An Update on the Genus Aeromonas: Taxonomy, Epidemiology, and Pathogenicity

Abstract
:1. Introduction
2. Historic Perspective: Past and Present
3. Taxonomy and Identification Methods
3.1. Phenotypic Identification
3.2. Molecular Identification
3.2.1. Techniques Based on the 16S rRNA Gene
3.2.2. Housekeeping Genes
3.2.3. Genotyping Methods
3.2.4. Genomics
4. Ecology and Epidemiology
4.1. Aeromonas in Aquatic Environments
4.2. Aeromonas in Food
4.3. Aeromonas in Animals
4.4. Aeromonas in Human Infections
4.5. Gastroenteritis
4.5.1. Wound Infections
4.5.2. Bacteremia/Septicemia
4.5.3. Other Infections
5. Virulence Factors
5.1. Structural Components
5.2. Extracellular Proteins
5.3. Secretion Systems
5.4. Quorum Sensing
5.5. Metal Ions
6. Host–Pathogen Interaction
7. Pathogenicity Studies
7.1. In Vitro Studies
7.2. Animal Models
8. Antimicrobial Resistance
9. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Martin-Carnahan, A.; Joseph, S.W. Order XII. Aeromonadales ord. nov. In Bergey’s Manual of Systematic Bacteriology; Brenner, D.J., Krieg, N.R., Staley, J.T., Garrity, G.M., Eds.; Williams & Wilkins: Philadelphia, PA, USA, 2005; pp. 556–578. [Google Scholar]
- Martínez-Murcia, A.; Beaz-Hidalgo, R.; Navarro, A.; Carvalho, M.J.; Aravena-Román, M.; Correia, A.; Figueras, M.J.; Saavedra, M.J. Aeromonas lusitana sp. nov., isolated from untreated water and vegetables. Curr. Microbiol. 2016, 72, 795–803. [Google Scholar] [CrossRef]
- Figueras, M.J.; Suarez-Franquet, A.; Chacon, M.R.; Soler, L.; Navarro, M.; Alejandre, C.; Grasa, B.; Martínez-Murcia, A.J.; Guarro, J. First record of the rare species Aeromonas culicicola from a drinking water supply. Appl. Environ. Microbiol. 2005, 71, 538–541. [Google Scholar] [CrossRef] [Green Version]
- Janda, J.M.; Abbott, S.L. The genus Aeromonas: Taxonomy, pathogenicity, and infection. Clin. Microbiol. Rev. 2010, 23, 35–73. [Google Scholar] [CrossRef] [Green Version]
- Figueras, M.J.; Beaz-Hidalgo, R. Aeromonas infections in humans. In Aeromonas; Academic Press: Norfolk, UK, 2015; pp. 65–108. [Google Scholar]
- Salvat, M.J.F.; Ashbolt, N. Aeromonas . In Global Water Pathogen Project; University of Alberta: Edmonton, AB, Canada, 2019. [Google Scholar]
- Beaz-Hidalgo, R.; Alperi, A.; Buján, N.; Romalde, J.L.; Figueras, M.J. Comparison of phenotypical and genetic identification of Aeromonas strains isolated from diseased fish. Syst. Appl. Microbiol. 2010, 33, 149–153. [Google Scholar] [CrossRef]
- Martínez-Murcia, A.J.; Benlloch, S.; Collins, M.D. Phylogenetic interrelationships of members of the genera Aeromonas and Plesiomonas as determined by 16S ribosomal DNA sequencing: Lack of congruence with results of DNA-DNA hybridizations. Int. J. Syst. Bacteriol. 1992, 42, 412–421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yáñez, M.A.; Catalan, V.; Apráiz, D.; Figueras, M.J.; Martínez-Murcia, A.J. Phylogenetic analysis of members of the genus Aeromonas based on gyrB gene sequences. Int. J. Syst. Evol. Microbiol. 2003, 53, 875–883. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soler, L.; Yáñez, M.A.; Chacon, M.R.; Aguilera-Arreola, M.G.; Catalan, V.; Figueras, M.J.; Martínez-Murcia, A.J. Phylogenetic analysis of the genus Aeromonas based on two housekeeping genes. Int. J. Syst. Evol. Microbiol. 2004, 54, 1511–1519. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Murcia, A.J.; Monera, A.; Saavedra, M.J.; Oncina, R.; Lopez-Alvarez, M.; Lara, E.; Figueras, M.J. Multilocus phylogenetic analysis of the genus Aeromonas. Syst. Appl. Microbiol. 2011, 34, 189–199. [Google Scholar] [CrossRef] [PubMed]
- Beaz-Hidalgo, R.; Hossain, M.J.; Liles, M.R.; Figueras, M.-J. Strategies to avoid wrongly labelled genomes using as example the detected wrong taxonomic affiliation for Aeromonas genomes in the genbank database. PLoS ONE 2015, 10. [Google Scholar] [CrossRef]
- Beaz-Hidalgo, R.; Laviad-Shitrit, S.; Shakèd, T.; Halpern, M.; Figueras, M.J. Chironomid egg masses harbour the clinical species Aeromonas taiwanensis and Aeromonas sanarellii. FEMS Microbiol. Lett. 2012, 337, 48–54. [Google Scholar] [CrossRef]
- Figueras, M.J.; Beaz-Hidalgo, R.; Hossain, M.J.; Liles, M.R. Taxonomic affiliation of new genomes should be verified using average nucleotide identity and multilocus phylogenetic analysis. Genome Announc. 2014, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Latif-Eugenin, F.L. Aeromonas, un microorganismo ambiental de importancia en salud humana y animal; Universidad Rovira i Virgili: Tarragona, Spain, 2015. [Google Scholar]
- Vincent, A.T.; Fernández-Bravo, A.; Sanchis, M.; Mayayo, E.; Figueras, M.J.; Charette, S.J. Investigation of the virulence and genomics of Aeromonas salmonicida strains isolated from human patients. Infect. Genet. Evol. 2019, 68, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Romero, A.; Saraceni, P.R.; Merino, S.; Figueras, A.; Tomás, J.M.; Novoa, B. The animal model determines the results of Aeromonas virulence factors. Front. Microbiol. 2016, 7, 1245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tomás, J.M. The main Aeromonas pathogenic factors. ISRN Microbiol. 2012, 2012, 1–22. [Google Scholar] [CrossRef] [Green Version]
- Beaz-Hidalgo, R.; Figueras, M.J. Aeromonas spp. whole genomes and virulence factors implicated in fish disease. J. Fish Dis. 2013, 36, 371–388. [Google Scholar] [CrossRef]
- Fernández-Bravo, A. Epidemiology and pathogenic characterization of species of the genus Aeromonas; Universidad Rovira i Virgili: Tarragona, Spain, 2019. [Google Scholar]
- Su, J.; Yang, C.; Xiong, F.; Wang, Y.; Zhu, Z. Toll-like receptor 4 signaling pathway can be triggered by grass carp reovirus and Aeromonas hydrophila infection in rare minnow Gobiocypris rarus. Fish Shellfish. Immunol. 2009, 27, 33–39. [Google Scholar] [CrossRef]
- McCoy, A.J.; Koizumi, Y.; Higa, N.; Suzuki, T. Differential regulation of caspase-1 activation via NLRP3/NLRC4 inflammasomes mediated by aerolysin and type III secretion system during Aeromonas veronii infection. J. Immunol. 2010, 185, 7077–7084. [Google Scholar] [CrossRef] [Green Version]
- McCoy, A.J.; Koizumi, Y.; Toma, C.; Higa, N.; Dixit, V.; Taniguchi, S.; Tschopp, J.; Suzuki, T. Cytotoxins of the human pathogen Aeromonas hydrophila trigger, via the NLRP3 inflammasome, caspase-1 activation in macrophages. Eur. J. Immunol. 2010, 40, 2797–2803. [Google Scholar] [CrossRef]
- Srivastava, N.; Shelly, A.; Kumar, M.; Pant, A.; Das, B.; Majumdar, T.; Mazumder, S. Aeromonas hydrophila utilizes TLR4 topology for synchronous activation of MyD88 and TRIF to orchestrate anti-inflammatory responses in zebrafish. Cell Death Discov. 2017, 3, 17067. [Google Scholar] [CrossRef] [Green Version]
- Sanarelli, G. Ueber einen neuen mikroorganismus des Wassers, welcher fur thiere mit veraenderlicher und konstanter temperatur pathogen ist. Zentrable Bakteriol. Parasitenkd. Infekt. Hyg. Abt. 1991, 2, 222–228. [Google Scholar]
- Colwell, R.R.; McDonell, M.R.; Ley, D.J. Proposal to recognise the family Aeromonadaceae fam. nov. Int. J. Med. Microbiol. 1986, 26, 19–27. [Google Scholar]
- Tanaka, K.H.; Dallaire-Dufresne, S.; Daher, R.K.; Frenette, M.; Charette, S.J. An insertion sequence-dependent plasmid rearrangement in Aeromonas salmonicida causes the loss of the type three secretion system. PLoS ONE 2012, 7, e33725. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanaka, K.H.; Vincent, A.T.; Emond-Rheault, J.-G.; Adamczuk, M.; Frenette, M.; Charette, S.J. Plasmid composition in Aeromonas salmonicida subsp. salmonicida 01-B526 unravels unsuspected type three secretion system loss patterns. BMC Genom. 2017, 18, 528. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martinez-Murcia, A.; Garay, E.; Collins, M.; Esteve, C. Aeromonas allosaccharophila sp. nov., a new mesophilic member of the genus Aeromonas. FEMS Microbiol. Lett. 1992, 91, 199–205. [Google Scholar] [CrossRef]
- Beaz-Hidalgo, R.; Latif-Eugenin, F.; Hossain, M.; Berg, K.; Niemi, R.; Rapala, J.; Lyra, C.; Liles, M.; Figueras, M.J. Aeromonas aquatica sp. nov., Aeromonas finlandiensis sp. nov. and Aeromonas lacus sp. nov. isolated from Finnish waters associated with cyanobacterial blooms. Syst. Appl. Microbiol. 2015, 38, 161–168. [Google Scholar] [CrossRef]
- Figueras, M.J.; Latif-Eugenin, F.; Ballester, F.; Pujol, I.; Tena, D.; Berg, K.; Hossain, M.J.; Beaz-Hidalgo, R.; Liles, M.R. ‘Aeromonas intestinalis’ and ‘Aeromonas enterica’ isolated from human faeces, ‘Aeromonas crassostreae’ from oyster and ‘Aeromonas aquatilis’ isolated from lake water represent novel species. New Microbes New Infect. 2017, 15, 74–76. [Google Scholar] [CrossRef]
- Aravena-Román, M.; Beaz-Hidalgo, R.; Inglis, T.J.J.; Riley, T.V.; Martínez-Murcia, A.J.; Chang, B.J.; Figueras, M.J. Aeromonas australiensis sp. nov., isolated from irrigation water. Int. J. Syst. Evol. Microbiol. 2013, 63, 3130. [Google Scholar] [CrossRef]
- Ali, A.; Carnahan, A.; Altwegg, M.; Luthy-Hottenstein, J.; Josep, S.W. Aeromonas bestiarum sp. nov. (formely genomospecies DNA group 2 Aeromonas hydrophila), a new species isolated from non human sources. Med. Microbiol. Lett. 1996, 5, 156–165. [Google Scholar]
- Miñana-Galbis, D.; Farfán, M.; Fusté, M.C.; Lorén, J.G. Aeromonas bivalvium sp. nov., isolated from bivalve molluscs. Int. J. Syst. Evol. Microbiol. 2007, 57, 582–587. [Google Scholar] [CrossRef]
- Martinez-Murcia, A.; Beaz-Hidalgo, R.; Svec, P.; Saavedra, M.J.; Figueras, M.J.; Sedlacek, I. Aeromonas cavernicola sp. nov., isolated from fresh water of a brook in a cavern. Curr. Microbiol. 2013, 66, 197–204. [Google Scholar] [CrossRef]
- Schubert, R.H.; Hegazi, M. Aeromonas eucrenophila species nova Aeromonas caviae a later and illegitimate synonym of Aeromonas punctata. Zent. Bakteriol. Mikrobiol. Hyg. Ser. A Med. Microbiol. Infect. Dis. Virol. Parasitol. 1988, 268, 34–39. [Google Scholar] [CrossRef]
- Beaz-Hidalgo, R.; Martinez-Murcia, A.; Figueras, M.J. Reclassification of Aeromonas hydrophila subsp. dhakensis Huys et al. 2002 and Aeromonas aquariorum Martinez-Murcia et al. 2008 as Aeromonas dhakensis sp. nov. comb nov. and emendation of the species Aeromonas hydrophila. Syst. Appl. Microbiol. 2013, 36, 171–176. [Google Scholar] [CrossRef] [PubMed]
- Miñana-Galbis, D.; Farfán, M.; Lorén, J.G.; Fusté, M.C. Proposal to assign Aeromonas diversa sp. nov. as a novel species designation for Aeromonas group 501. Syst. Appl. Microbiol. 2010, 33, 15–19. [Google Scholar] [CrossRef] [PubMed]
- Esteve, C.; Gutierrez, M.C.; Ventosa, A.; Verdonck, L.; Mergaert, J.; Rijckaert, C.; Swings, J.; Kersters, K.; De Ley, J. Aeromonas encheleia sp. nov., isolated from European eels. Int. J. Syst. Bacteriol. 1995, 45, 462–466. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alperi, A.; Martínez-Murcia, A.J.; Monera, A.; Saavedra, M.J.; Figueras, M.J. Aeromonas fluvialis sp. nov., isolated from a Spanish river. Int. J. Syst. Evol. Microbiol. 2010, 60, 1008. [Google Scholar] [CrossRef] [Green Version]
- Stanier, R.Y. A note on the taxonomy of P. hydrophilus. J. Bacteriol. 1943, 46, 213–214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carnahan, A.M.; Chakraborty, T.; Fanning, G.R.; Verma, D.; Ali, A.; Janda, J.M.; Joseph, S.W. Aeromonas trota sp. nov., an ampicillin-susceptible species isolated from clinical specimens. J. Clin. Microbiol. 1991, 29, 1206–1210. [Google Scholar] [CrossRef] [Green Version]
- Allen, D.A.; Austin, B.; Colwell, R.R.; Gavini, F.; Ferragut, C.; Izard, D.; Trinel, P.A.; Leclerc, H.; Lefebvre, B.; Mossel, D.A.A. Aeromonas media, a new species isolated from river water. Int. J. Syst. Bacteriol. 1983, 33, 599–604. [Google Scholar] [CrossRef] [Green Version]
- Miñana-Galbis, D. Aeromonas molluscorum sp. nov., isolated from bivalve molluscs. Int. J. Syst. Evol. Microbiol. 2004, 54, 2073–2078. [Google Scholar] [CrossRef]
- Beaz-Hidalgo, R.; Alperi, A.; Figueras, M.J.; Romalde, J.L. Aeromonas piscicola sp. nov., isolated from diseased fish. Syst. Appl. Microbiol. 2009, 32, 471–479. [Google Scholar] [CrossRef]
- Huys, G.; Kampfer, P.; Altwegg, M.; Kersters, I.; Lamb, A.; Coopman, R.; Lüthy-Hottenstein, J.; Vancanneyt, M.; Janssen, P.; Kersters, K. Aeromonas popoffii sp. nov., a mesophilic bacterium isolated from drinking water production plants and reservoirs. Int. J. Syst. Bacteriol. 1997, 47, 1165–1171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marti, E.; Balcázar, J.L. Aeromonas rivipollensis sp. nov., a novel species isolated from aquatic samples. J. Basic Microbiol. 2015, 55, 1435–1439. [Google Scholar] [CrossRef] [PubMed]
- Figueras, M.J.; Alperi, A.; Beaz-Hidalgo, R.; Stackebrandt, E.; Brambilla, E.; Monera, A.; Martinez-Murcia, A.J. Aeromonas rivuli sp. nov., isolated from the upstream region of a karst water rivulet. Int. J. Syst. Evol. Microbiol. 2011, 61, 242–248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Griffin, P.J.; Snieszko, S.F.; Friddle, S.B. Pigment formation by Bacterium salmonicida. J. Bacteriol. 1953, 65, 652–659. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alperi, A.; Martinez-Murcia, A.J.; Ko, W.C.; Monera, A.; Saavedra, M.J.; Figueras, M.J. Aeromonas taiwanensis sp. nov. and Aeromonas sanarellii sp. nov., clinical species from Taiwan. Int. J. Syst. Evol. Microbiol. 2010, 60, 2048–2055. [Google Scholar] [CrossRef] [Green Version]
- Hickman-Brenner, F.W.; Fanning, G.R.; Arduino, M.J.; Brenner, D.J.; Farmer, J.J. Aeromonas schubertii, a new mannitol-negative species found in human clinical specimens. J. Clin. Microbiol. 1988, 26, 1561–1564. [Google Scholar] [CrossRef] [Green Version]
- Harf-Monteil, C.; Le Flèche, A.; Riegel, P.; Prévost, G.; Bermond, D.; Grimont, P.A.D.; Monteil, H. Aeromonas simiae sp. nov., isolated from monkey faeces. Int. J. Syst. Evol. Microbiol. 2004, 54, 481–485. [Google Scholar] [CrossRef]
- Popoff, M.; Veron, M. Validation of the new names and new combinations previously effectively published outside the IJSB. Int. J. Syst. Bacteriol. 1981, 31, 215. [Google Scholar]
- Demarta, A.; Küpfer, M.; Riegel, P.; Harf-Monteil, C.; Tonolla, M.; Peduzzi, R.; Monera, A.; Saavedra, M.J.; Martínez-Murcia, A. Aeromonas tecta sp. nov., isolated from clinical and environmental sources. Syst. Appl. Microbiol. 2008, 31, 278–286. [Google Scholar] [CrossRef]
- Hickman-Brenner, F.W.; Macdonald, K.L.; Steigerwalt, A.G.; Fanning, G.R.; Brenner, D.J.; Farmer, J.J. Aeromonas veronii, a new ornithine decarboxylase-positive species that may cause diarrhea. J. Clin. Microbiol. 1987, 25, 900–906. [Google Scholar] [CrossRef] [Green Version]
- Stackebrandt, E.; Garrity, G.M.; Trüper, H.G.; Whitman, W.B.; Grimont, P.A.D.; Nesme, X.; Frederiksen, W.; Vauterin, L.; Kampfer, P.; Rosselló-Mora, R.; et al. Report of the ad hoc committee for the re-evaluation of the species definition in bacteriology. Int. J. Syst. Evol. Microbiol. 2002, 52, 1043–1047. [Google Scholar] [PubMed] [Green Version]
- Figueras, M.J.; Beaz-Hidalgo, R.; Collado, L.; Martínez-Murcia, A.J. Recommendations for a new bacterial species description based on analysis of the unrelated genera Aeromonas and Arcobacter. Bull. BISMiS 2011, 2, 1–16. [Google Scholar]
- Abbott, S.L.; Cheung, W.K.W.; Janda, J.M. The genus Aeromonas: Biochemical characteristics, atypical reactions, and phenotypic identification schemes. J. Clin. Microbiol. 2003, 41, 2348–2357. [Google Scholar] [CrossRef] [Green Version]
- Lamy, B.; Laurent, F.; Verdier, I.; Decousser, J.-W.; Lecaillon, E.; Marchandin, H.; Roger, F.; Tigaud, S.; De Montclos, H.; Kodjo, A. Accuracy of 6 commercial systems for identifying clinical Aeromonas isolates. Diagn. Microbiol. Infect. Dis. 2010, 67, 9–14. [Google Scholar] [CrossRef]
- Chacón, M.R.; Castro-Escarpulli, G.; Soler, L.; Guarro, J.; Figueras, M.J. A DNA probe specific for Aeromonas colonies. Diagn. Microbiol. Infect. Dis. 2002, 44, 221–225. [Google Scholar] [CrossRef]
- Soler, L.; Marco, F.; Vilá, J.; Chacon, M.R.; Guarro, J.; Figueras, M.J. Evaluation of two miniaturized systems, MicroScan W/A and BBL Crystal E/NF, for identification of clinical isolates of Aeromonas spp. J. Clin. Microbiol. 2003, 41, 5732–5734. [Google Scholar] [CrossRef] [Green Version]
- Woo, P.; Lau, S.; Teng, J.; Tse, H.; Yuen, K.-Y. Then and now: Use of 16S rDNA gene sequencing for bacterial identification and discovery of novel bacteria in clinical microbiology laboratories. Clin. Microbiol. Infect. 2008, 14, 908–934. [Google Scholar] [CrossRef] [Green Version]
- Vega-Sánchez, V. Caracterización fenotípica y genotípica de aislamientos de Aeromonas spp. obtenidos de trucha arcoíris (Oncorhynchus mykiss); Universidad Autónoma del Estado de México: Toluca, Mexico, 2015. [Google Scholar]
- Martínez-Murcia, A.; Lamy, B. Molecular diagnostics by genetic methods. In Aeromonas; Academic Press: Norfolk, UK, 2015; pp. 155–200. [Google Scholar] [CrossRef]
- Alperi, A.; Figueras, M.J.; Inza, I.; Martínez-Murcia, A.J. Analysis of 16S rRNA gene mutations in a subset of Aeromonas strains and their impact in species delineation. Int. Microbiol. 2008, 11, 185–194. [Google Scholar]
- Roger, F.; Marchandin, H.; Jumas-Bilak, E.; Kodjo, A.; Lamy, B. Multilocus genetics to reconstruct aeromonad evolution. BMC Microbiol. 2012, 12, 62. [Google Scholar] [CrossRef] [Green Version]
- Vavrova, A.; Balazova, T.; Sedlacek, I.; Tvrzova, L.; Sedo, O. Evaluation of the MALDI-TOF MS profiling for identification of newly described Aeromonas spp. Folia Microbiol. 2015, 60, 375–383. [Google Scholar] [CrossRef]
- Chen, P.-L.; Wu, C.-J.; Chen, C.-S.; Tsai, P.-J.; Tang, H.-J.; Ko, W.-C.; Cutler, S. A comparative study of clinical Aeromonas dhakensis and Aeromonas hydrophila isolates in southern Taiwan: A. dhakensis is more predominant and virulent. Clin. Microbiol. Infect. 2014, 20, O428–O434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shin, H.B.; Yoon, J.; Lee, Y.; Kim, M.S.; Lee, K. Comparison of MALDI-TOF MS, housekeeping gene sequencing, and 16S rRNA gene sequencing for identification of Aeromonas clinical isolates. Yonsei Med. J. 2015, 56, 550–555. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pérez-Sancho, M.; Cerdá, I.; Fernández-Bravo, A.; Domínguez, L.; Figueras, M.J.; Fernández-Garayzábal, J.F.; Vela, A.I. Limited performance of MALDI-TOF for identification of fish Aeromonas isolates at species level. J. Fish Dis. 2018, 41, 1485–1493. [Google Scholar] [CrossRef] [PubMed]
- Zhong, C.; Han, M.; Yang, P.; Chen, C.; Yu, H.; Wang, L.; Ning, K. Comprehensive analysis reveals the evolution and pathogenicity of Aeromonas, viewed from both single isolated species and microbial communities. mSystems 2019, 4. [Google Scholar] [CrossRef] [Green Version]
- Harayama, S.; Kasai, H. Bacterial phylogeny reconstruction from molecular sequences. In Molecular Identification, Systematics, and Population Structure of Prokaryotes; Springer Science and Business Media: Berlin/Heidelberg, Germany, 2006; pp. 105–139. [Google Scholar]
- Küpfer, M.; Kuhnert, P.; Korczak, B.M.; Peduzzi, R.; Demarta, A. Genetic relationships of Aeromonas strains inferred from 16S rRNA, gyrB and rpoB gene sequences. Int. J. Syst. Evol. Microbiol. 2006, 56, 2743–2751. [Google Scholar] [CrossRef] [Green Version]
- Nhung, P.H.; Hata, H.; Ohkusu, K.; Noda, M.; Shah, M.M.; Goto, K.; Ezaki, T. Use of the novel phylogenetic marker dnaJ and DNA-DNA hybridization to clarify interrelationships within the genus Aeromonas. Int. J. Syst. Evol. Microbiol. 2007, 57, 1232–1237. [Google Scholar] [CrossRef] [Green Version]
- Sepe, A.; Barbieri, P.; Peduzzi, R.; Demarta, A. Evaluation of recA sequencing for the classification of Aeromonas strains at the genotype level. Lett. Appl. Microbiol. 2008, 46, 439–444. [Google Scholar] [CrossRef]
- Miñana-Galbis, D.; Urbizu-Serrano, A.; Farfán, M.; Fusté, M.C.; Lorén, J.G. Phylogenetic analysis and identification of Aeromonas species based on sequencing of the cpn60 universal target. Int. J. Syst. Evol. Microbiol. 2009, 59, 1976–1983. [Google Scholar] [CrossRef]
- Martino, M.E.; Fasolato, L.; Montemurro, F.; Rosteghin, M.; Manfrin, A.; Patarnello, T.; Novelli, E.; Cardazzo, B. Determination of microbial diversity of Aeromonas strains on the basis of multilocus sequence typing, phenotype, and presence of putative virulence genes. Appl. Environ. Microbiol. 2011, 77, 4986–5000. [Google Scholar] [CrossRef] [Green Version]
- Soler, L.; Figueras, M.J.; Chacón, M.R.; Guarro, J.; Martinez-Murcia, A.J. Comparison of three molecular methods for typing Aeromonas popoffii isolates. Antonie Leeuwenhoek 2003, 83, 341–349. [Google Scholar] [CrossRef]
- Aguilera-Arreola, M.G.; Hernandez-Rodriguez, C.; Zuniga, G.; Figueras, M.J.; Castro-Escarpulli, G.; Hernández-Rodríguez, C. Aeromonas hydrophila clinical and environmental ecotypes as revealed by genetic diversity and virulence genes. FEMS Microbiol. Lett. 2005, 242, 231–240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maiti, N.; Mandal, A.; Mohanty, S.; Mandal, R. Phenotypic and genetic characterization of Edwardsiella tarda isolated from pond sediments. Comp. Immunol. Microbiol. Infect. Dis. 2009, 32, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Fontes, M.C.; Saavedra, M.J.; Monera, A.; Martins, C.; Martínez-Murcia, A. Phylogenetic identification of Aeromonas simiae from a pig, first isolate since species description. Veter. Microbiol. 2010, 142, 313–316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Latif-Eugenín, F.; Beaz-Hidalgo, R.; Figueras, M.J. Evaluation of different conditions and culture media for the recovery of Aeromonas spp. from water and shellfish samples. J. Appl. Microbiol. 2016, 121, 883–891. [Google Scholar] [CrossRef] [PubMed]
- Jolley, K.A.; Maiden, M.C. BIGSdb: Scalable analysis of bacterial genome variation at the population level. BMC Bioinform. 2010, 11, 595. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seshadri, R.; Joseph, S.W.; Chopra, A.K.; Sha, J.; Shaw, J.; Graf, J.; Haft, D.; Wu, M.; Ren, Q.; Rosovitz, M.J.; et al. Genome sequence of Aeromonas hydrophila ATCC 7966T: Jack of all trades. J. Bacteriol. 2006, 188, 8272–8282. [Google Scholar] [CrossRef] [Green Version]
- Beatson, S.A.; das Gracas de Luna, M.; Bachmann, N.L.; Alikhan, N.F.; Hanks, K.R.; Sullivan, M.J.; Wee, B.A.; Freitas-Almeida, A.C.; Dos Santos, P.A.; de Melo, J.T.; et al. Genome sequence of the emerging pathogen Aeromonas caviae. J. Bacteriol. 2011, 193, 1286–1287. [Google Scholar] [CrossRef] [Green Version]
- Charette, S.J.; Brochu, F.; Boyle, B.; Filion, G.; Tanaka, K.H.; Derome, N. Draft genome sequence of the virulent strain 01-B526 of the fish pathogen Aeromonas salmonicida. J. Bacteriol. 2012, 194, 722–723. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Liu, Y.; Zhou, Z.; Huang, H.; Ren, Y.; Zhang, Y.; Li, G.; Zhou, Z.; Wang, L. Complete genome sequence of Aeromonas veronii strain B565. J. Bacteriol. 2011, 193, 3389–3390. [Google Scholar] [CrossRef] [Green Version]
- Reith, M.E.; Singh, R.K.; Curtis, B.; Boyd, J.M.; Bouevitch, A.; Kimball, J.; Munholland, J.; Murphy, C.; Sarty, D.; Williams, J.; et al. The genome of Aeromonas salmonicida subsp. salmonicida A449: Insights into the evolution of a fish pathogen. BMC Genom. 2008, 9, 427. [Google Scholar] [CrossRef]
- Wu, C.-J.; Wang, H.-C.; Chen, C.-S.; Shu, H.-Y.; Kao, A.-W.; Chen, P.-L.; Ko, W.-C. Genome sequence of a novel human pathogen, Aeromonas aquariorum. J. Bacteriol. 2012, 194, 4114–4115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Colston, S.M.; Fullmer, M.S.; Beka, L.; Lamy, B.; Gogarten, J.P.; Graf, J. Bioinformatic genome comparisons for taxonomic and phylogenetic assignments using Aeromonas as a test case. MBio 2014, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meier-Kolthoff, J.P.; Göker, M.; Spröer, C.; Klenk, H.-P. When should a DDH experiment be mandatory in microbial taxonomy? Arch. Microbiol. 2013, 195, 413–418. [Google Scholar] [CrossRef] [PubMed]
- Richter, M.; Rossello-Mora, R. Shifting the genomic gold standard for the prokaryotic species definition. Proc. Natl. Acad. Sci. USA 2009, 106, 19126–19131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, M.; Oh, H.-S.; Park, S.-C.; Chun, J. Towards a taxonomic coherence between average nucleotide identity and 16S rRNA gene sequence similarity for species demarcation of prokaryotes. Int. J. Syst. Evol. Microbiol. 2014, 64, 346–351. [Google Scholar] [CrossRef] [PubMed]
- Awan, F.; Dong, Y.; Liu, J.; Wang, N.; Mushtaq, M.H.; Lu, C.; Liu, Y. Comparative genome analysis provides deep insights into Aeromonas hydrophila taxonomy and virulence-related factors. BMC Genom. 2018, 19, 712. [Google Scholar] [CrossRef] [Green Version]
- Talagrand-Reboul, E.; Roger, F.; Kimper, J.-L.; Colston, S.M.; Graf, J.; Latif-Eugenín, F.; Figueras, M.J.; Petit, F.; Marchandin, H.; Jumas-Bilak, E.; et al. Delineation of taxonomic species within complex of species: Aeromonas media and related species as a test case. Front. Microbiol. 2017, 8, 23. [Google Scholar] [CrossRef]
- Vincent, A.T.; Charette, S.J. Completion of genome of Aeromonas salmonicida subsp. salmonicida 01-B526 reveals how sequencing technologies can influence sequence quality and result interpretations. New Microbes New Infect. 2018, 25, 24–26. [Google Scholar] [CrossRef]
- Pessoa, R.B.G.; De Oliveira, W.F.; Marques, D.S.C.; Correia, M.T.D.S.; De Carvalho, E.V.M.M.; Coelho, L.C.B.B. The genus Aeromonas: A general approach. Microb. Pathog. 2019, 130, 81–94. [Google Scholar] [CrossRef]
- Monfort, P.; Baleux, B. Dynamics of Aeromonas hydrophila, Aeromonas sobria, and Aeromonas caviae in a sewage treatment pond. Appl. Environ. Microbiol. 1990, 56, 1999–2006. [Google Scholar] [CrossRef] [Green Version]
- Dumontet, S.; Krovacek, K.; Baloda, S.; Grottoli, R.; Pasquale, V.; Vanucci, S. Ecological relationship between Aeromonas and Vibrio spp. and planktonic copepods in the coastal marine environment in Southern Italy. Comp. Immunol. Microbiol. Infect. Dis. 1996, 19, 245–254. [Google Scholar] [CrossRef]
- Borrell, N.; Figueras, M.J.; Guarro, J. Phenotypic identification of Aeromonas genomospecies from clinical and environmental sources. Can. J. Microbiol. 1998, 44, 103–108. [Google Scholar] [CrossRef] [PubMed]
- Montes, M.; Perez, M.J.; Nieto, T.P. Numerical taxonomy of gram-negative, facultative anaerobic bacteria isolated from skin of turbot (Scophthalmus maximus) and surrounding water. Syst. Appl. Microbiol. 1999, 22, 604–618. [Google Scholar] [CrossRef]
- Egorov, A.I.; Best, J.M.B.; Frebis, C.P.; Karapondo, M.S. Occurrence of Aeromonas spp. in a random sample of drinking water distribution systems in the USA. J. Water Heal. 2011, 9, 785–798. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, M.; Resta, R.; Car, R. Intermolecular dynamical charge fluctuations in water: A signature of the H-bond network. Phys. Rev. Lett. 2005, 95, 187401. [Google Scholar] [CrossRef] [PubMed]
- Araujo, R.M.; Parés, R.; Lucena, F. The effect of terrestrial effluents on the incidence of Aeromonas spp. in coastal waters. J. Appl. Bacteriol. 1990, 69, 439–444. [Google Scholar] [CrossRef]
- Berg, K.A.; Lyra, C.; Sivonen, K.; Paulin, L.; Suomalainen, S.; Tuomi, P.; Rapala, J. High diversity of cultivable heterotrophic bacteria in association with cyanobacterial water blooms. ISME J. 2009, 3, 314–325. [Google Scholar] [CrossRef]
- Berg, K.A.; Lyra, C.; Niemi, R.M.; Heens, B.; Hoppu, K.; Erkomaa, K.; Sivonen, K.; Rapala, J. Virulence genes of Aeromonas isolates, bacterial endotoxins and cyanobacterial toxins from recreational water samples associated with human health symptoms. J. Water Heal. 2011, 9, 670–679. [Google Scholar] [CrossRef] [Green Version]
- McLellan, S.L.; Huse, S.M.; Mueller-Spitz, S.R.; Andreishcheva, E.N.; Sogin, M.L. Diversity and population structure of sewage-derived microorganisms in wastewater treatment plant influent. Environ. Microbiol. 2010, 12, 378–392. [Google Scholar] [CrossRef] [Green Version]
- Al-Jassim, N.; Ansari, M.I.; Harb, M.; Hong, P.-Y. Removal of bacterial contaminants and antibiotic resistance genes by conventional wastewater treatment processes in Saudi Arabia: Is the treated wastewater safe to reuse for agricultural irrigation? Water Res. 2015, 73, 277–290. [Google Scholar] [CrossRef] [Green Version]
- Varela, A.R.; Nunes, O.C.; Manaia, C.M. Quinolone resistant Aeromonas spp. as carriers and potential tracers of acquired antibiotic resistance in hospital and municipal wastewater. Sci. Total. Environ. 2016, 542, 665–671. [Google Scholar] [CrossRef] [PubMed]
- Holmes, P.; Nicolls, L.M.; Sartory, D.P. The ecology of mesophilic Aeromonas in the aquatic environment. In The Genus Aeromonas; John Wiley & Sons Ltd.: West Sussex, UK, 1996; pp. 127–150. [Google Scholar]
- Erhart, R.; Beimfohr, C.; Wagner, M.; Amann, R. Characterization of bacterial communities from activated sludge: Culture-dependent numerical identification versus in situ identification using group- and genus-specific rRNA-targeted oligonucleotide probes. Microb. Ecol. 1996, 32, 101–121. [Google Scholar]
- Latif-Eugenín, F.; Beaz-Hidalgo, R.; José, F.M. A culture independent method for the detection of Aeromonas sp. from water samples. Ital. J. Food Saf. 2016, 5, 5489. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Latif-Eugenín, F.; Beaz-Hidalgo, R.; Silvera-Simón, C.; Fernandez-Cassi, X.; Figueras, M.J. Chlorinated and ultraviolet radiation -treated reclaimed irrigation water is the source of Aeromonas found in vegetables used for human consumption. Environ. Res. 2017, 154, 190–195. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Cassi, X.; Silvera, C.; Cervero-Aragó, S.; Rusiñol, M.; Latif-Eugeni, F.; Bruguera-Casamada, C.; Civit, S.; Araujo, R.M.; Figueras, M.J.; Gironés, R.; et al. Evaluation of the microbiological quality of reclaimed water produced from a lagooning system. Environ. Sci. Pollut. Res. 2016, 23, 16816–16833. [Google Scholar] [CrossRef]
- Boussaid, A.; Baleux, B.; Hassani, L.; Lesne, J. Aeromonas species in stabilization ponds in the arid region of Marrakesh, Morocco, and relation to fecal-pollution and climatic factors. Microb. Ecol. 1991, 21, 11–20. [Google Scholar] [CrossRef]
- Hassani, L.; Imziln, B.; Boussaid, A.; Gauthier, M.J. Seasonal incidence of and antibiotic resistance among Aeromonas species isolated from domestic wastewater before and after treatment in stabilization ponds. Microb. Ecol. 1992, 23, 227–237. [Google Scholar] [CrossRef]
- Jjemba, P.K.; Weinrich, L.A.; Cheng, W.; Giraldo, E.; Lechevallier, M.W. Regrowth of potential opportunistic pathogens and algae in reclaimed-water distribution systems. Appl. Environ. Microbiol. 2010, 76, 4169–4178. [Google Scholar] [CrossRef] [Green Version]
- Pianetti, A.; Bruscolini, F.; Sabatini, L.; Colantoni, P. Microbial characteristics of marine sediments in bathing area along Pesaro-Gabicce coast (Italy): A preliminary study. J. Appl. Microbiol. 2004, 97, 682–689. [Google Scholar] [CrossRef]
- Khajanchi, B.K.; Fadl, A.A.; Borchardt, M.A.; Berg, R.L.; Horneman, A.J.; Stemper, M.E.; Joseph, S.W.; Moyer, N.P.; Sha, J.; Chopra, A.K. Distribution of virulence factors and molecular fingerprinting of Aeromonas species isolates from water and clinical samples: Suggestive evidence of water-to-human transmission. Appl. Environ. Microbiol. 2010, 76, 2313–2325. [Google Scholar] [CrossRef] [Green Version]
- Pablos, M.; Huys, G.; Cnockaert, M.; Rodríguez-Calleja, J.M.; Otero, A.; Santos, J.A.; García-López, M.-L. Identification and epidemiological relationships of Aeromonas isolates from patients with diarrhea, drinking water and foods. Int. J. Food Microbiol. 2011, 147, 203–210. [Google Scholar] [CrossRef] [PubMed]
- Van Der Kooij, D.; Martijn, B.; Schaap, P.G.; Hoogenboezem, W.; Veenendaal, H.R.; Van Der Wielen, P.W. Improved biostability assessment of drinking water with a suite of test methods at a water supply treating eutrophic lake water. Water Res. 2015, 87, 347–355. [Google Scholar] [CrossRef] [PubMed]
- Sisti, M.; Albano, A.; Brandi, G. Bactericidal effect of chlorine on motile Aeromonas spp. in drinking water supplies and influence of temperature on disinfection efficacy. Lett. Appl. Microbiol. 1998, 26, 347–351. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.-C.; Wang, J.-Y.; Selvam, A.; Kao, S.-C.; Yang, S.-S.; Shih, D.Y.-C. Multiplex PCR detection of enterotoxin genes in Aeromonas spp. from suspect food samples in northern Taiwan. J. Food Prot. 2008, 71, 2094–2099. [Google Scholar] [CrossRef]
- Nagar, V.; Shashidhar, R.; Bandekar, J.R. Prevalence, characterization, and antimicrobial resistance of Aeromonas strains from various retail food products in Mumbai, India. J. Food Sci. 2011, 76, M486–M492. [Google Scholar] [CrossRef]
- Nagar, V.; Shashidhar, R.; Bandekar, J.R. Characterization of Aeromonas strains isolated from Indian foods using rpoD gene sequencing and whole cell protein analysis. World J. Microbiol. Biotechnol. 2013, 29, 745–752. [Google Scholar] [CrossRef]
- Hoel, S.; Vadstein, O.; Jakobsen, A.N. Species distribution and prevalence of putative virulence factors in mesophilic Aeromonas spp. isolated from fresh retail sushi. Front. Microbiol. 2017, 8, 931. [Google Scholar] [CrossRef]
- Hoel, S.; Vadstein, O.; Jakobsen, A.N. The significance of mesophilic Aeromonas spp. in minimally processed ready-to-eat seafood. Microorganisms 2019, 7, 91. [Google Scholar] [CrossRef] [Green Version]
- Figueras, M.J.; Beaz-Hidalgo, R. Aeromonas: Introduction, detection by cultural and modern techniques. In Encyclopedia of Food Microbiology; Academic Press: London, UK, 2014; pp. 25–30. [Google Scholar]
- Hammad, A.M.; Moustafa, A.-E.H.; Mansour, M.M.; Fahmy, B.M.; Hamada, M.G.; Shimamoto, T.; Shimamoto, T. Molecular and phenotypic analysis of hemolytic Aeromonas strains isolated from food in Egypt revealed clinically important multidrug resistance and virulence profiles. J. Food Prot. 2018, 81, 1015–1021. [Google Scholar] [CrossRef]
- John, N.; Vidyalakshmi, V.; Hatha, A.M. Effect of pH and salinity on the production of extracellular virulence factors by Aeromonas from food sources. J. Food Sci. 2019, 84, 2250–2255. [Google Scholar] [CrossRef]
- Praveen, P.K.; Debnath, C.; Shekhar, S.; Dalai, N.; Ganguly, S. Incidence of Aeromonas spp. infection in fish and chicken meat and its related public health hazards: A review. Veter. World 2016, 9, 6–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, Y.; Xu, C.; Sun, Q.; Schwarz, S.; Ou, Y.; Yang, L.; Huang, Z.; Eichhorn, I.; Walsh, T.R.; Wang, Y.; et al. Prevalence and genetic analysis of mcr-3-positive Aeromonas species from humans, retail meat, and environmental water samples. Antimicrob. Agents Chemother. 2018, 62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castro-Escarpulli, G.; Figueras, M.J.; Aguilera-Arreola, G.; Soler, L.; Fernández-Rendón, E.; Aparicio, G.O.; Guarro, J.; Chacón, M.R. Characterisation of Aeromonas spp. isolated from frozen fish intended for human consumption in Mexico. Int. J. Food Microbiol. 2003, 84, 41–49. [Google Scholar] [CrossRef]
- Woodring, J.; Srijan, A.; Puripunyakom, P.; Oransathid, W.; Wongstitwilairoong, B.; Mason, C. Prevalence and antimicrobial susceptibilities of Vibrio, Salmonella, and Aeromonas isolates from various uncooked seafoods in Thailand. J. Food Prot. 2012, 75, 41–47. [Google Scholar] [CrossRef]
- Yano, Y.; Hamano, K.; Tsutsui, I.; Aue-Umneoy, D.; Ban, M.; Satomi, M. Occurrence, molecular characterization, and antimicrobial susceptibility of Aeromonas spp. in marine species of shrimps cultured at inland low salinity ponds. Food Microbiol. 2015, 47, 21–27. [Google Scholar] [CrossRef]
- Figueras, M.J.; Borrego, J.J. New perspectives in monitoring drinking water microbial quality. Int. J. Environ. Res. Public Heal. 2010, 7, 4179–4202. [Google Scholar] [CrossRef] [Green Version]
- Farmer, J.J.; Arduino, M.J.; Hickman-Brenner, F.W. The genera Aeromonas and Plesiomonas. In The Prokaryotes; Springer Science and Business Media: Berlin/Heidelberg, Germany, 2006; pp. 564–596. [Google Scholar]
- Rasmussen-Ivey, C.R.; Hossain, M.J.; Odom, S.E.; Terhune, J.S.; Hemstreet, W.G.; Shoemaker, C.A.; Zhang, D.; Xu, D.-H.; Griffin, M.J.; Liu, Y.-J.; et al. Classification of a hypervirulent Aeromonas hydrophila pathotype responsible for epidemic outbreaks in warm-water fishes. Front. Microbiol. 2016, 7, 334. [Google Scholar] [CrossRef] [Green Version]
- Nawaz, M.; Khan, S.A.; Khan, A.A.; Sung, K.; Tran, Q.; Kerdahi, K.; Steele, R. Detection and characterization of virulence genes and integrons in Aeromonas veronii isolated from catfish. Food Microbiol. 2010, 27, 327–331. [Google Scholar] [CrossRef]
- Li, Y.; Cai, S.H. Identification and pathogenicity of Aeromonas sobria on tail-rot disease in juvenile tilapia Oreochromis niloticus. Curr. Microbiol. 2011, 62, 623–627. [Google Scholar] [CrossRef]
- Chen, Y.F.; Liang, R.S.; Zhuo, X.L.; Wu, X.T.; Zou, J.X. Isolation and characterization of Aeromonas schubertii from diseased snakehead, Channa maculata (Lacepède). J. Fish Dis. 2012, 35, 421–430. [Google Scholar] [CrossRef]
- Yi, S.-W.; You, M.-J.; Cho, H.-S.; Lee, C.-S.; Kwon, J.-K.; Shin, G.-W. Molecular characterization of Aeromonas species isolated from farmed eels (Anguilla japonica). Veter. Microbiol. 2013, 164, 195–200. [Google Scholar] [CrossRef] [PubMed]
- Esteve, C.; Gutiérrez, M.C.; Ventosa, A. DNA Relatedness among Aeromonas allosaccharophila strains and DNA hybridization groups of the genus Aeromonas. Int. J. Syst. Bacteriol. 1995, 45, 390–391. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Esteve, C.; Alcaide, E.; Blasco, M.D. Aeromonas hydrophila subsp. dhakensis isolated from feces, water and fish in Mediterranean Spain. Microbes Environ. 2012, 27, 367–373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghenghesh, K.S.; Abeid, S.S.; Jaber, M.M.; Ben-Taher, S.A. Isolation and haemolytic activity of Aeromonas species from domestic dogs and cats. Comp. Immunol. Microbiol. Infect. Dis. 1999, 22, 175–179. [Google Scholar] [CrossRef]
- Forga-Martel, J.; Gonzalez-Valle, F.; Weinzierl, J. Infectious abortion associated with Aeromonas hydrophila in a mare. Equine Pract. 2000, 22, 22–24. [Google Scholar]
- Waldridge, B.M.; Stewart, A.J.; Taylor, D.C.; Saville, W.J. The incidence of Aeromonas species in the feces of nondiarrheic horses. J. Equine Veter. Sci. 2011, 31, 700–702. [Google Scholar] [CrossRef]
- Worthing, K.A.; Norris, J.M.; Briscoe, K.A. Severe acute cellulitis and sepsis caused by Aeromonas spp. in a dog on immunosuppressive therapy. J. Veter. Emerg. Crit. Care 2019, 29, 444–449. [Google Scholar] [CrossRef]
- Gugliandolo, C.; Irrera, G.; Lentini, V.; Maugeri, T. Pathogenic Vibrio, Aeromonas and Arcobacter spp. associated with copepods in the Straits of Messina (Italy). Mar. Pollut. Bull. 2008, 56, 600–606. [Google Scholar] [CrossRef]
- Pu, W.; Guo, G.; Yang, N.; Li, Q.; Yin, F.; Wang, P.; Zheng, J.; Zeng, J. Three species of Aeromonas (A. dhakensis, A. hydrophila and A. jandaei) isolated from freshwater crocodiles (Crocodylus siamensis) with pneumonia and septicemia. Lett. Appl. Microbiol. 2019, 68, 212–218. [Google Scholar] [CrossRef]
- King, G.E.; Werner, S.B.; Kizer, K.W. Epidemiology of Aeromonas infections in California. Clin. Infect. Dis. 1992, 15, 449–452. [Google Scholar] [CrossRef]
- Lamy, B.; Kodjo, A.; Laurent, F. Prospective nationwide study of Aeromonas infections in France. J. Clin. Microbiol. 2009, 47, 1234–1237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, C.-J.; Chen, P.-L.; Tang, H.-J.; Chen, H.-M.; Tseng, F.-C.; Shih, H.-I.; Hung, Y.-P.; Chung, C.-H.; Ko, W.-C. Incidence of Aeromonas bacteremia in southern Taiwan: Vibrio and Salmonella bacteremia as comparators. J. Microbiol. Immunol. Infect. 2014, 47, 145–148. [Google Scholar] [CrossRef] [Green Version]
- Mathur, P.; Batra, P.; Misra, M.C. Aeromonas spp.: An emerging nosocomial pathogen. J. Lab. Physicians 2016, 8, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Castro Escarpulli, G.; Pena del Barrio, D.; Castaneda, N.; Garcia Azcuaga, A.; Morier Dias, L.; Aguilera-Arreola, M.G.; Bravo Farias, L. Virulence factors of A. caviae strains isolated from acute diarrheic disease in Cuba. Rev. Latinoam. Microbiol. 2002, 44, 11–13. [Google Scholar] [PubMed]
- Tsai, Y.-H.; Shen, S.-H.; Yang, T.-Y.; Chen, P.-H.; Huang, K.-C.; Lee, M.S. Monomicrobial necrotizing fasciitis caused by Aeromonas hydrophila and Klebsiella pneumoniae. Med. Princ. Pract. 2015, 24, 416–423. [Google Scholar] [CrossRef] [PubMed]
- Hochedez, P.; Hope-Rapp, E.; Olive, C.; Nicolas, M.; Beaucaire, G.; Cabie, A. Bacteremia caused by Aeromonas species [corrected] complex in the Caribbean Islands of Martinique and Guadeloupe. Am. J. Trop. Med. Hyg. 2010, 83, 1123–1127. [Google Scholar] [CrossRef] [Green Version]
- Su, S.Y.; Chao, C.M.; Lai, C.C. Aeromonas bacteremia in southern Taiwan. Scand. J. Infect. Dis. 2013, 45, 728. [Google Scholar] [CrossRef]
- Igbinosa, I.H.; Igumbor, E.U.; Aghdasi, F.; Tom, M.; Okoh, A.I. Emerging Aeromonas species infections and their significance in public health. Sci. World J. 2012, 2012, 1–13. [Google Scholar]
- Aravena-Román, M.; Chang, B.J.; Riley, T.V.; Inglis, T.J. Phenotypic characteristics of human clinical and environmental Aeromonas in Western Australia. Pathology 2011, 43, 350–356. [Google Scholar] [CrossRef]
- Puthucheary, S.D.; Puah, S.M.; Chua, K.H. Molecular characterization of clinical isolates of Aeromonas species from Malaysia. PLoS ONE 2012, 7. [Google Scholar] [CrossRef] [Green Version]
- Huys, G.; Denys, R.; Albert, M.J.; Swings, J.; Kühn, I.; Kampfer, P. Aeromonas hydrophila subsp. dhakensis subsp. nov., isolated from children with diarrhoea in Bangladesh, and extended description of Aeromonas hydrophila subsp. hydrophila (Chester 1901) Stanier 1943 (approved lists 1980). Int. J. Syst. Evol. Microbiol. 2002, 52, 705–712. [Google Scholar] [PubMed] [Green Version]
- Hua, H.T.; Bollet, C.; Tercian, S.; Drancourt, M.; Raoult, D. Aeromonas popoffii urinary tract infection. J. Clin. Microbiol. 2004, 42, 5427–5428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al-Benwan, K.; Abbott, S.; Janda, J.M.; Huys, G.; Albert, M.J. Cystitis caused by Aeromonas caviae. J. Clin. Microbiol. 2007, 45, 2348–2350. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bossi-Kupfer, M.; Genini, A.; Peduzzi, R.; Demarta, A. Tracheobronchitis caused by Aeromonas veronii biovar sobria after near-drowning. J. Med. Microbiol. 2007, 56, 1563–1564. [Google Scholar] [CrossRef]
- Figueras, M.J.; Alperi, A.; Saavedra, M.J.; Ko, W.-C.; Gonzalo, N.; Navarro, M.; Martínez-Murcia, A.J. Clinical relevance of the recently described species Aeromonas aquariorum. J. Clin. Microbiol. 2009, 47, 3742–3746. [Google Scholar] [CrossRef] [Green Version]
- Tena, D.; Aspiroz, C.; Figueras, M.J.; González-Praetorius, A.; Aldea, M.J.; Alperi, A.; Bisquert, J. Surgical site infection due to Aeromonas species: Report of nine cases and literature review. Scand. J. Infect. Dis. 2009, 41, 164–170. [Google Scholar] [CrossRef]
- Alperi, A.; Figueras, M.J. Human isolates of Aeromonas possess Shiga toxin genes (stx1 and stx2) highly similar to the most virulent gene variants of Escherichia coli. Clin. Microbiol. Infect. 2010, 16, 1563–1567. [Google Scholar] [CrossRef] [Green Version]
- Sedláček, I.; Krejčí, E.; Andělová, A.; Sedláčková, M.; Porazilová, I.; Holochová, P. Aeromonas hydrophila subsp. Dhakensis—A causative agent of gastroenteritis imported into the Czech Republic. Ann. Agric. Environ. Med. 2012, 19, 409–413. [Google Scholar]
- Senderovich, Y.; Ken-Dror, S.; Vainblat, I.; Blau, D.; Izhaki, I.; Halpern, M. A molecular study on the prevalence and virulence potential of Aeromonas spp. recovered from patients suffering from diarrhea in Israel. PLoS ONE 2012, 7, e30070. [Google Scholar] [CrossRef]
- Chen, P.-L.; Tsai, P.-J.; Chen, C.-S.; Lu, Y.-C.; Chen, H.-M.; Lee, N.-Y.; Lee, C.-C.; Li, C.-W.; Li, M.-C.; Wu, C.-J.; et al. Aeromonas stool isolates from individuals with or without diarrhea in southern Taiwan: Predominance of Aeromonas veronii. J. Microbiol. Immunol. Infect. 2015, 48, 618–624. [Google Scholar] [CrossRef] [Green Version]
- Morinaga, Y.; Yanagihara, K.; Eugenin, F.L.L.; Beaz-Hidalgo, R.; Kohno, S.; Salvat, M.J.F. Identification error of Aeromonas aquariorum: A causative agent of septicemia. Diagn. Microbiol. Infect. Dis. 2013, 76, 106–109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shin, G.-W.; You, M.-J.; Cho, H.-S.; Yi, S.-W.; Lee, C.-S. Severe sepsis due to Aeromonas aquariorum in a patient with liver cirrhosis. Jpn. J. Infect. Dis. 2013, 66, 519–522. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perez-Valdespino, A.; Celestino-Mancera, M.; Villegas-Rodriguez, V.L.; Curiel-Quesada, E. Characterization of mercury-resistant clinical Aeromonas species. Braz. J. Microbiol. 2013, 44, 1279–1283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, H.; Zou, X.; Zhao, X.; Cai, L.; Chen, W.; Yang, L.; Liu, F.; Qin, D.; Zeng, G.; Bai, T.; et al. Fatal Aeromonas bacteraemia in West Africa. J. Infect. 2016, 72, 258–260. [Google Scholar] [CrossRef]
- Wu, C.-J.; Ko, W.-C.; Lee, N.-Y.; Su, S.-L.; Li, C.-W.; Li, M.-C.; Chen, Y.-W.; Su, Y.-C.; Shu, C.-Y.; Lin, Y.-T.; et al. Aeromonas isolates from fish and patients in Tainan City, Taiwan: Genotypic and phenotypic characteristics. Appl. Environ. Microbiol. 2019, 85. [Google Scholar] [CrossRef]
- Sinclair, H.A.; Heney, C.; Sidjabat, H.E.; George, N.M.; Bergh, H.; Anuj, S.N.; Nimmo, G.R.; Paterson, D.L. Genotypic and phenotypic identification of Aeromonas species and CphA-mediated carbapenem resistance in Queensland, Australia. Diagn. Microbiol. Infect. Dis. 2016, 85, 98–101. [Google Scholar] [CrossRef]
- Da Silva, L.C.A.; Leal-Balbino, T.C.; De Melo, B.S.T.; Mendes-Marques, C.L.; Rezende, A.M.; De Almeida, A.M.P.; Leal, N.C. Genetic diversity and virulence potential of clinical and environmental Aeromonas spp. isolates from a diarrhea outbreak. BMC Microbiol. 2017, 17, 179. [Google Scholar] [CrossRef]
- Van Zwetselaar, M.; Nyombi, B.; Sonda, T.; Kumburu, H.; Chamba, N.; Dekker, M.C.J.; Kilonzo, K.G.; Urasa, S.J.; Mmbaga, B.T. Aeromonas caviae mimicking Vibrio cholerae infectious enteropathy in a cholera-endemic region with possible public health consequences: Two case reports. J. Med. Case Rep. 2018, 12, 71. [Google Scholar] [CrossRef]
- Dallagassa, C.B.; Surek, M.; Vizzotto, B.S.; Prediger, K.C.; Moriel, B.; Wolf, S.; Weiss, V.; Cruz, L.M.; Assis, F.E.; Paludo, K.S.; et al. Characteristics of an Aeromonas trota strain isolated from cerebrospinal fluid. Microb. Pathog. 2018, 116, 109–112. [Google Scholar] [CrossRef]
- Khor, W.C.; Puah, S.M.; Koh, T.H.; Tan, J.A.M.A.; Puthucheary, S.D.; Chua, K.H. Comparison of clinical isolates of Aeromonas from Singapore and Malaysia with regard to molecular identification, virulence, and antimicrobial profiles. Microb. Drug Resist. 2018, 24, 469–478. [Google Scholar] [CrossRef]
- Zhou, Y.; Yu, L.; Nan, Z.; Zhang, P.; Kan, B.; Yan, D.; Su, J. Taxonomy, virulence genes and antimicrobial resistance of Aeromonas isolated from extra-intestinal and intestinal infections. BMC Infect. Dis. 2019, 19, 158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mosser, T.; Talagrand-Reboul, E.; Colston, S.M.; Graf, J.; Figueras, M.J.; Jumas-Bilak, E.; Lamy, B. Exposure to pairs of Aeromonas strains enhances virulence in the Caenorhabditis elegans infection model. Front. Microbiol. 2015, 6, 3785. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Joseph, S.W.; Carnahan, A.M.; Brayton, P.R.; Fanning, G.R.; Almazan, R.; Drabick, C.; Trudo, E.W.; Colwell, R.R. Aeromonas jandaei and Aeromonas veronii dual infection of a human wound following aquatic exposure. J. Clin. Microbiol. 1991, 29, 565–569. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shak, J.R.; Whitaker, J.A.; Ribner, B.S.; Burd, E.M. Aminoglycoside-resistant Aeromonas hydrophila as part of a polymicrobial infection following a traumatic fall into freshwater. J. Clin. Microbiol. 2011, 49, 1169–1170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grim, C.J.; Kozlova, E.V.; Ponnusamy, D.; Fitts, E.C.; Sha, J.; Kirtley, M.L.; Van Lier, C.J.; Tiner, B.L.; Erova, T.E.; Joseph, S.J.; et al. Functional genomic characterization of virulence factors from necrotizing fasciitis-causing strains of Aeromonas hydrophila. Appl. Environ. Microbiol. 2014, 80, 4162–4183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ponnusamy, D.; Kozlova, E.V.; Sha, J.; Erova, T.E.; Azar, S.R.; Fitts, E.C.; Kirtley, M.L.; Tiner, B.L.; Andersson, J.A.; Grim, C.J.; et al. Cross-talk among flesh-eating Aeromonas hydrophila strains in mixed infection leading to necrotizing fasciitis. Proc. Natl. Acad. Sci. USA 2016, 113, 722–727. [Google Scholar] [CrossRef] [Green Version]
- Fernández-Bravo, A.; Kilgore, P.B.; Andersson, J.A.; Blears, E.; Figueras, M.J.; Hasan, N.A.; Colwell, R.R.; Sha, J.; Chopra, A.K. T6SS and ExoA of flesh-eating Aeromonas hydrophila in peritonitis and necrotizing fasciitis during mono- and polymicrobial infections. Proc. Natl. Acad. Sci. USA 2019, 116, 24084–24092. [Google Scholar] [CrossRef]
- Von Graevenitz, A. The role of Aeromonas in diarrhea: A Review. Infection 2007, 35, 59–64. [Google Scholar] [CrossRef]
- Ottaviani, D.; Leoni, F.; Rocchegiani, E.; Santarelli, S.; Masini, L.; D’Annibale, M.L.; Pianetti, A.; Carraturo, A. A severe case of Aeromonas veronii biovar sobria travellers’ diarrhoea characterized by Vibrio parahaemolyticus co-isolation. J. Med. Microbiol. 2013, 62, 161–164. [Google Scholar] [CrossRef] [Green Version]
- Qamar, F.N.; Nisar, M.I.; Quadri, F.; Shakoor, S.; Sow, S.O.; Nasrin, D.; Blackwelder, W.C.; Wu, Y.; Farag, T.; Panchalingham, S.; et al. Aeromonas-associated diarrhea in children under 5 Years: The GEMS experience. Am. J. Trop. Med. Hyg. 2016, 95, 774–780. [Google Scholar] [CrossRef] [Green Version]
- Teunis, P.; Figueras, M.J. Reassessment of the enteropathogenicity of mesophilic Aeromonas species. Front. Microbiol. 2016, 7, 643. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Figueras, M.J.; Horneman, A.J.; Martinez-Murcia, A.; Guarro, J. Controversial data on the association of Aeromonas with diarrhoea in a recent Hong Kong study. J. Med. Microbiol. 2007, 56, 996–998. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mansour, A.M.; Abd Elkhalek, R.; Shaheen, H.I.; El Mohammady, H.; Refaey, S.; Hassan, K.; Riddle, M.; Sanders, J.W.; Sebeny, P.J.; Young, S.Y.; et al. Burden of Aeromonas hydrophila-associated diarrhea among children younger than 2 years in rural Egyptian community. J. Infect. Dev. Ctries. 2012, 6, 842–846. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schuetz, A.N. Emerging agents of gastroenteritis: Aeromonas, Plesiomonas, and the diarrheagenic pathotypes of Escherichia coli. Semin. Diagn. Pathol. 2019, 36, 187–192. [Google Scholar] [CrossRef]
- Figueras, M.J. Clinical relevance of Aeromonas sM503. Rev. Med. Microbiol. 2005, 16, 145–153. [Google Scholar] [CrossRef]
- Bravo, L.; Fernandez, A.; Nunez, F.A.; Rivero, L.A.; Ramirez, M.; Aguila, A.; Ledo, Y.; Cruz, Y.; Hernandez, J. Aeromonas spp asociated to acute diarrheic disease in Cuba: Case-control study. Rev. Chil. Infectol. 2012, 29, 44–48. [Google Scholar] [CrossRef]
- Chopra, A.K.; Houston, C.W. Enterotoxins in Aeromonas-associated gastroenteritis. Microbes Infect. 1999, 1, 1129–1137. [Google Scholar] [CrossRef]
- Vila, J.; Ruiz, J.; Gallardo, F.; Vargas, M.; Soler, L.; Figueras, M.J.; Gascón, J. Aeromonas spp. and traveler’s diarrhea: Clinical features and antimicrobial resistance. Emerg. Infect. Dis. 2003, 9, 552–555. [Google Scholar] [CrossRef]
- Granum, P.E.; O’Sullivan, K.; Tomas, J.M.; Ormen, O. Possible virulence factors of Aeromonas spp. from food and water. FEMS Immunol. Med. Microbiol. 1998, 21, 131–137. [Google Scholar] [CrossRef] [Green Version]
- Krovacek, K.; Dumontet, S.; Eriksson, E.; Baloda, S.B. Isolation, and Virulence Profiles, of Aeromonas hydrophila implicated in an outbreak of food poisoning in Sweden. Microbiol. Immunol. 1995, 39, 655–661. [Google Scholar] [CrossRef]
- Zhang, Q.; Shi, G.-Q.; Tang, G.-P.; Zou, Z.-T.; Yao, G.-H.; Zeng, G. A foodborne outbreak of Aeromonas hydrophila in a college, Xingyi City, Guizhou, China, 2012. West. Pac. Surveill. Response J. 2012, 3, 39–43. [Google Scholar] [CrossRef] [PubMed]
- Morgan, D.R.; Johnson, P.C.; Dupont, H.L.; Satterwhite, T.K.; Wood, L.V. Lack of correlation between known virulence properties of Aeromonas hydrophila and enteropathogenicity for humans. Infect. Immun. 1985, 50, 62–65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Altwegg, M.; Lucchini, G.M.; Lüthy-Hottenstein, J.; Rohrbach, M. Aeromonas-associated gastroenteritis after consumption of contaminated shrimp. Eur. J. Clin. Microbiol. Infect. Dis. 1991, 10, 44–45. [Google Scholar] [CrossRef] [PubMed]
- Janda, J.M.; Abbott, S.L. Evolving concepts regarding the genus Aeromonas: An expanding panorama of species, disease presentations, and unanswered questions. Clin. Infect. Dis. 1998, 27, 332–344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chao, C.-M.; Lai, C.-C.; Gau, S.-J.; Hsueh, P.-R. Skin and soft tissue infection caused by Aeromonas species in cancer patients. J. Microbiol. Immunol. Infect. 2013, 46, 144–146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hiransuthikul, N.; Tantisiriwat, W.; Lertutsahakul, K.; Vibhagool, A.; Boonma, P. Skin and soft-tissue infections among tsunami survivors in Southern Thailand. Clin. Infect. Dis. 2005, 41, 93–96. [Google Scholar] [CrossRef] [Green Version]
- Presley, S.M.; Rainwater, T.R.; Austin, G.P.; Platt, S.G.; Zak, J.C.; Cobb, G.P.; Marsland, E.J.; Tian, K.; Zhang, B.; Anderson, T.A.; et al. Assessment of pathogens and toxicants in New Orleans, LA following Hurricane Katrina. Environ. Sci. Technol. 2006, 40, 468–474. [Google Scholar] [CrossRef]
- Etemadi, J.; Ardalan, M.R.; Motavali, R.; Tubbs, R.S.; Shoja, M.M. Thrombotic microangiopathy as a complication of medicinal leech therapy. South. Med. J. 2008, 101, 845–847. [Google Scholar] [CrossRef]
- Sartor, C.; Bornet, C.; Guinard, D.; Fournier, P.-E. Transmission of Aeromonas hydrophila by leeches. Lancet 2013, 381, 1686. [Google Scholar] [CrossRef]
- Spadaro, S.; Berselli, A.; Marangoni, E.; Romanello, A.; Colamussi, M.V.; Ragazzi, R.; Zardi, S.; Volta, C.A. Aeromonas sobria necrotizing fasciitis and sepsis in an immunocompromised patient: A case report and review of the literature. J. Med. Case Rep. 2014, 8, 315. [Google Scholar] [CrossRef] [Green Version]
- Furusu, A.; Yoshizuka, N.; Abe, K.; Sasaki, O.; Miyazaki, K.; Miyazaki, M.; Hirakata, Y.; Ozono, Y.; Harada, T.; Kohno, S. Aeromonas hydrophila necrotizing fasciitis and gas gangrene in a diabetic patient on haemodialysis. Nephrol. Dial. Transplant. 1997, 12, 1730–1734. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Joseph, S.W.; Daily, O.P.; Hunt, W.S.; Seidler, R.J.; Allen, D.A.; Colwell, R.R. Aeromonas primary wound infection of a diver in polluted waters. J. Clin. Microbiol. 1979, 10, 46–49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sever, R.; Lee Goldstein, A.; Steinberg, E.; Soffer, D. Trauma with a touch of fresh water: Necrotizing fasciitis caused by Aeromonas hydrophila after a motorcycle accident. Am. Surg. 2013, 79, E326–E328. [Google Scholar] [PubMed]
- Pampin, F.; Bou, G.; Galeiras, R.; Freire, D.; Bouza, M.T.; Zuniga Mdel, C. Aeromonas and meningitis: An unusual presentation. Neurocirugia 2012, 23, 200–202. [Google Scholar] [CrossRef]
- Tang, H.-J.; Lai, C.-C.; Lin, H.-L.; Chao, C.-M. Clinical manifestations of bacteremia caused by Aeromonas species in southern Taiwan. PLoS ONE 2014, 9, e91642. [Google Scholar] [CrossRef]
- Wu, C.-J.; Chen, P.-L.; Hsueh, P.-R.; Chang, M.-C.; Tsai, P.-J.; Shih, H.-I.; Wang, H.-C.; Chou, P.-H.; Ko, W.-C. Clinical implications of species identification in monomicrobial Aeromonas bacteremia. PLoS ONE 2015, 10, e0117821. [Google Scholar] [CrossRef]
- Ohta, T. A Case of severe sepsis presenting marked decrease of neutrophils and interesting findings on dynamic CT. Am. J. Case Rep. 2015, 16, 322–327. [Google Scholar] [CrossRef]
- Cordingley, F.T.; Rajanayagam, A. Aeromonas hydrophila bacteraemia in haematological patients. Med. J. Aust. 1981, 1, 364–365. [Google Scholar] [CrossRef]
- Janda, J.M.; Guthertz, L.S.; Kokka, R.P.; Shimada, T. Aeromonas species in septicemia: Laboratory characteristics and clinical observations. Clin. Infect. Dis. 1994, 19, 77–83. [Google Scholar] [CrossRef]
- Shizuma, T.; Tanaka, C.; Mori, H.; Fukuyama, N. Investigation of bacteremia due to Aeromonas species and comparison with that due to enterobacteria in patients with liver cirrhosis. Gastroenterol. Res. Pract. 2011, 2011, 1–4. [Google Scholar] [CrossRef] [Green Version]
- Kao, H.-T.; Huang, Y.-C.; Lin, T.-Y. Fatal bacteremic pneumonia caused by Aeromonas hydrophila in a previously healthy child. J. Microbiol. Immunol. Infect. 2003, 36, 209–211. [Google Scholar] [PubMed]
- Nagata, K.; Takeshima, Y.; Tomii, K.; Imai, Y. Fulminant fatal bacteremic pneumonia due to Aeromonas hydrophila in a non-immunocompromised woman. Intern. Med. 2011, 50, 63–65. [Google Scholar] [CrossRef] [Green Version]
- Tena, D.; González-Praetorius, A.; Gimeno, C.; Pérez-Pomata, M.T.; Bisquert, J. Extraintestinal infection due to Aeromonas spp.: Review of 38 cases. Enferm. Infecc. Microbiol. Clín. 2007, 25, 235–241. [Google Scholar] [CrossRef] [PubMed]
- Mandal, J.; Dhodapkar, R.; Acharya, N.S.; Sastry, A.; Parija, S.C. Urinary tract infection due to Aeromonas spp., a lesser known causative bacterium. J. Infect. Dev. Ctries. 2010, 4, 679–681. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garrido, E.; Sanromán, A.L.; Rodríguez-Gandía, M.A.; Rivero, M.; Royuela, A.; Moreira, V.; Milicua, J.M. Optimized protocol for diagnosis of acute ileitis. Clin. Gastroenterol. Hepatol. 2009, 7, 1183–1188. [Google Scholar] [CrossRef]
- Lowry, R.; Balboa, S.; Parker, J.L.; Shaw, J.G. Aeromonas flagella and colonisation mechanisms. Adv. Microb. Physiol. 2014, 65, 203–256. [Google Scholar]
- Nazarenko, E.L.; Crawford, R.J.; Ivanova, E.P. The structural diversity of carbohydrate antigens of selected gram-negative marine bacteria. Mar. Drugs 2011, 9, 1914–1954. [Google Scholar] [CrossRef] [Green Version]
- Heiss, C.; Wang, Z.; Thurlow, C.M.; Hossain, M.J.; Sun, D.; Liles, M.R.; Saper, M.A.; Azadi, P. Structure of the capsule and lipopolysaccharide O-antigen from the channel catfish pathogen, Aeromonas hydrophila. Carbohydr. Res. 2019, 486, 107858. [Google Scholar] [CrossRef]
- Merino, S.; Tomás, J. Bacterial capsules and evasion of immune responses. eLS 2010. [Google Scholar] [CrossRef]
- Galindo, C.L.; Gutierrez, C., Jr.; Chopra, A.K. Potential involvement of galectin-3 and SNAP23 in Aeromonas hydrophila cytotoxic enterotoxin-induced host cell apoptosis. Microb. Pathog. 2006, 40, 56–68. [Google Scholar] [CrossRef]
- Merino, S.; Rubires, X.; Knøchel, S.; Tomás, J.M. Emerging pathogens: Aeromonas spp. Int. J. Food Microbiol. 1995, 28, 157–168. [Google Scholar] [CrossRef]
- Janda, J.M. Aeromonas and Plesiomonas. In Molecular Medical Microbiology; Academic Press: Cambridge, MA, USA, 2001; pp. 1237–1270. [Google Scholar]
- Palma-Martínez, I.; Guerrero-Mandujano, A.; Ruiz-Ruiz, M.J.; Hernández-Cortez, C.; Molina-López, J.; Bocanegra-García, V.; Castro-Escarpulli, G. Active Shiga-like toxin produced by some Aeromonas spp., isolated in Mexico City. Front. Microbiol. 2016, 7, 4496. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chopra, A.K.; Xu, X.-J.; Ribardo, D.; Gonzalez, M.; Kuhl, K.; Peterson, J.W.; Houston, C.W. The cytotoxic enterotoxin of Aeromonas hydrophila induces proinflammatory cytokine production and activates arachidonic acid metabolism in macrophages. Infect. Immun. 2000, 68, 2808–2818. [Google Scholar] [CrossRef] [Green Version]
- Singh, D.V.; Sanyal, S.C. Haemolysin and enterotoxin production by Aeromonas caviae isolated from diarrhoeal patients, fish and environment. J. Diarrhoeal Dis. Res. 1992, 10, 16–20. [Google Scholar] [PubMed]
- Bernheimer, A.W.; Avigad, L.S. Partial characterization of aerolysin, a lytic exotoxin from Aeromonas hydrophila. Infect. Immun. 1974, 9, 1016–1021. [Google Scholar] [CrossRef] [Green Version]
- Sha, J.; Erova, T.E.; Alyea, R.A.; Wang, S.; Olano, J.P.; Pancholi, V.; Chopra, A.K. Surface-expressed enolase contributes to the pathogenesis of clinical isolate SSU of Aeromonas hydrophila. J. Bacteriol. 2009, 191, 3095–3107. [Google Scholar] [CrossRef] [Green Version]
- Huang, L.; Qin, Y.; Yan, Q.; Lin, G.; Huang, L.; Huang, B.; Huang, W. MinD plays an important role in Aeromonas hydrophila adherence to Anguilla japonica mucus. Gene 2015, 565, 275–281. [Google Scholar] [CrossRef]
- Herold, S.; Karch, H.; Schmidt, H. Shiga toxin-encoding bacteriophages-genomes in motion. Int. J. Med. Microbiol. 2004, 294, 115–121. [Google Scholar] [CrossRef]
- Tseng, T.-T.; Tyler, B.M.; Setubal, J.C. Protein secretion systems in bacterial-host associations, and their description in the Gene Ontology. BMC Microbiol. 2009, 9, S2. [Google Scholar] [CrossRef] [Green Version]
- Cianciotto, N.P. Type II secretion: A protein secretion system for all seasons. Trends Microbiol. 2005, 13, 581–588. [Google Scholar] [CrossRef]
- Burr, S.E.; Stuber, K.; Wahli, T.; Frey, J. Evidence for a type III secretion system in Aeromonas salmonicida subsp. salmonicida. J. Bacteriol. 2002, 184, 5966–5970. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chacon, M.R.; Soler, L.; Groisman, E.A.; Guarro, J.; Figueras, M.J. Type III secretion system genes in clinical Aeromonas isolates. J. Clin. Microbiol. 2004, 42, 1285–1287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sha, J.; Wang, S.; Suarez, G.; Sierra, J.; Fadl, A.; Erova, T.; Foltz, S.; Khajanchi, B.; Silver, A.; Graf, J.; et al. Further characterization of a type III secretion system (T3SS) and of a new effector protein from a clinical isolate of Aeromonas hydrophila—Part I. Microb. Pathog. 2007, 43, 127–146. [Google Scholar] [CrossRef] [PubMed]
- Tan, Y.W.; Yu, H.B.; Leung, K.Y.; Sivaraman, J.; Mok, Y.-K. Structure of AscE and induced burial regions in AscE and AscG upon formation of the chaperone needle-subunit complex of type III secretion system in Aeromonas hydrophila. Protein Sci. 2008, 17, 1748–1760. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vilches, S.; Jimenez, N.; Merino, S.; Tomás, J.M. The Aeromonas dsbA mutation decreased their virulence by triggering type III secretion system but not flagella production. Microb. Pathog. 2012, 52, 130–139. [Google Scholar] [CrossRef] [PubMed]
- Vilches, S.; Urgell, C.; Merino, S.; Chacón, M.R.; Soler, L.; Castro-Escarpulli, G.; Figueras, M.J.; Tomás, J.M. Complete type III secretion system of a mesophilic Aeromonas hydrophila strain. Appl. Environ. Microbiol. 2004, 70, 6914–6919. [Google Scholar] [CrossRef] [Green Version]
- Vilches, S.; Wilhelms, M.; Yu, H.B.; Leung, K.Y.; Tomás, J.M.; Merino, S. Aeromonas hydrophila AH-3 AexT is an ADP-ribosylating toxin secreted through the type III secretion system. Microb. Pathog. 2008, 44, 1–12. [Google Scholar] [CrossRef]
- Yu, H.B.; Rao, P.S.S.; Lee, H.C.; Vilches, S.; Merino, S.; Tomas, J.M.; Leung, K.Y. A Type III secretion system is required for Aeromonas hydrophila AH-1 pathogenesis. Infect. Immun. 2004, 72, 1248–1256. [Google Scholar] [CrossRef] [Green Version]
- Burr, S.E.; Frey, J. Analysis of type III effector genes in typical and atypical Aeromonas salmonicida. J. Fish Dis. 2007, 30, 711–714. [Google Scholar] [CrossRef]
- Rangel, L.T.; Marden, J.; Colston, S.; Setubal, J.C.; Graf, J.; Gogarten, J.P. Identification and characterization of putative Aeromonas spp. T3SS effectors. PLoS ONE 2019, 14, e0214035. [Google Scholar] [CrossRef] [Green Version]
- Bingle, L.E.; Bailey, C.M.; Pallen, M.J. Type VI secretion: A beginner’s guide. Curr. Opin. Microbiol. 2008, 11, 3–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suarez, G.; Sierra, J.C.; Sha, J.; Wang, S.; Erova, T.E.; Fadl, A.A.; Foltz, S.M.; Horneman, A.J.; Chopra, A.K. Molecular characterization of a functional type VI secretion system from a clinical isolate of Aeromonas hydrophila. Microb. Pathog. 2008, 44, 344–361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- English, G.; Trunk, K.; Rao, V.A.; Srikannathasan, V.; Hunter, W.N.; Coulthurst, S.J. New secreted toxins and immunity proteins encoded within the type VI secretion system gene cluster of Serratia marcescens. Mol. Microbiol. 2012, 86, 921–936. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carruthers, M.D.; Nicholson, P.A.; Tracy, E.N.; Munson, R.S. Acinetobacter baumannii utilizes a type VI secretion system for bacterial competition. PLoS ONE 2013, 8, e59388. [Google Scholar] [CrossRef]
- Anderson, M.C.; Vonaesch, P.; Saffarian, A.; Marteyn, B.S.; Sansonetti, P.J. Shigella sonnei encodes a functional T6SS used for interbacterial competition and niche occupancy. Cell Host Microbe 2017, 21, 769–776. [Google Scholar] [CrossRef] [Green Version]
- Miller, M.B.; Bassler, B.L. Quorum sensing in bacteria. Annu. Rev. Microbiol. 2001, 55, 165–199. [Google Scholar] [CrossRef] [Green Version]
- Khajanchi, B.K.; Kirtley, M.L.; Brackman, S.M.; Chopra, A.K. Immunomodulatory and protective roles of quorum-sensing signaling molecules N-Acyl homoserine lactones during infection of mice with Aeromonas hydrophila. Infect. Immun. 2011, 79, 2646–2657. [Google Scholar] [CrossRef] [Green Version]
- Chan, X.-Y.; How, K.-Y.; Yin, W.-F.; Chan, K.-G. N-Acyl homoserine lactone-mediated quorum sensing in Aeromonas veronii biovar sobria strain 159: Identification of LuxRI homologs. Front. Microbiol. 2016, 6, 5772. [Google Scholar] [CrossRef] [Green Version]
- Blöcher, R.; Ramírez, A.R.; Castro-Escarpulli, G.; Curiel-Quesada, E.; Reyes-Arellano, A. design, synthesis, and evaluation of Alkyl-Quinoxalin-2(1H)-one derivatives as anti-quorum sensing molecules, inhibiting biofilm formation in Aeromonas caviae Sch3. Molecules 2018, 23, 3075. [Google Scholar] [CrossRef] [Green Version]
- Ding, T.; Li, T.; Li, J. Impact of curcumin liposomes with anti-quorum sensing properties against foodborne pathogens Aeromonas hydrophila and Serratia grimesii. Microb. Pathog. 2018, 122, 137–143. [Google Scholar] [CrossRef]
- Ding, T.; Li, T.; Wang, Z.; Li, J. Curcumin liposomes interfere with quorum sensing system of Aeromonas sobria and in silico analysis. Sci. Rep. 2017, 7, 8612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, L.; Yan, Y.; Feng, L.; Zhu, J. Quorum sensing asaI mutants affect spoilage phenotypes, motility, and biofilm formation in a marine fish isolate of Aeromonas salmonicida. Food Microbiol. 2018, 76, 40–51. [Google Scholar] [CrossRef] [PubMed]
- Patel, B.; Kumari, S.; Banerjee, R.; Samanta, M.; Das, S. Disruption of the quorum sensing regulated pathogenic traits of the biofilm-forming fish pathogen Aeromonas hydrophila by tannic acid, a potent quorum quencher. Biofouling 2017, 33, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Talagrand-Reboul, E.; Jumas-Bilak, E.; Lamy, B. The social life of Aeromonas through biofilm and quorum sensing systems. Front. Microbiol. 2017, 8, 449. [Google Scholar] [CrossRef] [PubMed]
- Waldron, K.J.; Rutherford, J.C.; Ford, D.; Robinson, N.J. Metalloproteins and metal sensing. Nature 2009, 460, 823–830. [Google Scholar] [CrossRef] [PubMed]
- Capdevila, D.A.; Wang, J.; Giedroc, D.P. Bacterial strategies to maintain zinc metallostasis at the host-pathogen interface. J. Boil. Chem. 2016, 291, 20858–20868. [Google Scholar] [CrossRef] [Green Version]
- O’Halloran, T.V.; Culotta, V.C. Metallochaperones, an intracellular shuttle service for metal ions. J. Boil. Chem. 2000, 275, 25057–25060. [Google Scholar] [CrossRef] [Green Version]
- Palmer, L.D.; Skaar, E.P. Transition metals and virulence in bacteria. Annu. Rev. Genet. 2016, 50, 67–91. [Google Scholar] [CrossRef] [Green Version]
- Fones, H.; Preston, G.M. The impact of transition metals on bacterial plant disease. FEMS Microbiol. Rev. 2013, 37, 495–519. [Google Scholar] [CrossRef] [Green Version]
- Francki, K.T.; Chang, B.J.; Mee, B.J.; Collignon, P.J.; Susai, V.; Keese, P.K. Identification of genes associated with copper tolerance in an adhesion-defective mutant of Aeromonas veronii biovar sobria. FEMS Immunol. Med. Microbiol. 2000, 29, 115–121. [Google Scholar] [CrossRef]
- Fernandez-Bravo, A.; Lopez-Fernandez, L.; Figueras, M.J. The metallochaperone encoding gene hypA is widely distributed among pathogenic Aeromonas spp. and its expression is increased under acidic pH and within macrophages. Microorganisms 2019, 7, 415. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Janeway, C.A., Jr. How the immune system protects the host from infection. Microbes Infect. 2001, 3, 1167–1171. [Google Scholar] [CrossRef]
- Bergsbaken, T.; Fink, S.L.; Cookson, B.T. Pyroptosis: Host cell death and inflammation. Nat. Rev. Genet. 2009, 7, 99–109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, H.; Kawai, T.; Akira, S. Pathogen recognition by the innate immune system. Int. Rev. Immunol. 2011, 30, 16–34. [Google Scholar] [CrossRef] [PubMed]
- Mahla, R.S.; Reddy, M.C.; Prasad, D.V.R.; Kumar, H. Sweeten PAMPs: Role of sugar complexed PAMPs in innate immunity and vaccine biology. Front. Immunol. 2013, 4, 248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kawai, T.; Akira, S. TLR signaling. Semin. Immunol. 2007, 19, 24–32. [Google Scholar] [CrossRef]
- Shrestha, N.; Bahnan, W.; Wiley, D.J.; Barber, G.; Fields, K.A.; Schesser, K. Eukaryotic initiation factor 2 (eIF2) signaling regulates proinflammatory cytokine expression and bacterial invasion. J. Boil. Chem. 2012, 287, 28738–28744. [Google Scholar] [CrossRef] [Green Version]
- Lamkanfi, M.; Dixit, V.M. Inflammasomes: Guardians of cytosolic sanctity. Immunol. Rev. 2009, 227, 95–105. [Google Scholar] [CrossRef]
- Saxena, M.; Yeretssian, G. NOD-Like receptors: Master regulators of inflammation and cancer. Front. Immunol. 2014, 5, 327. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.-T.; Zhang, G.-R.; Shi, Z.-C.; Yuan, Y.-J.; Zheng, H.; Lin, L.; Wei, K.-J.; Ji, W. Expression analysis of nine Toll-like receptors in yellow catfish (Pelteobagrus fulvidraco) responding to Aeromonas hydrophila challenge. Fish Shellfish. Immunol. 2017, 63, 384–393. [Google Scholar] [CrossRef]
- Gong, Y.; Feng, S.; Li, S.; Zhang, Y.; Zhao, Z.; Hu, M.; Xu, P.; Jiang, Y. Genome-wide characterization of Toll-like receptor gene family in common carp (Cyprinus carpio) and their involvement in host immune response to Aeromonas hydrophila infection. Comp. Biochem. Physiol. Part D Genom. Proteom. 2017, 24, 89–98. [Google Scholar] [CrossRef] [PubMed]
- Takeuchi, O.; Kawai, T.; Sanjo, H.; Copeland, N.; Gilbert, D.; Jenkins, N.; Takeda, K.; Akira, S. TLR6: A novel member of an expanding toll-like receptor family. Gene 1999, 231, 59–65. [Google Scholar] [CrossRef]
- Takeda, K. Toll-like receptor. Jpn. J. Clin. Immunol. 2005, 28, 309–317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lai, R.F.; Jakovlic, I.; Liu, H.; Wei, J.; Zhan, F.B.; Yang, P.H.; Wang, W.M. Characterization and expression of Megalobrama amblycephala toll-like receptor 22 involved in the response to Aeromonas hydrophila. J. Fish Biol. 2017, 90, 803–818. [Google Scholar] [CrossRef]
- Basu, M.; Swain, B.; Sahoo, B.R.; Maiti, N.K.; Samanta, M. Induction of toll-like receptor (TLR) 2, and MyD88-dependent TLR- signaling in response to ligand stimulation and bacterial infections in the Indian major carp, mrigal (Cirrhinus mrigala). Mol. Biol. Rep. 2012, 39, 6015–6028. [Google Scholar] [CrossRef]
- Basu, M.; Swain, B.; Maiti, N.; Routray, P.; Samanta, M. Inductive expression of toll-like receptor 5 (TLR5) and associated downstream signaling molecules following ligand exposure and bacterial infection in the Indian major carp, mrigal (Cirrhinus mrigala). Fish Shellfish. Immunol. 2012, 32, 121–131. [Google Scholar] [CrossRef]
- Merino, S.; Wilhelms, M.; Tomás, J.M. Role of Aeromonas hydrophila flagella glycosylation in adhesion to Hep-2 cells, biofilm formation and immune stimulation. Int. J. Mol. Sci. 2014, 15, 21935–21946. [Google Scholar] [CrossRef] [Green Version]
- Brietzke, A.; Korytář, T.; Jaros, J.; Köllner, B.; Goldammer, T.; Seyfert, H.-M.; Rebl, A. Aeromonas salmonicida infection only moderately regulates expression of factors contributing to toll-like receptor signaling but massively activates the cellular and humoral branches of innate immunity in rainbow trout (Oncorhynchus mykiss). J. Immunol. Res. 2015, 2015, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Reyes-Becerril, M.; Ascencio-Valle, F.; Hirono, I.; Kondo, H.; Jirapongpairoj, W.; Esteban, M.A.; Alamillo, E.; Angulo, C.; Valadez, C.E.A. TLR21’s agonists in combination with Aeromonas antigens synergistically up-regulate functional TLR21 and cytokine gene expression in yellowtail leucocytes. Dev. Comp. Immunol. 2016, 61, 107–115. [Google Scholar] [CrossRef]
- Zhang, J.; Li, L.; Kong, X.; Wu, F.; Zhou, C.; Nie, G.; Li, X. Expression patterns of Toll-like receptors in natural triploid Carassius auratus after infection with Aeromonas hydrophila. Veter. Immunol. Immunopathol. 2015, 168, 77–82. [Google Scholar] [CrossRef]
- Chandrarathna, H.; Nikapitiya, C.; Dananjaya, S.; Wijerathne, C.; Wimalasena, S.; Kwun, H.J.; Heo, G.-J.; Lee, J.; De Zoysa, M. Outcome of co-infection with opportunistic and multidrug resistant Aeromonas hydrophila and A. veronii in zebrafish: Identification, characterization, pathogenicity and immune responses. Fish Shellfish. Immunol. 2018, 80, 573–581. [Google Scholar] [CrossRef] [PubMed]
- Kong, W.-G.; Chen, X.-X.; Huang, Y.-Q.; Wu, Z.-X.; Li, S.-S.; Tang, Y. A study of the damage of the intestinal mucosa barrier structure and function of Ctenopharyngodon idella with Aeromonas hydrophila. Fish Physiol. Biochem. 2017, 43, 1223–1235. [Google Scholar] [CrossRef] [PubMed]
- Sierra, J.C.; Suárez, G.; Sha, J.; Baze, W.B.; Foltz, S.M.; Chopra, A.K. Unraveling the mechanism of action of a new type III secretion system effector AexU from Aeromonas hydrophila. Microb. Pathog. 2010, 49, 122–134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, Q.Q.; Xu, P.; Zhou, J.W.; Pan, T.S.; Tuo, R.; Ai, K.; Yang, D.Q. Cloning and expression analysis of two pro-inflammatory cytokines, IL-1beta and its receptor, IL-1R2, in the Asian swamp eel Monopterus albus. Mol. Biol. 2016, 50, 760–774. [Google Scholar] [CrossRef]
- Arockiaraj, J.; Bhatt, P.; Kumaresan, V.; Dhayanithi, N.B.; Arshad, A.; Harikrishnan, R.; Arasu, M.V.; Al-Dhabi, N.A. Fish chemokines 14, 20 and 25: A comparative statement on computational analysis and mRNA regulation upon pathogenic infection. Fish Shellfish. Immunol. 2015, 47, 221–230. [Google Scholar] [CrossRef] [PubMed]
- Liao, Z.; Wan, Q.; Xiao, X.; Ji, J.; Su, J. A systematic investigation on the composition, evolution and expression characteristics of chemokine superfamily in grass carp Ctenopharyngodon idella. Dev. Comp. Immunol. 2018, 82, 72–82. [Google Scholar] [CrossRef]
- Hayes, S.; Waltmann, M.; Donohue, M.; Lye, D.; Vesper, S. Predicting virulence of Aeromonas isolates based on changes in transcription of c-jun and c-fos in human tissue culture cells. J. Appl. Microbiol. 2009, 107, 964–969. [Google Scholar] [CrossRef]
- Tian, L.; Zhou, X.-Q.; Jiang, W.-D.; Liu, Y.; Wu, P.; Jiang, J.; Kuang, S.-Y.; Tang, L.; Tang, W.-N.; Zhang, Y.-A.; et al. Sodium butyrate improved intestinal immune function associated with NF-κB and p38MAPK signalling pathways in young grass carp (Ctenopharyngodon idella). Fish Shellfish. Immunol. 2017, 66, 548–563. [Google Scholar] [CrossRef]
- Banerjee, C.; Goswami, R.; Verma, G.; Datta, M.; Mazumder, S. Aeromonas hydrophila induced head kidney macrophage apoptosis in Clarias batrachus involves the activation of calpain and is caspase-3 mediated. Dev. Comp. Immunol. 2012, 37, 323–333. [Google Scholar] [CrossRef]
- Lu, A.J.; Hu, X.C.; Wang, Y.; Zhu, A.H.; Shen, L.L.; Tian, J.; Feng, Z.Z.; Feng, Z.J. Skin immune response in the zebrafish, Danio rerio (Hamilton), to Aeromonas hydrophila infection: A transcriptional profiling approach. J. Fish Dis. 2015, 38, 137–150. [Google Scholar] [CrossRef]
- Ni, P.-J.; Jiang, W.-D.; Wu, P.; Liu, Y.; Kuang, S.-Y.; Tang, L.; Tang, W.-N.; Zhang, Y.-A.; Zhou, X.-Q.; Feng, L. Dietary low or excess levels of lipids reduced growth performance, and impaired immune function and structure of head kidney, spleen and skin in young grass carp (Ctenopharyngodon idella) under the infection of Aeromonas hydrophila. Fish Shellfish. Immunol. 2016, 55, 28–47. [Google Scholar] [CrossRef] [PubMed]
- Reyes-Becerril, M.; Sanchez, V.; Delgado, K.; Guerra, K.; Velazquez, E.; Ascencio, F.; Angulo, C. Caspase -1, -3, -8 and antioxidant enzyme genes are key molecular effectors following Vibrio parahaemolyticus and Aeromonas veronii infection in fish leukocytes. Immunobiology 2018, 223, 562–576. [Google Scholar] [CrossRef] [PubMed]
- Shelly, A.; Banerjee, C.; Saurav, G.K.; Ray, A.; Rana, V.S.; Raman, R.; Mazumder, S. Aeromonas hydrophila-induced alterations in cytosolic calcium activate pro-apoptotic cPKC-MEK1/2-TNFα axis in infected headkidney macrophages of Clarias gariepinus. Dev. Comp. Immunol. 2017, 76, 392–402. [Google Scholar] [CrossRef] [PubMed]
- Mulder, I.; Wadsworth, S.; Secombes, C. Cytokine expression in the intestine of rainbow trout (Oncorhynchus mykiss) during infection with Aeromonas salmonicida. Fish Shellfish. Immunol. 2007, 23, 747–759. [Google Scholar] [CrossRef] [PubMed]
- Hop, H.T.; Arayan, L.T.; Huy, T.X.N.; Reyes, A.W.B.; Vu, S.H.; Min, W.; Lee, H.J.; Rhee, M.H.; Chang, H.H.; Kim, S. The key role of c-Fos for immune regulation and bacterial dissemination in Brucella infected macrophage. Front. Microbiol. 2018, 8. [Google Scholar] [CrossRef] [PubMed]
- Dos Santos, P.A.; Pereira, A.C.M.; Ferreira, A.F.; Alves, M.A.D.M.; Rosa, A.C.P.; Freitas-Almeida, A.C.; De Freitas-Almeida, A.C. Adhesion, invasion, intracellular survival and cytotoxic activity of strains of Aeromonas spp. in HEp-2, Caco-2 and T-84 cell lines. Antonie Leeuwenhoek 2015, 107, 1225–1236. [Google Scholar] [CrossRef]
- Ghatak, S.; Agarwal, R.; Bhilegaonkar, K. Comparative study of cytotoxicity of Aeromonas spp. on four different cell lines. Comp. Immunol. Microbiol. Infect. Dis. 2006, 29, 233–241. [Google Scholar] [CrossRef]
- Krzyminska, S.; Mokracka, J.; Koczura, R.; Cwiertnia, A.; Kaznowski, A. Aeromonas spp.-mediated cell-contact cytotoxicity is associated with the presence of type III secretion system. Antonie Leeuwenhoek 2012, 101, 243–251. [Google Scholar] [CrossRef] [Green Version]
- Ottaviani, D.; Parlani, C.; Citterio, B.; Masini, L.; Leoni, F.; Canonico, C.; Sabatini, L.; Bruscolini, F.; Pianetti, A. Putative virulence properties of Aeromonas strains isolated from food, environmental and clinical sources in Italy: A comparative study. Int. J. Food Microbiol. 2011, 144, 538–545. [Google Scholar] [CrossRef]
- Albarral, V.; Sanglas, A.; Palau, M.; Miñana-Galbis, D.; Fusté, M.C. Potential pathogenicity of Aeromonas hydrophila complex strains isolated from clinical, food, and environmental sources. Can. J. Microbiol. 2016, 62, 296–306. [Google Scholar] [CrossRef] [Green Version]
- Abdallah, M.M. The cytotoxicity effect of metalloprotease produced and isolated from Aeromonas spp. Iraqi J. Vet. Med. 2019, 42, 41–45. [Google Scholar]
- Dias, C.; Ribeiro, M.; Correia-Branco, A.; Domínguez-Perles, R.; Martel, F.; Saavedra, M.J.; Simões, M. Virulence, attachment and invasion of Caco-2 cells by multidrug-resistant bacteria isolated from wild animals. Microb. Pathog. 2019, 128, 230–235. [Google Scholar] [CrossRef] [PubMed]
- Chen, P.-L.; Wu, C.-J.; Tsai, P.-J.; Tang, H.-J.; Chuang, Y.-C.; Lee, N.-Y.; Lee, C.-C.; Li, C.-W.; Li, M.-C.; Chen, C.-C.; et al. Virulence diversity among bacteremic Aeromonas isolates: Ex vivo, animal, and clinical evidences. PLoS ONE 2014, 9. [Google Scholar] [CrossRef] [PubMed]
- Dong, Y.; Yang, Y.; Liu, J.; Awan, F.; Lu, C.; Liu, Y. Inhibition of Aeromonas hydrophila-induced intestinal inflammation and mucosal barrier function damage in crucian carp by oral administration of Lactococcus lactis. Fish Shellfish. Immunol. 2018, 83, 359–367. [Google Scholar] [CrossRef] [PubMed]
- Peatman, E.; Mohammed, H.; Kirby, A.; Shoemaker, C.A.; Yildirim-Aksoy, M.; Beck, B.H. Mechanisms of pathogen virulence and host susceptibility in virulent Aeromonas hydrophila infections of channel catfish (Ictalurus punctatus). Aquaculture 2018, 482, 1–8. [Google Scholar] [CrossRef]
- Sellegounder, D.; Gupta, Y.R.; Murugananthkumar, R.; Senthilkumaran, B. Enterotoxic effects of Aeromonas hydrophila infection in the catfish, Clarias gariepinus: Biochemical, histological and proteome analyses. Veter. Immunol. Immunopathol. 2018, 204, 1–10. [Google Scholar] [CrossRef]
- Wang, W.; Tan, S.; Luo, J.; Shi, H.; Zhou, T.; Yang, Y.; Jin, Y.; Wang, X.; Niu, D.; Yuan, Z.; et al. GWAS analysis indicated importance of NF-κB signaling pathway in host resistance against motile Aeromonas septicemia disease in catfish. Mar. Biotechnol. 2019, 21, 335–347. [Google Scholar] [CrossRef]
- Fock, W.; Chen, C.; Lam, T.; Sin, Y. Roles of an endogenous serum lectin in the immune protection of blue gourami, Trichogaster trichopterus (Pallus) against Aeromonas hydrophila. Fish Shellfish. Immunol. 2001, 11, 101–113. [Google Scholar] [CrossRef]
- Yu, H.B.; Zhang, Y.L.; Lau, Y.L.; Yao, F.; Vilches, S.; Merino, S.; Tomas, J.M.; Howard, S.P.; Leung, K.Y. Identification and characterization of putative virulence genes and gene clusters in Aeromonas hydrophila PPD134/91. Appl. Environ. Microbiol. 2005, 71, 4469–4477. [Google Scholar] [CrossRef] [Green Version]
- Froquet, R.; Cherix, N.; Burr, S.E.; Frey, J.; Vilches, S.; Tomas, J.M.; Cosson, P. Alternative host model to evaluate Aeromonas virulence. Appl. Environ. Microbiol. 2007, 73, 5657–5659. [Google Scholar] [CrossRef] [Green Version]
- Chen, P.L.; Chen, Y.W.; Ou, C.C.; Lee, T.M.; Wu, C.J.; Ko, W.C.; Chen, C.S. A disease model of muscle necrosis caused by Aeromonas dhakensis infection in Caenorhabditis elegans. Front. Microbiol. 2016, 7, 2058. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.-W.; Ko, W.-C.; Chen, C.-S.; Chen, P.-L. Evaluating virulence and pathogenesis of Aeromonas infection in a Caenorhabditis elegans model. J. Vis. Exp. 2018, e58768. [Google Scholar] [CrossRef] [Green Version]
- Buer, J.; Balling, R. Mice, microbes and models of infection. Nat. Rev. Genet. 2003, 4, 195–205. [Google Scholar] [CrossRef] [PubMed]
- Aravena-Roman, M.; Inglis, T.J.; Henderson, B.; Riley, T.V.; Chang, B.J. Antimicrobial susceptibilities of Aeromonas strains isolated from clinical and environmental sources to 26 antimicrobial agents. Antimicrob. Agents Chemother. 2012, 56, 1110–1112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Codjoe, F.S.; Donkor, E.S. Carbapenem resistance: A review. Med. Sci. 2017, 6, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ndi, O.L.; Barton, M.D. Incidence of class 1 integron and other antibiotic resistance determinants in Aeromonas spp. from rainbow trout farms in Australia. J. Fish Dis. 2011, 34, 589–599. [Google Scholar] [CrossRef]
- Dias, C.; Serra, C.R.; Simoes, L.C.; Simões, M.; Martinez-Murcia, A.; Saavedra, M.J. Extended-spectrum β-lactamase and carbapenemase-producing Aeromonas species in wild animals from Portugal. Veter. Rec. 2014, 174, 532. [Google Scholar]
- Fosse, T.; Giraud-Morin, C.; Madinier, I. Phenotypes of beta-lactam resistance in the genus Aeromonas. Pathol. Boil. 2003, 51, 290–296. [Google Scholar] [CrossRef]
- Cespedes, J.S.; Figueras, M.J.; Aspiroz, C.; Aldea, M.J.; Toledo, M.; Alperi, A.; Marco, F.; Vilá, J. Development of imipenem resistance in an Aeromonas veronii biovar sobria clinical isolate recovered from a patient with cholangitis. J. Med. Microbiol. 2009, 58, 451–455. [Google Scholar] [CrossRef] [Green Version]
- Dias, C.; Borges, A.; Saavedra, M.J.; Simoes, M. Biofilm formation and multidrug-resistant Aeromonas spp. from wild animals. J. Glob. Antimicrob. Resist. 2018, 12, 227–234. [Google Scholar] [CrossRef]
- Hossain, S.; De Silva, B.; Dahanayake, P.; Heo, G.-J. Characterization of virulence properties and multi-drug resistance profiles in motile Aeromonas spp. isolated from zebrafish (Danio rerio). Lett. Appl. Microbiol. 2018, 67, 598–605. [Google Scholar] [CrossRef] [PubMed]
- Hossain, S.; De Silva, B.; Wickramanayake, M.; Dahanayake, P.; Wimalasena, S.; Heo, G. Incidence of antimicrobial resistance genes and class 1 integron gene cassettes in multidrug-resistant motile Aeromonas sp. isolated from ornamental guppy (Poecilia reticulata). Lett. Appl. Microbiol. 2019, 69, 2–10. [Google Scholar] [CrossRef] [PubMed]
- Ramadan, H.; Ibrahim, N.; Samir, M.; El-Moaty, A.A.; Gad, T. Aeromonas hydrophila from marketed mullet (Mugil cephalus) in Egypt: PCR characterization of β -lactam resistance and virulence genes. J. Appl. Microbiol. 2018, 124, 1629–1637. [Google Scholar] [CrossRef] [PubMed]
- Scarano, C.; Piras, F.; Virdis, S.; Ziino, G.; Nuvoloni, R.; Dalmasso, A.; De Santis, E.; Spanu, C. Antibiotic resistance of Aeromonas ssp. strains isolated from Sparus aurata reared in Italian mariculture farms. Int. J. Food Microbiol. 2018, 284, 91–97. [Google Scholar] [CrossRef] [PubMed]
- Wimalasena, S.H.M.P.; Shin, G.-W.; Hossain, S.; Heo, G.-J. Potential enterotoxicity and antimicrobial resistance pattern of Aeromonas species isolated from pet turtles and their environment. J. Veter. Med. Sci. 2017, 79, 921–926. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cattoir, V.; Poirel, L.; Aubert, C.; Soussy, C.-J.; Nordmann, P. Unexpected occurrence of plasmid-mediated quinolone resistance determinants in environmental Aeromonas spp. Emerg. Infect. Dis. 2008, 14, 231–237. [Google Scholar] [CrossRef]
- Cespedes, J.S.; Blasco, M.D.; Marti, S.; Alba, V.; Alcalde, E.; Esteve, C.; Vila, J. Plasmid-mediated QnrS2 determinant from a clinical Aeromonas veronii isolate. Antimicrob. Agents Chemother. 2008, 52, 2990–2991. [Google Scholar] [CrossRef] [Green Version]
- Piotrowska, M.; Popowska, M. Insight into the mobilome of Aeromonas strains. Front. Microbiol. 2015, 6, 494. [Google Scholar] [CrossRef] [Green Version]
- Vega-Sánchez, V.; Latif-Eugenín, F.; Soriano-Vargas, E.; Beaz-Hidalgo, R.; Figueras, M.J.; Aguilera-Arreola, M.G.; Castro-Escarpulli, G. Re-identification of Aeromonas isolates from rainbow trout and incidence of class 1 integron and β-lactamase genes. Veter. Microbiol. 2014, 172, 528–533. [Google Scholar] [CrossRef]
- Piotrowska, M.; Przygodzińska, D.; Matyjewicz, K.; Popowska, M. Occurrence and variety of β-Lactamase genes among Aeromonas spp. isolated from urban wastewater treatment plant. Front. Microbiol. 2017, 8, 863. [Google Scholar] [CrossRef] [Green Version]




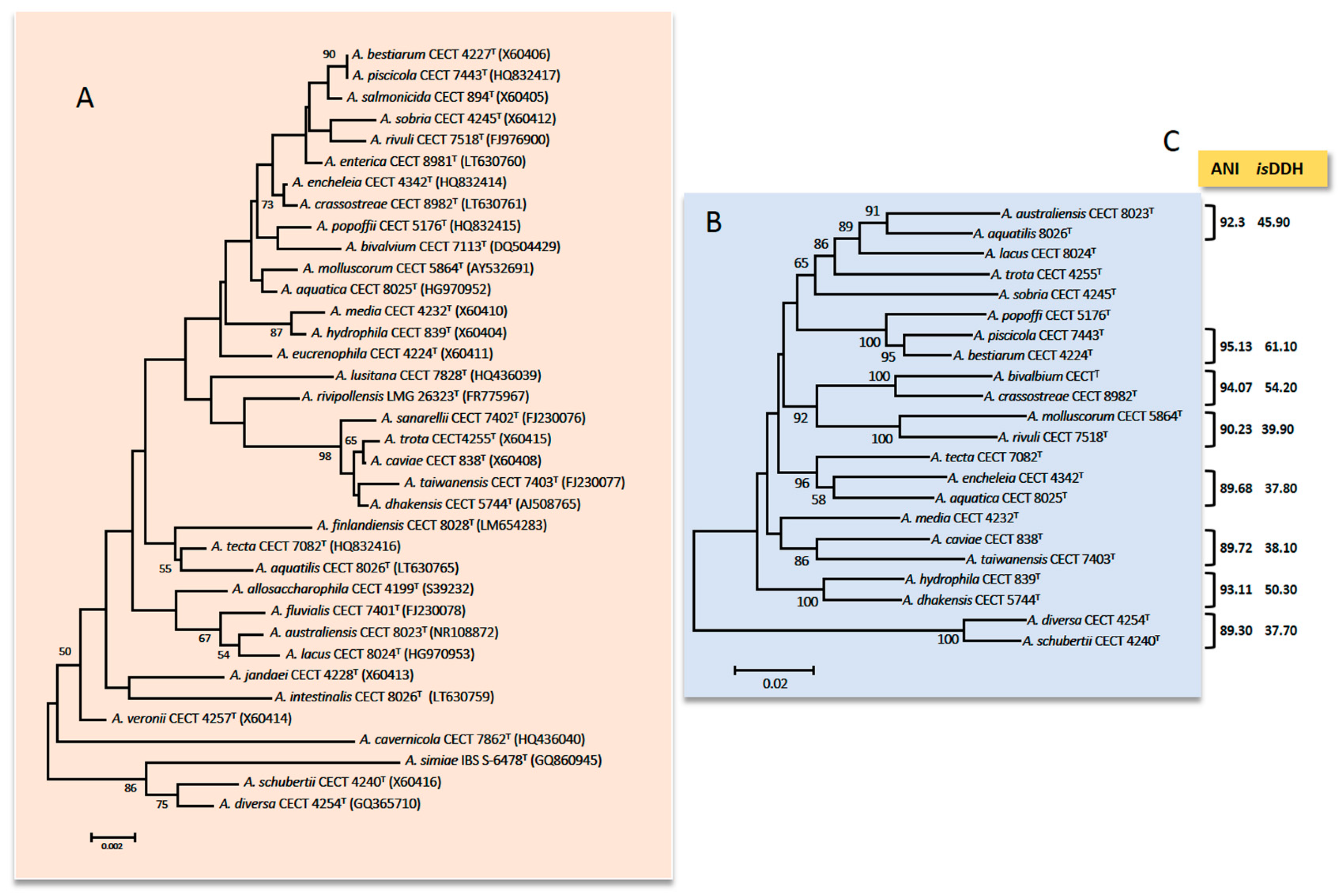{kind=link}
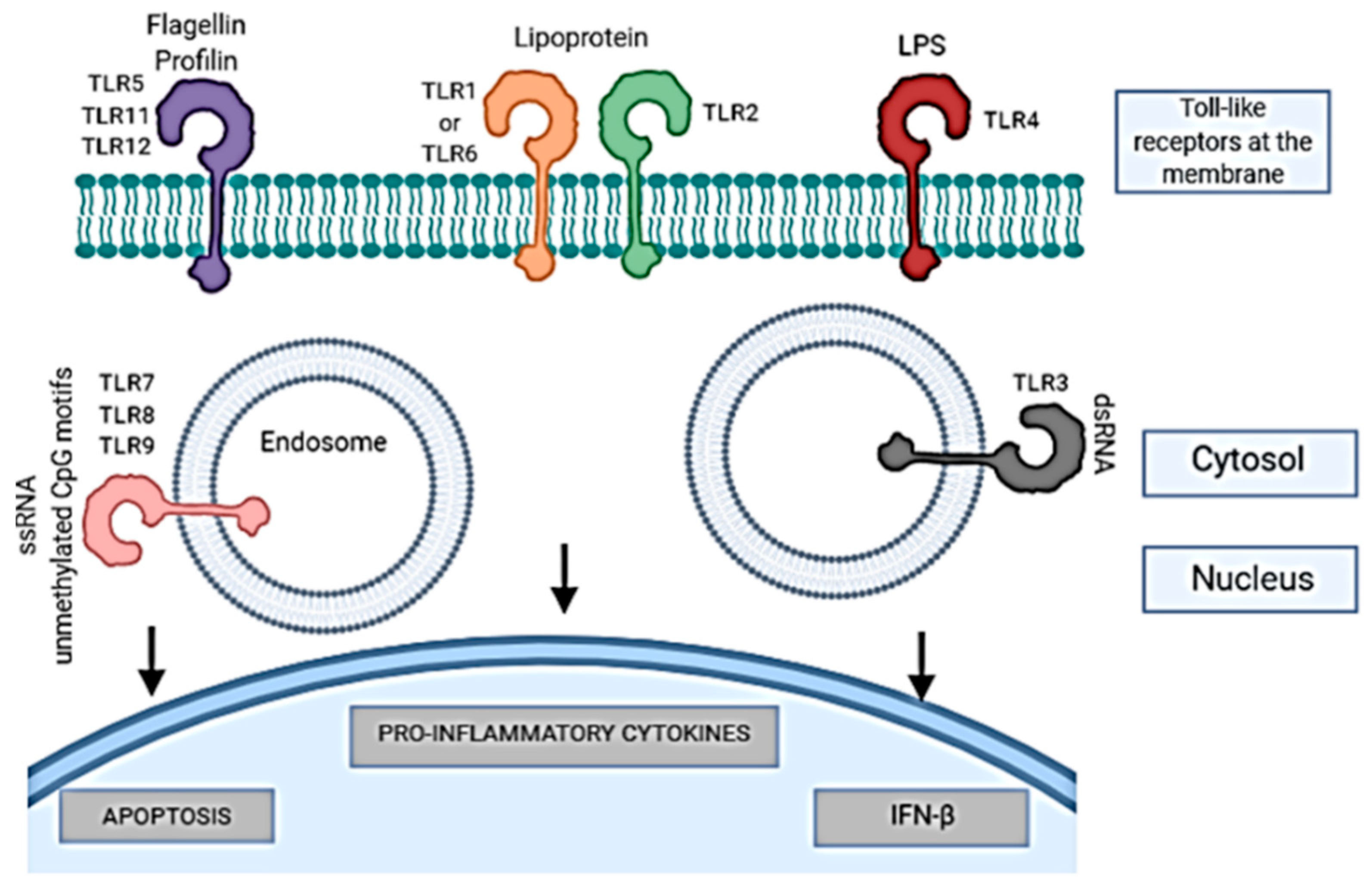{kind=link}
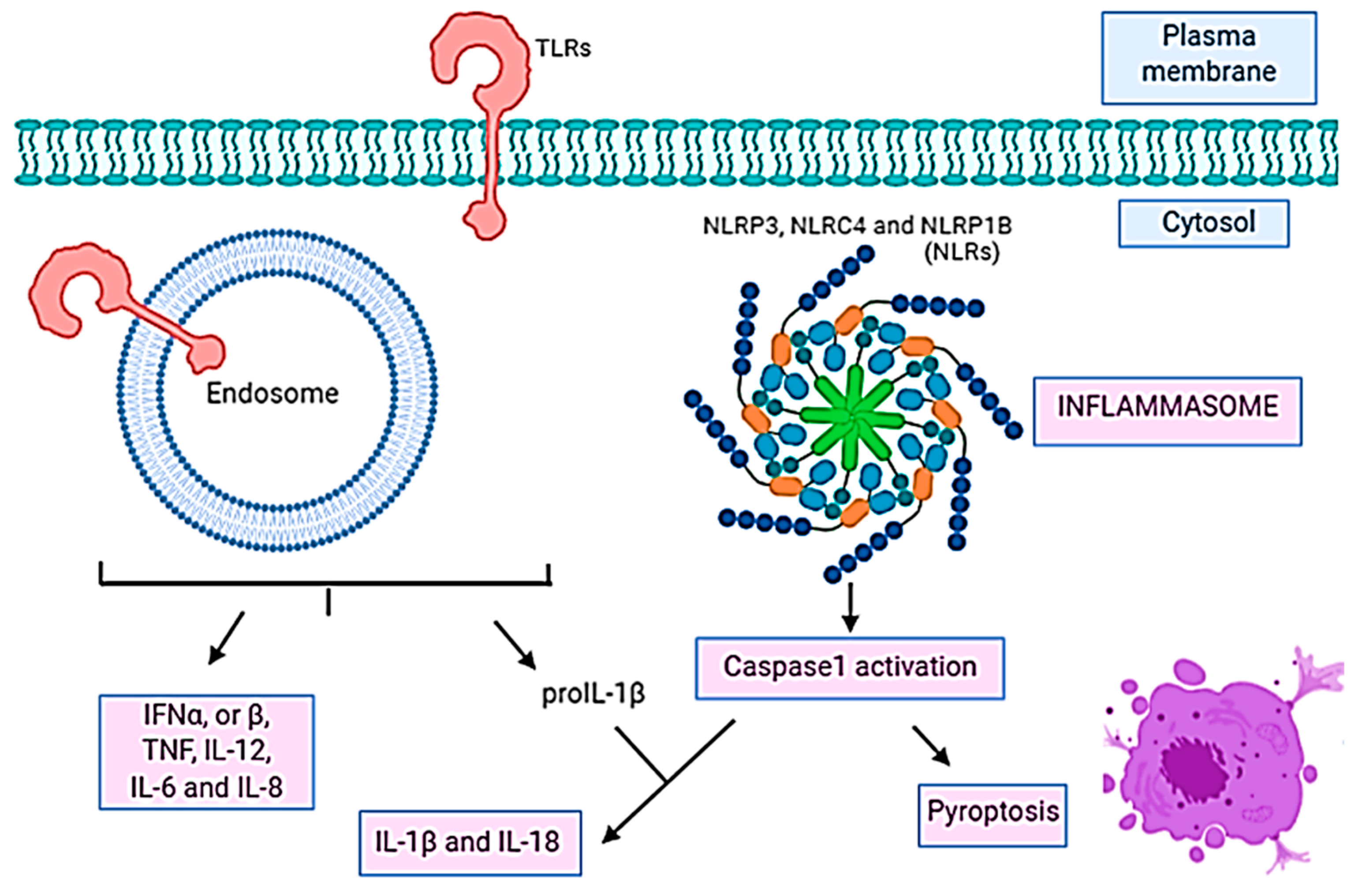{kind=link}
| Species | Faeces | Wound | Blood | Respiratory Tract b | Urine | Peritoneal Dialysate | Bilis | Ascitic | Abscess | Other c Fluid | Total |
|---|---|---|---|---|---|---|---|---|---|---|---|
| A. caviae | 446 | 43 | 147 | 11 | 12 | 11 | 11 | 2 | 3 | 4 | 690 (37.26) |
| A. veronii | 231 | 49 | 125 | 12 | 1 | 1 | 6 | 7 | 1 | 2 | 435 (23.49) |
| A. dhakensisd | 111 | 133 | 111 | 4 | 6 | 11 | 11 | 2 | 1 | 9 | 399 (21.54) |
| A. hydrophila | 69 | 96 | 45 | 9 | 5 | 8 | 3 | 1 | 2 | 4 | 242 (13.07) |
| A. media | 32 | 3 | 6 | 1 | 42 (2.27) | ||||||
| A. trota | 2 | 2 | 1 | 1 | 6 (0.27) | ||||||
| A. taiwanensis | 2 | 3 | 1 | 1 | 7 (0.32) | ||||||
| A. salmonicida | 1 | 2 | 1 | 4 (0.21) | |||||||
| A. jandaei | 3 | 2 | 1 | 1 | 1 | 8 (0.43) | |||||
| A. sanarellii | 1 | 4 | 5 (0.27) | ||||||||
| A. allosaccharophila | 2 | 1 | 3 (0.16) | ||||||||
| A. tecta | 2 | 2 (0.11) | |||||||||
| A. diversa | 2 | 2 (0.11) | |||||||||
| A. schubertii | 1 | 1 | 2 (0.11) | ||||||||
| A. bestiarum | 1 | 1 | 1 | 1 | 4 (0.21) | ||||||
| A. popoffii | 1 | 1 (0.05) | |||||||||
| A. intestinalise | 1 | 1 (0.05) | |||||||||
| A. entericae | 1 | 1 (0.05) | |||||||||
| Total | 905 | 341 | 439 | 37 | 27 | 32 | 32 | 12 | 8 | 20 | 1852 |
| Proteins or Process Studied | Host | Aeromonas sp. | Reference |
|---|---|---|---|
| Toll-like receptors (TLRs) (TLR1, TLR2, TLR3, TLR4, TLR5, TLR7, TLR8, TLR9, TLR18, TLR19, TLR20, TLR22) | Fish, cells (EPC, HKLs, Hep-2) | A. hydrophila, A. salmonicida, A. veronii | [21,24,282,283,286,287,288,289,290,291,292] |
| Cytokines (TNF-α, TNF- β, IFN-γ, IL-6, IL-8, IL-10) | Fish, mice, cells (Hep-2, HKLs, RAW 264.7, HeLa) | A. hydrophila, A. salmonicida, A. dhakensis, A. veronii/A. hydrophila | [259,289,290,293,294,295,296] |
| Chemokines (CC-Chem14, CsCC-Chem20, CsCC-Chem25, CCL3, CCL5) | Fish, mice, cells (HeLa, RAW 264.7) | A. hydrophila, A. dhakensis | [259,295,297,298] |
| Transcription factors (JUN, RELA) | Fish, cells (Caco-2) | A. hydrophila, A. salmonicida, A. veronii, A. bestiarum, A. allosaccharophila | [299,300] |
| Apoptosis (TP53, CASP3) | Fish, mice, cells (HeLa, RAW 264.7, macrophages from fish, SLs, HKLs) | A. hydrophila, A. veronii | [295,301,302,303,304,305] |
| Pyroptosis (NLRP3, NLRC4, IL-1β) | Mice, macrophages from mice | A. hydrophila, A. veronii | [22,23] |
| Cell Line | Study | Aeromonas | Reference |
|---|---|---|---|
| HEp-2, Vero | Adhesion and cytotoxicity | A. hydrophila. A. salmonicida, A. veronii, A. bestiarum, A. schuberti, A. eucrenophila, A. encheleia, A. jandaei, A. sobria, A. caviae, A. trota, A. media | [311] |
| HEp-2, CHO | Cell-contact cytotoxicity | A. hydrophila, A. caviae, A. veronii | [310] a |
| C2C12 | Cytotoxicity | A. hydrophila, A. dhakensis | [68] |
| HEp-2 | Adhesion, biofilm formation, and immune stimulation | A. hydrophila | [289] |
| HEp-2, Caco-2, T-84 | Adhesion, invasion, and cytotoxicity | A. hydrophila, A. caviae | [308] a |
| RAW 264.7 | Role of mixed infections in Necrotizing fasciitis | A. hydrophila | [187] |
| Caco-2 | Adhesion and cytopathic effect | A. hydrophila, A. dhakensis, A. bestiarum, A. piscicola, A. salmonicida | [312] |
| HepG2, WLR-68 | Cytotoxicity effect of metalloprotease | A. hydrophila | [313] a |
| Caco-2 | Adhesion, invasion, and cytotoxicity | A. salmonicida | [314] |
| J744.1 | Role of metallochaperone HypA | A. hydrophila | [273] |
| RAW 264.7 | Role of mixed infections in Necrotizing fasciitis | A. hydrophila | [188] |
| Model | Study | Aeromonas | Reference |
|---|---|---|---|
| Mice (Mus musculus) | Virulence factors, role of mixed infections in necrotizing fasciitis | A. hydrophila, A. dhakensis, A. caviae, A. veronii, A. salmonicida | [16,17,186,187,188,315] |
| Catfish (Clarias gariepinus, Ictalurus punctatus, Hypophthalmichthys molitrix) | Enterotoxic effects, virulence mechanism, transcriptome, aerolysin activity | A. hydrophila, A. veronii | [316,317,318,319] |
| Blue gourami (Trichogaster trichopterus) | Septicemia, immune responses | A. hydrophila | [320,321] |
| Zebrafish (Danio rerio) | Immune response, role of mixed infections in the virulence | A. hydrophila/A. veronii coinfection, A. hydrophila | [17,24,293] |
| Slime mold (Trichogaster tricopterus) | Pathogenicity | A. salmonicida, A. hydrophila | [322] |
| Nematode (Caenorhabditis elegans) | Virulence, immune response, necrosis | A. hydrophila, A. dhakensis, A. veronii, A, caviae | [183,323,324] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fernández-Bravo, A.; Figueras, M.J. An Update on the Genus Aeromonas: Taxonomy, Epidemiology, and Pathogenicity. Microorganisms 2020, 8, 129. https://doi.org/10.3390/microorganisms8010129
Fernández-Bravo A, Figueras MJ. An Update on the Genus Aeromonas: Taxonomy, Epidemiology, and Pathogenicity. Microorganisms. 2020; 8(1):129. https://doi.org/10.3390/microorganisms8010129
Chicago/Turabian StyleFernández-Bravo, Ana, and Maria José Figueras. 2020. "An Update on the Genus Aeromonas: Taxonomy, Epidemiology, and Pathogenicity" Microorganisms 8, no. 1: 129. https://doi.org/10.3390/microorganisms8010129
APA StyleFernández-Bravo, A., & Figueras, M. J. (2020). An Update on the Genus Aeromonas: Taxonomy, Epidemiology, and Pathogenicity. Microorganisms, 8(1), 129. https://doi.org/10.3390/microorganisms8010129