Competence for Natural Transformation Is Common among Clinical Strains of Resistant Acinetobacter spp.

 , ,
, ,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Isolates and Growth Conditions
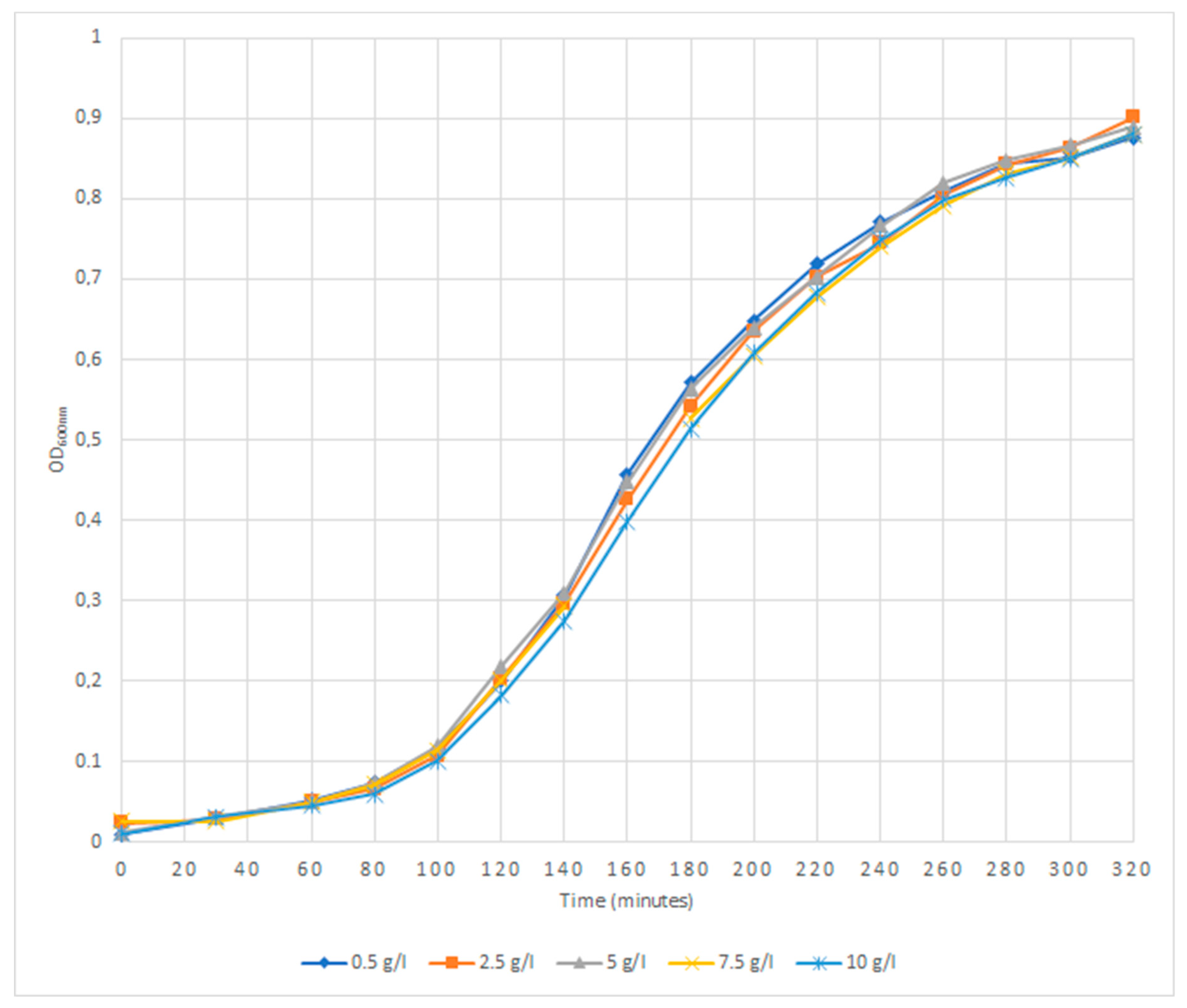
2.2. Growth Curve Under Different Sodium Chloride Concentrations
2.3. Natural Transformation
2.4. Transformants Confirmation
3. Results and Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Smalla, K.; Cook, K.; Djordjevic, S.P.; Klumper, U.; Gillings, M. Environmental dimensions of antibiotic resistance: Assessment of basic science gaps. FEMS Microbiol. Ecol. 2018, 94, fiy195. [Google Scholar] [CrossRef] [PubMed]
- Rice, L.B. Federal funding for the study of antimicrobial resistance in nosocomial pathogens: No ESKAPE. J. Infect. Dis. 2008, 197, 1079–1081. [Google Scholar] [CrossRef] [PubMed]
- Bergogne-Berezin, E.; Joly-Guillou, M.L. An underestimated nosocomial pathogen Acinetobacter calcoaceticus. J. Antimicrob. Chemother. 1985, 16, 535–538. [Google Scholar] [CrossRef] [PubMed]
- Da Silva, G.J.; Domingues, S. Insights on the horizontal gene transfer of carbapenemase determinants in the opportunistic pathogen Acinetobacter baumannii. Microorganisms 2016, 4, 23. [Google Scholar] [CrossRef] [PubMed]
- Tacconelli, E.; Magrini, N. Global Priority List of Antibiotic-Resistant Bacteria to Guide Research, Discovery, and Development of New Antibiotics; World Health Organization: Geneva, Switzerland, 2017. [Google Scholar]
- von Wintersdorff, C.J.; Penders, J.; van Niekerk, J.M.; Mills, N.D.; Majumder, S.; van Alphen, L.B.; Savelkoul, P.H.; Wolffs, P.F. Dissemination of antimicrobial resistance in microbial ecosystems through horizontal gene transfer. Front. Microbiol. 2016, 7, 173. [Google Scholar] [CrossRef] [PubMed]
- Domingues, S.; Nielsen, K.M. Horizontal gene transfer: Uptake of extracellular DNA by bacteria. In Reference Module in Biomedical Sciences; Elsevier: Amsterdam, The Netherlands, 2016. [Google Scholar]
- Barbe, V.; Vallenet, D.; Fonknechten, N.; Kreimeyer, A.; Oztas, S.; Labarre, L.; Cruveiller, S.; Robert, C.; Duprat, S.; Wincker, P.; et al. Unique features revealed by the genome sequence of Acinetobacter sp. ADP1, a versatile and naturally transformation competent bacterium. Nucleic Acids Res. 2004, 32, 5766–5779. [Google Scholar] [CrossRef] [PubMed]
- Domingues, S.; Harms, K.; Fricke, W.F.; Johnsen, P.J.; da Silva, G.J.; Nielsen, K.M. Natural transformation facilitates transfer of transposons, integrons and gene cassettes between bacterial species. PLoS Pathog. 2012, 8, e1002837. [Google Scholar] [CrossRef] [PubMed]
- Ramirez, M.S.; Don, M.; Merkier, A.K.; Bistue, A.J.; Zorreguieta, A.; Centron, D.; Tolmasky, M.E. Naturally competent Acinetobacter baumannii clinical isolate as a convenient model for genetic studies. J. Clin. Microbiol. 2010, 48, 1488–1490. [Google Scholar] [CrossRef] [PubMed]
- Carruthers, M.D.; Harding, C.M.; Baker, B.D.; Bonomo, R.A.; Hujer, K.M.; Rather, P.N.; Munson, R.S., Jr. Draft genome sequence of the clinical isolate Acinetobacter nosocomialis strain M2. Genome Announc. 2013, 1, e00906-13. [Google Scholar] [CrossRef] [PubMed]
- Harding, C.M.; Tracy, E.N.; Carruthers, M.D.; Rather, P.N.; Actis, L.A.; Munson, R.S., Jr. Acinetobacter baumannii strain M2 produces type IV pili which play a role in natural transformation and twitching motility but not surface-associated motility. MBio 2013, 4, e00360-13. [Google Scholar] [CrossRef] [PubMed]
- Wilharm, G.; Piesker, J.; Laue, M.; Skiebe, E. DNA uptake by the nosocomial pathogen Acinetobacter baumannii occurs during movement along wet surfaces. J. Bacteriol. 2013, 195, 4146–4153. [Google Scholar] [CrossRef] [PubMed]
- Godeux, A.S.; Lupo, A.; Haenni, M.; Guette-Marquet, S.; Wilharm, G.; Laaberki, M.H.; Charpentier, X. Fluorescence-based detection of natural transformation in drug resistant Acinetobacter baumannii. J. Bacteriol. 2018, 200, e00181-18. [Google Scholar] [CrossRef] [PubMed]
- Wilharm, G.; Skiebe, E.; Higgins, P.G.; Poppel, M.T.; Blaschke, U.; Leser, S.; Heider, C.; Heindorf, M.; Brauner, P.; Jackel, U.; et al. Relatedness of wildlife and livestock avian isolates of the nosocomial pathogen Acinetobacter baumannii to lineages spread in hospitals worldwide. Environ. Microbiol. 2017, 19, 4349–4364. [Google Scholar] [CrossRef] [PubMed]
- Traglia, G.M.; Quinn, B.; Schramm, S.T.; Soler Bistue, A.; Ramirez, M.S. Serum albumin and Ca2+ are natural competence inducers in the human pathogen Acinetobacter baumannii. Antimicrob. Agents Chemother. 2016, 60, 4920–4929. [Google Scholar] [CrossRef] [PubMed]
- Quinn, B.; Martinez, J.; Liu, C.; Nguyen, M.; Ramirez, M.S. The effect of sub-inhibitory concentrations of antibiotics on natural transformation in Acinetobacter baumannii. Int. J. Antimicrob. Agents 2018, 51, 809–810. [Google Scholar] [CrossRef] [PubMed]
- La Scola, B.; Gundi, V.A.; Khamis, A.; Raoult, D. Sequencing of the rpoB gene and flanking spacers for molecular identification of Acinetobacter species. J. Clin. Microbiol. 2006, 44, 827–832. [Google Scholar] [CrossRef] [PubMed]
- Wright, M.S.; Haft, D.H.; Harkins, D.M.; Perez, F.; Hujer, K.M.; Bajaksouzian, S.; Benard, M.F.; Jacobs, M.R.; Bonomo, R.A.; Adams, M.D. New insights into dissemination and variation of the health care-associated pathogen Acinetobacter baumannii from genomic analysis. MBio 2014, 5, e00963-13. [Google Scholar] [CrossRef] [PubMed]
- Nemec, A.; Krizova, L.; Maixnerova, M.; van der Reijden, T.J.; Deschaght, P.; Passet, V.; Vaneechoutte, M.; Brisse, S.; Dijkshoorn, L. Genotypic and phenotypic characterization of the Acinetobacter calcoaceticus-Acinetobacter baumannii complex with the proposal of Acinetobacter pittii sp. nov. (formerly Acinetobacter genomic species 3) and Acinetobacter nosocomialis sp. nov. (formerly Acinetobacter genomic species 13TU). Res. Microbiol. 2011, 162, 393–404. [Google Scholar] [PubMed]
- Ray, J.L.; Harms, K.; Wikmark, O.G.; Starikova, I.; Johnsen, P.J.; Nielsen, K.M. Sexual isolation in Acinetobacter baylyi is locus-specific and varies 10,000-fold over the genome. Genetics 2009, 182, 1165–1181. [Google Scholar] [CrossRef] [PubMed]
- Martinez, J.; Liu, C.; Rodman, N.; Fernandez, J.S.; Barberis, C.; Sieira, R.; Perez, F.; Bonomo, R.A.; Ramirez, M.S. Human fluids alter DNA-acquisition in Acinetobacter baumannii. Diagn Microbiol. Infect. Dis. 2018. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Isolate | Species | Year of Isolation | Hospital/Region | Natural Competence a |
|---|---|---|---|---|
| 013 | A. nosocomialis | 1992 | HUC/Coimbra | + |
| 015 | A. baumannii | 1994 | HUC/Coimbra | + |
| 113 | A. baumannii | 1998 | HUC/Coimbra | + |
| 118 | A. bereziniae | 1998 | HUC/Coimbra | - |
| 121 | A. baumannii | 1998 | HUC/Coimbra | + |
| 129 | A. baumannii | 1998 | HUC/Coimbra | + |
| 132 | A. baumannii | 1998 | HUC/Coimbra | + |
| 138022 F1Ev | n.d. | 2008 | HES/Évora | - |
| 144417 M1Ev | A. baumannii | 2008 | HES/Évora | + |
| 213 | n.d. | 2004 | HSAC/Lisbon | - |
| 241 | A. baumannii | 2005 | HSAC/Lisbon | + |
| 245 | A. baumannii | 2005 | HSAC/Lisbon | - |
| 274 | A. baumannii | 2006 | HSAC/Lisbon | - |
| 292 | A. baumannii | 2006 | HSAC/Lisbon | + |
| 319 | A. baumannii | 2007 | HSAC/Lisbon | + |
| 326744 C1Ev | A. nosocomialis | 2008 | HES/Évora | + |
| 3605 | n.d. | 1994 | Porto | - |
| 3625 | A. baumannii | 1995 | Porto | + |
| 532331 A1Ev | A. baumannii | 2008 | HES/Évora | + |
| 545663 F2Ev | A. baumannii | 2008 | HES/Évora | - |
| 065 | Acinetobacter sp. | 1999 | HSM/Lisbon | + |
| 65FFC | A. baumannii | 1998 | HUC/Coimbra | - |
| NaCl Concentration | Mean Number of Transformants (CFU) ± SD | Mean Number of Total Cells (CFU) ± SD | Transformation Frequency |
|---|---|---|---|
| 0.5 g/L | (4.73 ± 3.11) × 104 | (1.14 ± 0.65) × 108 | 4.1 × 10−5 |
| 2.5 g/L | (3.65 ± 4.30) × 104 | (6.38 ± 3.30) × 108 | 5.7 × 10−5 |
| 5 g/L | (9.11 ± 6.68) × 103 | (5.27 ± 3.04) × 108 | 1.7 × 10−5 |
| 7.5 g/L | (5.28 ± 56.1) × 102 | (8.14 ± 8.61) × 108 | 6.5 × 10−7 |
| 10 g/L | (1.35 ± 1.58) × 102 | (7.88 ± 2.32) × 108 | 1.7 × 10−7 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Domingues, S.; Rosário, N.; Cândido, Â.; Neto, D.; Nielsen, K.M.; Da Silva, G.J. Competence for Natural Transformation Is Common among Clinical Strains of Resistant Acinetobacter spp. Microorganisms 2019, 7, 30. https://doi.org/10.3390/microorganisms7020030
Domingues S, Rosário N, Cândido Â, Neto D, Nielsen KM, Da Silva GJ. Competence for Natural Transformation Is Common among Clinical Strains of Resistant Acinetobacter spp. Microorganisms. 2019; 7(2):30. https://doi.org/10.3390/microorganisms7020030
Chicago/Turabian StyleDomingues, Sara, Natasha Rosário, Ângela Cândido, Daniela Neto, Kaare M. Nielsen, and Gabriela J. Da Silva. 2019. "Competence for Natural Transformation Is Common among Clinical Strains of Resistant Acinetobacter spp." Microorganisms 7, no. 2: 30. https://doi.org/10.3390/microorganisms7020030
APA StyleDomingues, S., Rosário, N., Cândido, Â., Neto, D., Nielsen, K. M., & Da Silva, G. J. (2019). Competence for Natural Transformation Is Common among Clinical Strains of Resistant Acinetobacter spp. Microorganisms, 7(2), 30. https://doi.org/10.3390/microorganisms7020030