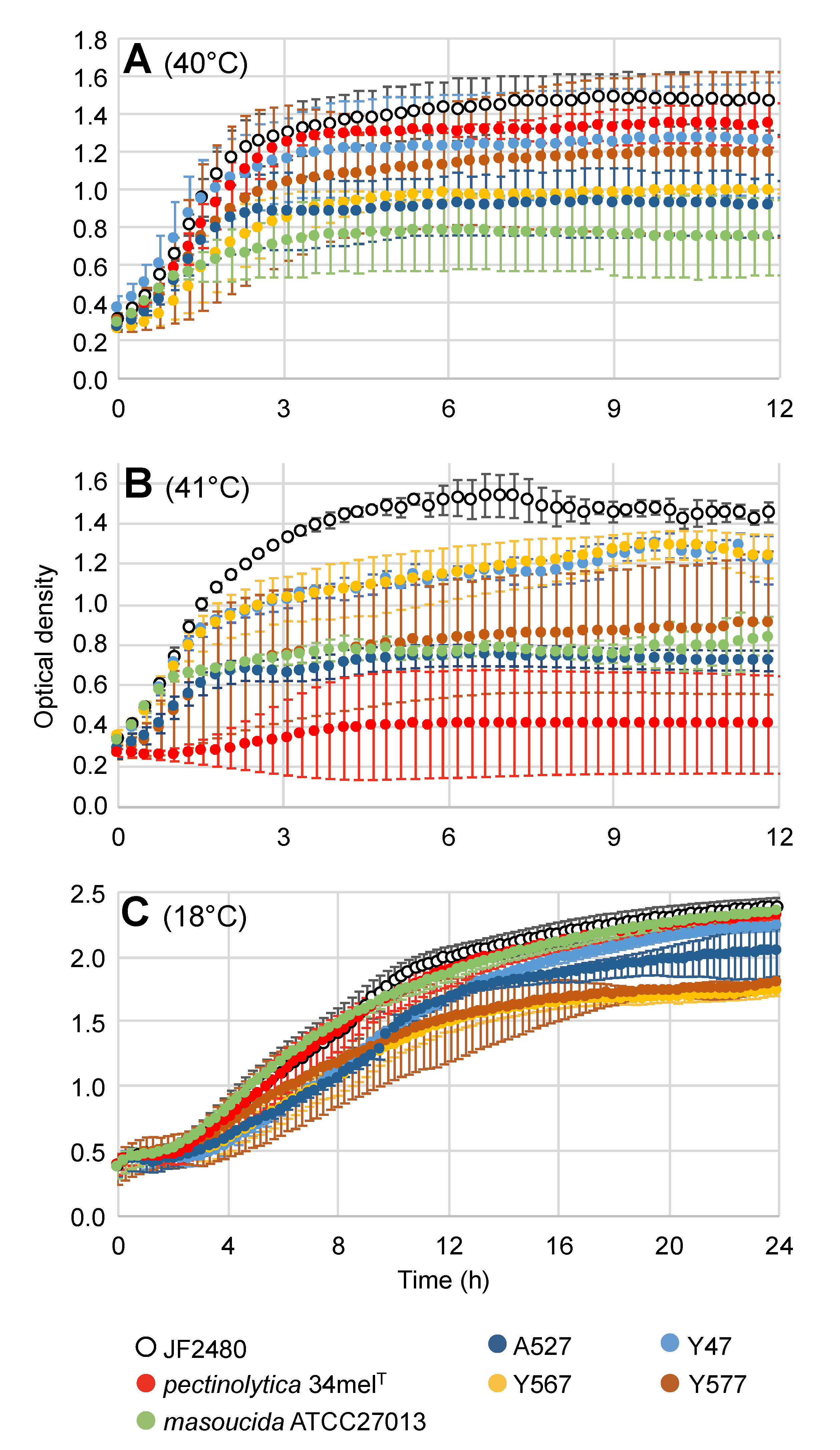
2.5. Growth Curve
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Martin-Carnahan, A.; Joseph, S. Aeromonadales ord. nov. In B, Bergey’s Manual of Systematic Bacteriology SE - 12; Brenner, D., Krieg, N., Staley, J., Garrity, G., Boone, D., De Vos, P., Goodfellow, M., Rainey, F., Schleifer, K.-H., Eds.; Springer US: New York, NY, USA, 2005. [Google Scholar]
- Dallaire-Dufresne, S.; Tanaka, K.H.; Trudel, M.V.; Lafaille, A.; Charette, S.J. Virulence, genomic features, and plasticity of Aeromonas salmonicida subsp. salmonicida, the causative agent of fish furunculosis. Vet. Microbiol. 2014, 169, 1–7. [Google Scholar]
- Austin, B.; Austin, D.A. Aeromonadaceae Representative (Aeromonas salmonicida). In Bacterial Fish Pathogens: Disease of Farmed and Wild Fish; Austin, B., Austin, D.A., Eds.; Springer International Publishing: New York, NY, USA, 2016. [Google Scholar]
- Pavan, M.E.; Abbott, S.L.; Zorzopulos, J.; Janda, J.M. Aeromonas salmonicida subsp. pectinolytica subsp. nov., a new pectinase-positive subspecies isolated from a heavily polluted river. Int. J. Syst. Evol. Microbiol. 2000, 50, 1119–1124. [Google Scholar] [PubMed]
- Abbott, S.L.; Cheung, W.K.; Kroske-Bystrom, S.; Malekzadeh, T.; Janda, J.M. Identification of Aeromonas strains to the genospecies level in the clinical laboratory. J. Clin. Microbiol. 1992, 30, 1262–1266. [Google Scholar]
- Altwegg, M.; Steigerwalt, A.G.; Altwegg-Bissig, R.; Luthy-Hottenstein, J.; Brenner, D.J. Biochemical identification of Aeromonas genospecies isolated from humans. J. Clin. Microbiol. 1990, 28, 258–264. [Google Scholar]
- Janda, J.M.; Abbott, S.L.; Khashe, S.; Kellogg, G.H.; Shimada, T. Further studies on biochemical characteristics and serologic properties of the genus Aeromonas. J. Clin. Microbiol. 1996, 34, 1930–1933. [Google Scholar]
- Janda, J.M.; Abbott, S.L. The genus Aeromonas: Taxonomy, pathogenicity, and infection. Clin. Microbiol. Rev. 2010, 23, 35–73. [Google Scholar] [CrossRef]
- Beaz-Hidalgo, R.; Alperi, A.; Bujan, N.; Romalde, J.L.; Figueras, M.J. Comparison of phenotypical and genetic identification of Aeromonas strains isolated from diseased fish. Syst. Appl. Microbiol. 2010, 33, 149–153. [Google Scholar] [CrossRef]
- Vincent, A.T.; Trudel, M.V.; Freschi, L.; Nagar, V.; Gagne-Thivierge, C.; Levesque, R.C.; Charette, S.J. Increasing genomic diversity and evidence of constrained lifestyle evolution due to insertion sequences in Aeromonas salmonicida. BMC Genom. 2016, 17, 44. [Google Scholar] [CrossRef]
- Vincent, A.T.; Rouleau, F.D.; Moineau, S.; Charette, S.J. Study of mesophilic Aeromonas salmonicida A527 strain sheds light on the species’ lifestyles and taxonomic dilemma. FEMS Microbiol. Lett. 2017, 364. [Google Scholar] [CrossRef]
- Nagar, V.; Shashidhar, R.; Bandekar, J.R. Prevalence, characterization, and antimicrobial resistance of Aeromonas strains from various retail food products in Mumbai, India. J. Food Sci. 2011, 76, M486–M492. [Google Scholar] [CrossRef]
- Kamble, R. Aeromonas salmonicida furunculosis in an adult male. Int. J. Curr. Microbiol. App. Sci. 2015, 4, 59–63. [Google Scholar]
- Ruppe, E.; Cherkaoui, A.; Wagner, N.; La Scala, G.C.; Beaulieu, J.Y.; Girard, M.; Frey, J.; Lazarevic, V.; Schrenzel, J. In vivo selection of a multidrug-resistant Aeromonas salmonicida during medicinal leech therapy. New Microbes New Infect. 2018, 21, 23–27. [Google Scholar] [CrossRef] [PubMed]
- Tewari, R.; Dudeja, M.; Nandy, S.; Das, A.K. Isolation of Aeromonas salmonicida from human blood sample: A case report. J. Clin. Diagn. Res. 2014, 8, 139–140. [Google Scholar] [CrossRef]
- Varshney, A.; Das, M.; Chaudhary, P.; Kumari, R.; Yadav, K. Aeromonas salmonicida as a causative agent for postoperative endophthalmitis. Middle East Afr. J. Ophthalmol. 2017, 24, 213–215. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Yang, Q.Q.; Guo, Q.Y.; Yi, C.Y.; Mao, H.P.; Lin, J.X.; Jiang, Z.P.; Yu, X.Q. Aeromonas salmonicida peritonitis after eating fish in a patient undergoing CAPD. Perit. Dial. Int. 2008, 28, 316–317. [Google Scholar]
- Vincent, A.T.; Fernandez-Bravo, A.; Sanchis, M.; Mayayo, E.; Figueras, M.J.; Charette, S.J. Investigation of the virulence and genomics of Aeromonas salmonicida strains isolated from human patients. J. Mol. Epidemiol. Evol. Genet. Infect. Dis. 2019, 68, 1–9. [Google Scholar] [CrossRef]
- Coil, D.; Jospin, G.; Darling, A.E. A5-miseq: An updated pipeline to assemble microbial genomes from Illumina MiSeq data. Bioinformatics 2015, 31, 587–589. [Google Scholar] [CrossRef]
- Tatusova, T.; DiCuccio, M.; Badretdin, A.; Chetvernin, V.; Nawrocki, E.P.; Zaslavsky, L.; Lomsadze, A.; Pruitt, K.D.; Borodovsky, M.; Ostell, J. NCBI prokaryotic genome annotation pipeline. Nucleic Acids Res. 2016, 44, 6614–6624. [Google Scholar] [CrossRef]
- Seemann, T. Prokka: Rapid prokaryotic genome annotation. Bioinformatics 2014, 30, 2068–2069. [Google Scholar] [CrossRef]
- Kristensen, D.M.; Kannan, L.; Coleman, M.K.; Wolf, Y.I.; Sorokin, A.; Koonin, E.V.; Mushegian, A. A low-polynomial algorithm for assembling clusters of orthologous groups from intergenomic symmetric best matches. Bioinformatics 2010, 26, 1481–1487. [Google Scholar] [CrossRef]
- Li, L.; Stoeckert, C.J., Jr.; Roos, D.S. OrthoMCL: Identification of ortholog groups for eukaryotic genomes. Genome Res. 2003, 13, 2178–2189. [Google Scholar] [CrossRef] [PubMed]
- Contreras-Moreira, B.; Vinuesa, P. GET_HOMOLOGUES, a versatile software package for scalable and robust microbial pangenome analysis. Appl. Environ. Microbiol. 2013, 79, 7696–7701. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [PubMed]
- Abascal, F.; Zardoya, R.; Telford, M.J. TranslatorX: Multiple alignment of nucleotide sequences guided by amino acid translations. Nucleic acids Res. 2010, 38, W7–W13. [Google Scholar] [CrossRef] [PubMed]
- Criscuolo, A.; Gribaldo, S. BMGE (Block Mapping and Gathering with Entropy): A new software for selection of phylogenetic informative regions from multiple sequence alignments. BMC Evol. Biol. 2010, 10, 210. [Google Scholar] [CrossRef] [PubMed]
- Borowiec, M.L. AMAS: A fast tool for alignment manipulation and computing of summary statistics. PeerJ 2016, 4, e1660. [Google Scholar] [CrossRef]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.F.; von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast model selection for accurate phylogenetic estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef]
- Nguyen, L.T.; Schmidt, H.A.; von Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum-likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef]
- Hoang, D.T.; Chernomor, O.; von Haeseler, A.; Minh, B.Q.; Vinh, L.S. UFBoot2: Improving the ultrafast bootstrap approximation. Mol. Biol. Evol. 2018, 35, 518–522. [Google Scholar] [CrossRef]
- Wattam, A.R.; Davis, J.J.; Assaf, R.; Boisvert, S.; Brettin, T.; Bun, C.; Conrad, N.; Dietrich, E.M.; Disz, T.; Gabbard, J.L.; et al. Improvements to PATRIC, the all-bacterial bioinformatics database and analysis resource center. Nucleic Acids Res. 2017, 45, D535–D542. [Google Scholar] [CrossRef]
- Abby, S.S.; Rocha, E.P.C. Identification of protein secretion systems in bacterial genomes using macsyfinder. Methods Mol. Biol. 2017, 1615, 1–21. [Google Scholar] [PubMed]
- Huerta-Cepas, J.; Forslund, K.; Coelho, L.P.; Szklarczyk, D.; Jensen, L.J.; von Mering, C.; Bork, P. Fast Genome-Wide Functional Annotation through Orthology Assignment by eggNOG-Mapper. Mol. Biol. Evol. 2017, 34, 2115–2122. [Google Scholar] [CrossRef] [PubMed]
- Froquet, R.; Cherix, N.; Burr, S.E.; Frey, J.; Vilches, S.; Tomas, J.M.; Cosson, P. Alternative host model to evaluate Aeromonas virulence. Appl. Environ. Microbiol. 2007, 73, 5657–5659. [Google Scholar] [CrossRef] [PubMed]
- Cornillon, S.; Pech, E.; Benghezal, M.; Ravanel, K.; Gaynor, E.; Letourneur, F.; Bruckert, F.; Cosson, P. Phg1p is a nine-transmembrane protein superfamily member involved in dictyostelium adhesion and phagocytosis. J. Biol. Chem. 2000, 275, 34287–34292. [Google Scholar] [CrossRef]
- Mercanti, V.; Charette, S.J.; Bennett, N.; Ryckewaert, J.J.; Letourneur, F.; Cosson, P. Selective membrane exclusion in phagocytic and macropinocytic cups. J. Cell Sci. 2006, 119, 4079–4087. [Google Scholar] [CrossRef]
- Baldwin, T.; Sakthianandeswaren, A.; Curtis, J.M.; Kumar, B.; Smyth, G.K.; Foote, S.J.; Handman, E. Wound healing response is a major contributor to the severity of cutaneous leishmaniasis in the ear model of infection. Parasite Immunol. 2007, 29, 501–513. [Google Scholar] [CrossRef]
- Joest, R. Junge Säbelschnäbler (Recurvirostra avosetta L.) in unterschiedlichen Klimazonen: Physiologische und ethologische Anpassungen an ökologische Bedingungen in Norddeutschland und Südspanien; Christian-Albrechts Universität Kiel: Kiel, Germany, 2003. [Google Scholar]
- Vincent, A.T.; Le Breton, A.; Bernatchez, A.; Gagne-Thivierge, C.; Paquet, V.E.; Thibault, E.; Charette, S.J.; Gantelet, H. Draft genome sequences of four Aeromonas salmonicida subsp. achromogenes Strains, 23051, 23053, 23055, and 23056, Isolated from senegalese sole (Solea senegalensis). Microbiol. Resour. Announc. 2019, 8. [Google Scholar] [CrossRef] [Green Version]
- Trudel, M.V.; Vincent, A.T.; Attere, S.A.; Labbe, M.; Derome, N.; Culley, A.I.; Charette, S.J. Diversity of antibiotic-resistance genes in Canadian isolates of Aeromonas salmonicida subsp. salmonicida: Dominance of pSN254b and discovery of pAsa8. Sci. Rep. 2016, 6, 35617. [Google Scholar] [CrossRef] [Green Version]
- Vincent, A.T.; Trudel, M.V.; Paquet, V.E.; Boyle, B.; Tanaka, K.H.; Dallaire-Dufresne, S.; Daher, R.K.; Frenette, M.; Derome, N.; Charette, S.J. Detection of variants of the pRAS3, pAB5S9, and pSN254 plasmids in Aeromonas salmonicida subsp. salmonicida: Multidrug resistance, interspecies exchanges, and plasmid reshaping. Antimicrob. Agents Chemother. 2014, 58, 7367–7374. [Google Scholar] [CrossRef] [Green Version]
- Massicotte, M.A.; Vincent, A.T.; Schneider, A.; Paquet, V.E.; Frenette, M.; Charette, S.J. One Aeromonas salmonicida subsp. salmonicida isolate with a pAsa5 variant bearing antibiotic resistance and a pRAS3 variant making a link with a swine pathogen. Sci. Total. Environ. 2019, 690, 313–320. [Google Scholar] [CrossRef]
- Costa, T.R.; Felisberto-Rodrigues, C.; Meir, A.; Prevost, M.S.; Redzej, A.; Trokter, M.; Waksman, G. Secretion systems in Gram-negative bacteria: Structural and mechanistic insights. Nat. Rev. Microbiol. 2015, 13, 343–359. [Google Scholar] [CrossRef] [PubMed]
- Daher, R.K.; Filion, G.; Tan, S.G.; Dallaire-Dufresne, S.; Paquet, V.E.; Charette, S.J. Alteration of virulence factors and rearrangement of pAsa5 plasmid caused by the growth of Aeromonas salmonicida in stressful conditions. Vet. Microbiol. 2011, 152, 353–360. [Google Scholar] [CrossRef] [PubMed]
- Froquet, R.; Lelong, E.; Marchetti, A.; Cosson, P. Dictyostelium discoideum: A model host to measure bacterial virulence. Nat. Protoc. 2009, 4, 25–30. [Google Scholar] [CrossRef] [PubMed]
- Studer, N.; Frey, J.; Vanden Bergh, P. Clustering subspecies of Aeromonas salmonicida using IS630 typing. BMC Microbiol. 2013, 13, 36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeng, W.B.; Chen, W.B.; Yan, Q.P.; Lin, G.F.; Qin, Y.X. Hemerythrin is required for Aeromonas hydraphlia to survive in the macrophages of Anguilla japonica. Genet. Mol. Res: GMR 2016, 15. [Google Scholar] [CrossRef]
- Eshghi, A.; Lourdault, K.; Murray, G.L.; Bartpho, T.; Sermswan, R.W.; Picardeau, M.; Adler, B.; Snarr, B.; Zuerner, R.L.; Cameron, C.E. Leptospira interrogans catalase is required for resistance to H2O2 and for virulence. Infect. Immun. 2012, 80, 3892–3899. [Google Scholar] [CrossRef] [Green Version]
- Waldor, M.K.; Mekalanos, J.J. Lysogenic conversion by a filamentous phage encoding cholera toxin. Science 1996, 272, 1910–1914. [Google Scholar] [CrossRef] [Green Version]
- Martin-Carnahan, A.; Joseph, S.W. Aeromonas. In Bergey’s Manual of Systematics of Archaea and Bacteria; Whitman, W.B., Rainey, F., Kämpfer, P., Trujillo, M., Chun, J., DeVos, P., Hedlund, B., Dedysh, S., Eds.; Wiley: Hoboken, NJ, USA, 2015; pp. 1–44. [Google Scholar] [CrossRef]
- Frey, J.; Origgi, F.C. Type III secretion system of aeromonas salmonicida undermining the host’s immune response. Front. Mar. Sci. 2016, 3. [Google Scholar] [CrossRef]
- Fehr, D.; Burr, S.E.; Gibert, M.; d’Alayer, J.; Frey, J.; Popoff, M.R. Aeromonas exoenzyme T of Aeromonas salmonicida is a bifunctional protein that targets the host cytoskeleton. J. Biol. Chem. 2007, 282, 28843–28852. [Google Scholar] [CrossRef] [Green Version]
- Ding, Y.; Liu, X.; Chen, F.; Di, H.; Xu, B.; Zhou, L.; Deng, X.; Wu, M.; Yang, C.G.; Lan, L. Metabolic sensor governing bacterial virulence in Staphylococcus aureus. Proc. Natl. Acad. Sci. USA 2014, 111, E4981–E4990. [Google Scholar] [CrossRef] [Green Version]
- Minato, Y.; Fassio, S.R.; Wolfe, A.J.; Hase, C.C. Central metabolism controls transcription of a virulence gene regulator in Vibrio cholerae. Microbiology 2013, 159, 792–802. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arora, G.; Chaudhary, D.; Kidwai, S.; Sharma, D.; Singh, R. Cite enzymes are essential for mycobacterium tuberculosis to establish infection in macrophages and guinea pigs. Front. Cell. Infect. Microbiol. 2018, 8, 385. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Subspecies | Lifestyle | Origin | Reference |
|---|---|---|---|---|
| JF2480 | N/A | Mesophile | Switzerland | This study |
| Y47 | N/A | Mesophile | India | [10] |
| Y567 | N/A | Mesophile | India | [10] |
| Y577 | N/A | Mesophile | India | [10] |
| A527 | N/A | Mesophile | India | [11] |
| 34 mel T | pectinolytica | Mesophile | Argentina | [4] |
| NBRC 13784 T | masoucida | Psychrophile | Japan | [19] |
| Protein | Virulence Trait | Strainsc | ||
|---|---|---|---|---|
| JF2480 | Y567 | Y47 | ||
| Two pore domain potassium channel family protein | N/A a | 40% d | 99% | 99% |
| Hemerythrin | A. hydrophila survival in host macrophages | 98% | 98% | 98% |
| Pseudaminic acid cytidylyltransferase | Colonisation of H. pylori | 99% | 50%d | 100% |
| Catalase KatE b | Virulence of Leptospira spp. in animal models | 98% | 99% | 98% |
| UDP-N-acetylglucosamine-1-phosphate transferase c | Production of enterobacterial antigen in S. enterica | 45% d | Absent | 61% d |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vincent, A.T.; Bernatchez, A.; Frey, J.; Charette, S.J. A Mesophilic Aeromonas salmonicida Strain Isolated from an Unsuspected Host, the Migratory Bird Pied Avocet. Microorganisms 2019, 7, 592. https://doi.org/10.3390/microorganisms7120592
Vincent AT, Bernatchez A, Frey J, Charette SJ. A Mesophilic Aeromonas salmonicida Strain Isolated from an Unsuspected Host, the Migratory Bird Pied Avocet. Microorganisms. 2019; 7(12):592. https://doi.org/10.3390/microorganisms7120592
Chicago/Turabian StyleVincent, Antony T., Alex Bernatchez, Joachim Frey, and Steve J. Charette. 2019. "A Mesophilic Aeromonas salmonicida Strain Isolated from an Unsuspected Host, the Migratory Bird Pied Avocet" Microorganisms 7, no. 12: 592. https://doi.org/10.3390/microorganisms7120592
APA StyleVincent, A. T., Bernatchez, A., Frey, J., & Charette, S. J. (2019). A Mesophilic Aeromonas salmonicida Strain Isolated from an Unsuspected Host, the Migratory Bird Pied Avocet. Microorganisms, 7(12), 592. https://doi.org/10.3390/microorganisms7120592