The Influence of Modernization and Disease on the Gastric Microbiome of Orang Asli, Myanmars and Modern Malaysians

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Screening of H. pylori Infections in Orang Asli
2.3. Collection of Gastric Biopsies
2.4. Illumina 16S rRNA Gene Sequencing
2.5. Data Analysis
2.6. Statistical Analysis
3. Results
3.1. H. pylori Prevalence among Orang Asli
3.2. The Gastric Microbial Community of H. pylori-Positive Orang Asli, Myanmar and Modern Malaysian Subjects
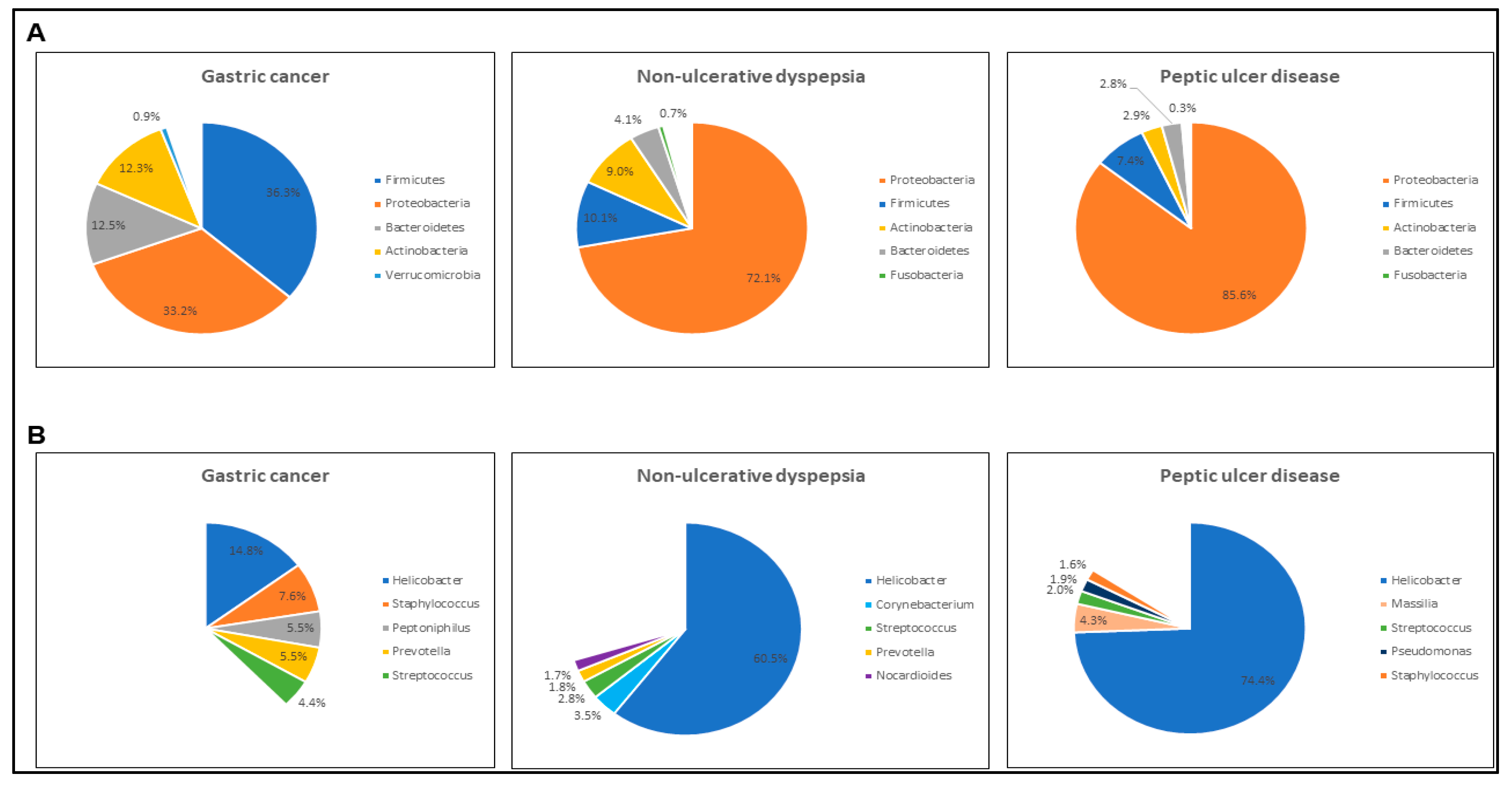
3.3. Gastric Diseases Altered the Microbial Community Structure in Modern Malaysian Patients
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Lederberg, J.; McCray, A.T. ‘Ome Sweet ‘Omics-A Genealogical Treasury of Words. Scientist 2001, 15, 8. [Google Scholar]
- Pei, Z.; Bini, E.J.; Yang, L.; Zhou, M.; Francois, F.; Blaser, M.J. Bacterial biota in the human distal esophagus. Proc. Natl. Acad. Sci. USA 2004, 101, 4250–4255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aas, J.A.; Paster, B.J.; Stokes, L.N.; Olsen, I.; Dewhirst, F.E. Defining the normal bacterial flora of the oral cavity. J. Clin. Microbiol. 2005, 43, 5721–5732. [Google Scholar] [CrossRef] [PubMed]
- Verhelst, R.; Verstraelen, H.; Claeys, G.; Verschraegen, G.; Delanghe, J.; Van Simaey, L.; De Ganck, C.; Temmerman, M.; Vaneechoutte, M. Cloning of 16S rRNA genes amplified from normal and disturbed vaginal microflora suggests a strong association between Atopobium vaginae, Gardnerella vaginalis and bacterial vaginosis. BMC Microbiol. 2004, 4, 16. [Google Scholar] [CrossRef] [PubMed]
- Nardone, G.; Compare, D. The human gastric microbiota: Is it time to rethink the pathogenesis of stomach diseases? United Eur. Gastroenterol. J. 2015, 3, 255–260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marshall, B.J.; Warren, J.R. Unidentified curved bacilli in the stomach of patients with gastritis and peptic ulceration. Lancet 1984, 1, 1311–1315. [Google Scholar] [CrossRef]
- Yang, I.; Woltemate, S.; Piazuelo, M.B.; Bravo, L.E.; Yepez, M.C.; Romero-Gallo, J.; Delgado, A.G.; Wilson, K.T.; Peek, R.M.; Correa, P.; et al. Different gastric microbiota compositions in two human populations with high and low gastric cancer risk in Colombia. Sci. Rep. 2016, 6, 18594. [Google Scholar] [CrossRef]
- Yatsunenko, T.; Rey, F.E.; Manary, M.J.; Trehan, I.; Dominguez-Bello, M.G.; Contreras, M.; Magris, M.; Hidalgo, G.; Baldassano, R.N.; Anokhin, A.P.; et al. Human gut microbiome viewed across age and geography. Nature 2012, 486, 222–227. [Google Scholar] [CrossRef]
- De Filippo, C.; Cavalieri, D.; Di Paola, M.; Ramazzotti, M.; Poullet, J.B.; Massart, S.; Collini, S.; Pieraccini, G.; Lionetti, P. Impact of diet in shaping gut microbiota revealed by a comparative study in children from Europe and rural Africa. Proc. Natl. Acad. Sci. USA 2010, 107, 14691–14696. [Google Scholar] [CrossRef] [Green Version]
- Schnorr, S.L.; Candela, M.; Rampelli, S.; Centanni, M.; Consolandi, C.; Basaglia, G.; Turroni, S.; Biagi, E.; Peano, C.; Severgnini, M.; et al. Gut microbiome of the Hadza hunter-gatherers. Nat. Commun. 2014, 5, 3654. [Google Scholar] [CrossRef]
- Suzuki, T.A.; Worobey, M. Geographical variation of human gut microbial composition. Biol. Lett. 2014, 10, 20131037. [Google Scholar] [CrossRef] [PubMed]
- The World Factbook. Available online: https://www.cia.gov/library/publications/the-world-factbook/ (accessed on 31 January 2019).
- UNICEF. Drinking Water, Sanitation and Hygiene Database; The WHO/UNICEF: Geneva, Switzerland, 2017. [Google Scholar]
- Nicholas, C.; International Work Group for Indigenous Affairs (IWGIA); Center for Orang Asli Concerns (COAC). The Orang Asli and the Contest for Resources: Indigenous Politics, Development, and Identity in Peninsular Malaysia; International Work Group for Indigenous Affairs: Copenhagen, Denmark; Center for Orang Asli Concerns: Subang Jaya, Malaysia, 2000. [Google Scholar]
- Ching, C.C.S.; Ross, V. Preliminary Exploration of a Semai Musician’s Transmission of Indigenous Musical Traditions in Peninsular Malaysia. Pertanika J. Soc. Sci. Humanit. 2015, 23, 20. [Google Scholar]
- Masron, T.; Masami, F.; Ismail, N. Orang Asli in Peninsular Malaysia: Population, Spatial Distribution and Socio-Economic Condition. J. Ritsumeikan Soc. Sci. Hum. 2013, 6, 75–115. [Google Scholar]
- Klindworth, A.; Pruesse, E.; Schweer, T.; Peplies, J.; Quast, C.; Horn, M.; Glockner, F.O. Evaluation of general 16S ribosomal RNA gene PCR primers for classical and next-generation sequencing-based diversity studies. Nucleic Acids Res. 2013, 41, e1. [Google Scholar] [CrossRef] [PubMed]
- Bartram, A.K.; Lynch, M.D.; Stearns, J.C.; Moreno-Hagelsieb, G.; Neufeld, J.D. Generation of multimillion-sequence 16S rRNA gene libraries from complex microbial communities by assembling paired-end illumina reads. Appl. Environ. Microbiol. 2011, 77, 3846–3852. [Google Scholar] [CrossRef] [PubMed]
- 16S Metagenomic Sequencing Library Preparation. Available online: https://support.illumina.com/documents/documentation/chemistry_documentation/16s/16s-metagenomic-library-prep-guide-15044223-b.pdf (accessed on 31 January 2019).
- Parikh, H.I.; Koparde, V.N.; Bradley, S.P.; Buck, G.A.; Sheth, N.U. MeFiT: Merging and filtering tool for illumina paired-end reads for 16S rRNA amplicon sequencing. BMC Bioinform. 2016, 17, 491. [Google Scholar] [CrossRef] [PubMed]
- Albanese, D.; Fontana, P.; De Filippo, C.; Cavalieri, D.; Donati, C. MICCA: A complete and accurate software for taxonomic profiling of metagenomic data. Sci Rep. 2015, 5, 9743. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.; Lin, H.; Revanna, K.; Dong, Q. A Bayesian taxonomic classification method for 16S rRNA gene sequences with improved species-level accuracy. BMC Bioinform. 2017, 18, 247. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Pena, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [Green Version]
- Dabdoub, S.M.; Fellows, M.L.; Paropkari, A.D.; Mason, M.R.; Huja, S.S.; Tsigarida, A.A.; Kumar, P.S. PhyloToAST: Bioinformatics tools for species-level analysis and visualization of complex microbial datasets. Sci Rep. 2016, 6, 29123. [Google Scholar] [CrossRef] [Green Version]
- Iwai, S.; Weinmaier, T.; Schmidt, B.L.; Albertson, D.G.; Poloso, N.J.; Dabbagh, K.; DeSantis, T.Z. Piphillin: Improved Prediction of Metagenomic Content by Direct Inference from Human Microbiomes. PLoS ONE 2016, 11, e0166104. [Google Scholar] [CrossRef] [PubMed]
- Kanehisa, M.; Goto, S. KEGG: Kyoto encyclopedia of genes and genomes. Nucleic Acids Res. 2000, 28, 27–30. [Google Scholar] [CrossRef] [PubMed]
- Turnbaugh, P.J.; Backhed, F.; Fulton, L.; Gordon, J.I. Diet-induced obesity is linked to marked but reversible alterations in the mouse distal gut microbiome. Cell Host Microbe 2008, 3, 213–223. [Google Scholar] [CrossRef] [PubMed]
- Ley, R.E.; Turnbaugh, P.J.; Klein, S.; Gordon, J.I. Microbial ecology: Human gut microbes associated with obesity. Nature 2006, 444, 1022–1023. [Google Scholar] [CrossRef] [PubMed]
- Mirnalini, K., Jr.; Zalilah, M.S.; Safiah, M.Y.; Tahir, A.; Siti Haslinda, M.D.; Siti Rohana, D.; Khairul Zarina, M.Y.; Mohd Hasyami, S.; Normah, H. Energy and Nutrient Intakes: Findings from the Malaysian Adult Nutrition Survey (MANS). Malays. J. Nutr. 2008, 14, 1–24. [Google Scholar] [PubMed]
- Raynaud-Simon, A.; Belabed, L.; Le Naour, G.; Marc, J.; Capron, F.; Cynober, L.; Darquy, S. Arginine plus proline supplementation elicits metabolic adaptation that favors wound healing in diabetic rats. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2012, 303, R1053–R1061. [Google Scholar] [CrossRef]
- Hobson, R.M.; Saunders, B.; Ball, G.; Harris, R.C.; Sale, C. Effects of beta-alanine supplementation on exercise performance: A meta-analysis. Amino Acids 2012, 43, 25–37. [Google Scholar] [CrossRef]
- Dubourg, G.; Sankar, S.A.; Rathored, J.; Lagier, J.C.; Robert, C.; Couderc, C.; Papazian, L.; Raoult, D.; Fournier, P.E. Noncontiguous finished genome sequence and description of Nocardioides massiliensis sp. nov. GD13(T). New Microbes New Infect. 2016, 10, 47–57. [Google Scholar] [CrossRef]
- Covacci, A.; Telford, J.L.; Del Giudice, G.; Parsonnet, J.; Rappuoli, R. Helicobacter pylori virulence and genetic geography. Science 1999, 284, 1328–1333. [Google Scholar] [CrossRef]
- Scholz, C.F.; Kilian, M. The natural history of cutaneous propionibacteria, and reclassification of selected species within the genus Propionibacterium to the proposed novel genera Acidipropionibacterium gen. nov., Cutibacterium gen. nov. and Pseudopropionibacterium gen. nov. Int J. Syst. Evol. Microbiol. 2016, 66, 4422–4432. [Google Scholar] [CrossRef]
- Montalban-Arques, A.; Wurm, P.; Trajanoski, S.; Schauer, S.; Kienesberger, S.; Halwachs, B.; Hogenauer, C.; Langner, C.; Gorkiewicz, G. Propionibacterium acnes overabundance and natural killer group 2 member D system activation in corpus-dominant lymphocytic gastritis. J. Pathol. 2016, 240, 425–436. [Google Scholar] [CrossRef] [PubMed]
- Madisch, A.; Andresen, V.; Enck, P.; Labenz, J.; Frieling, T.; Schemann, M. The Diagnosis and Treatment of Functional Dyspepsia. Dtsch Arztebl. Int. 2018, 115, 222–232. [Google Scholar] [CrossRef] [PubMed] [Green Version]






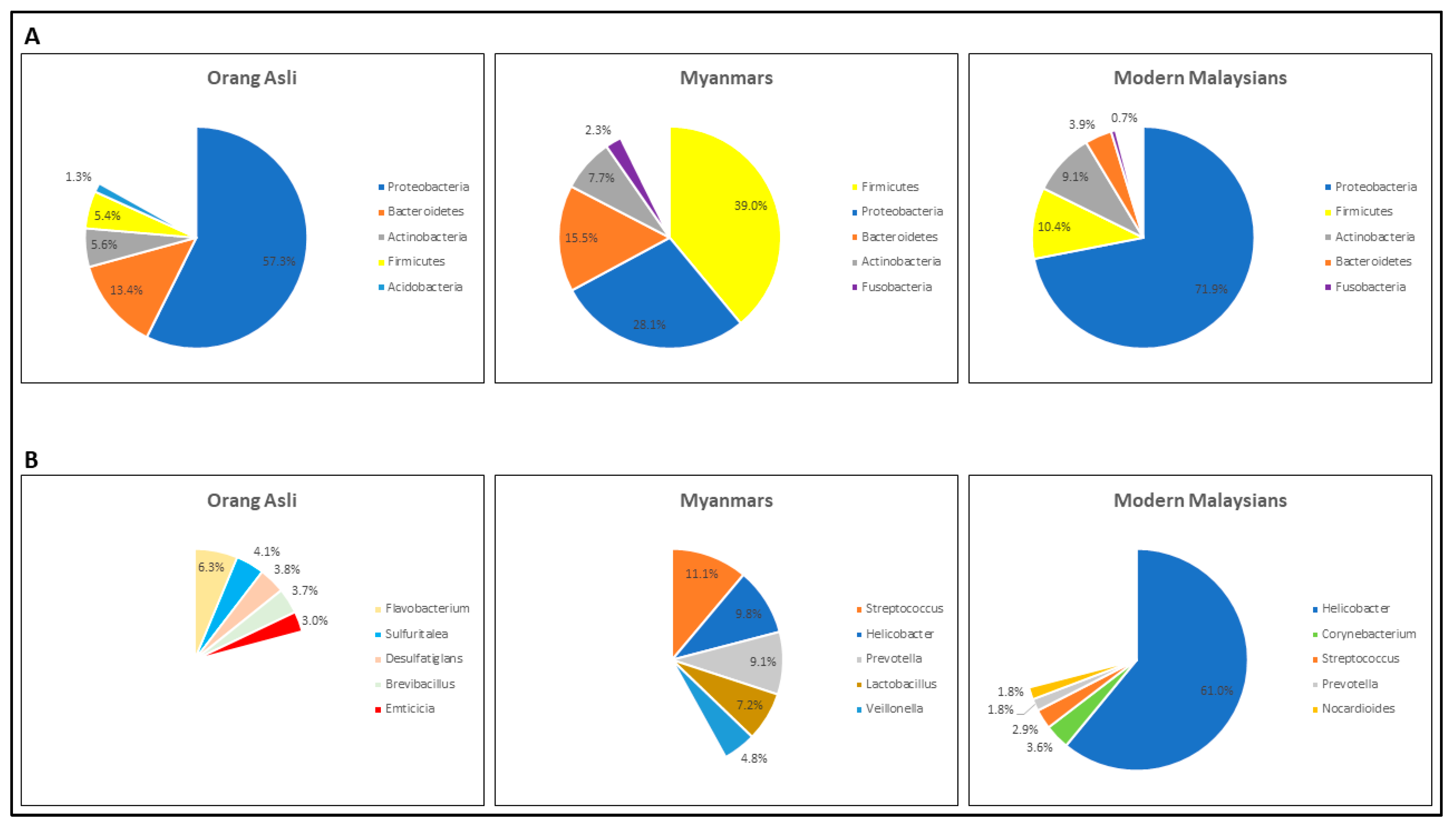{kind=link}
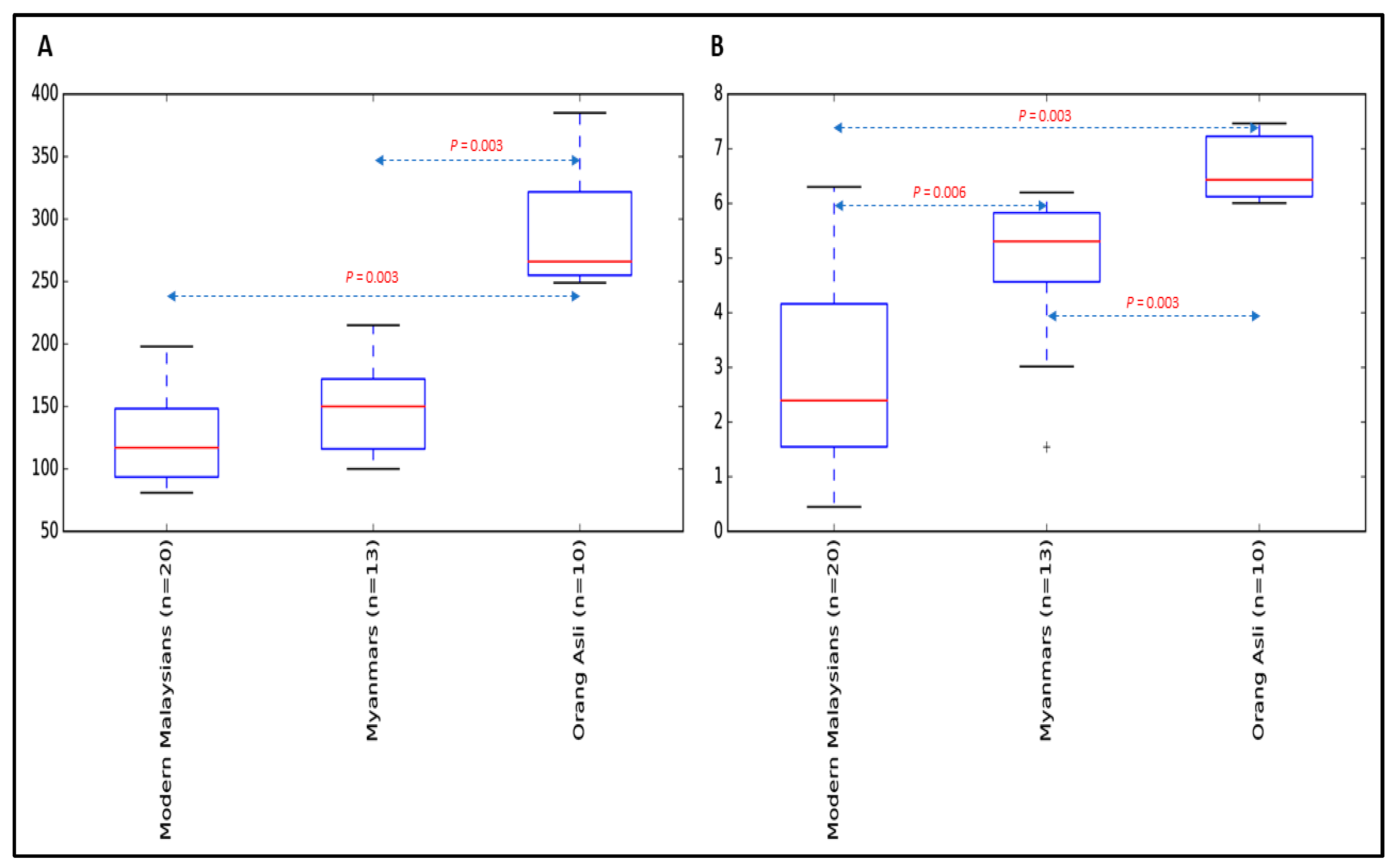{kind=link}
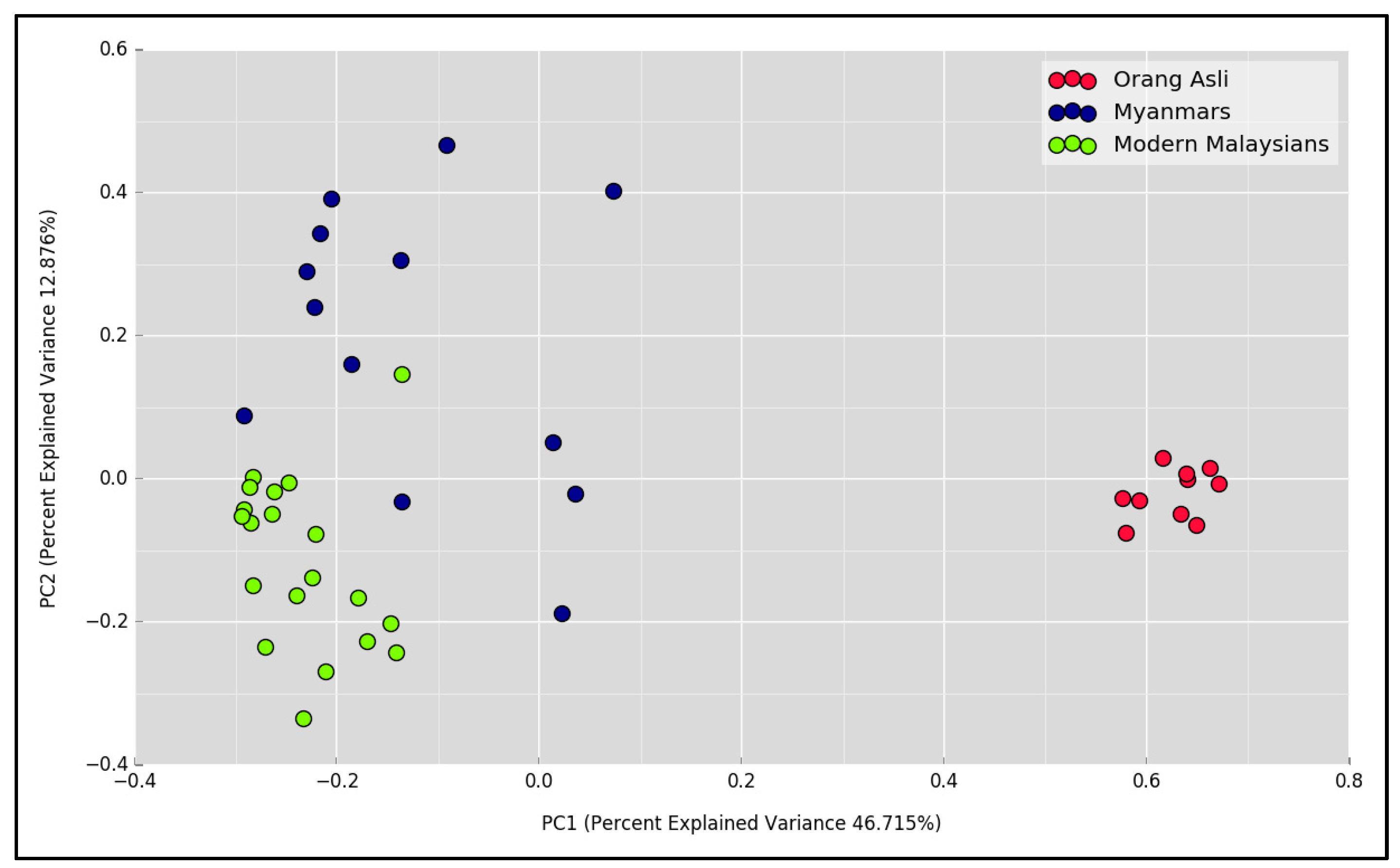{kind=link}
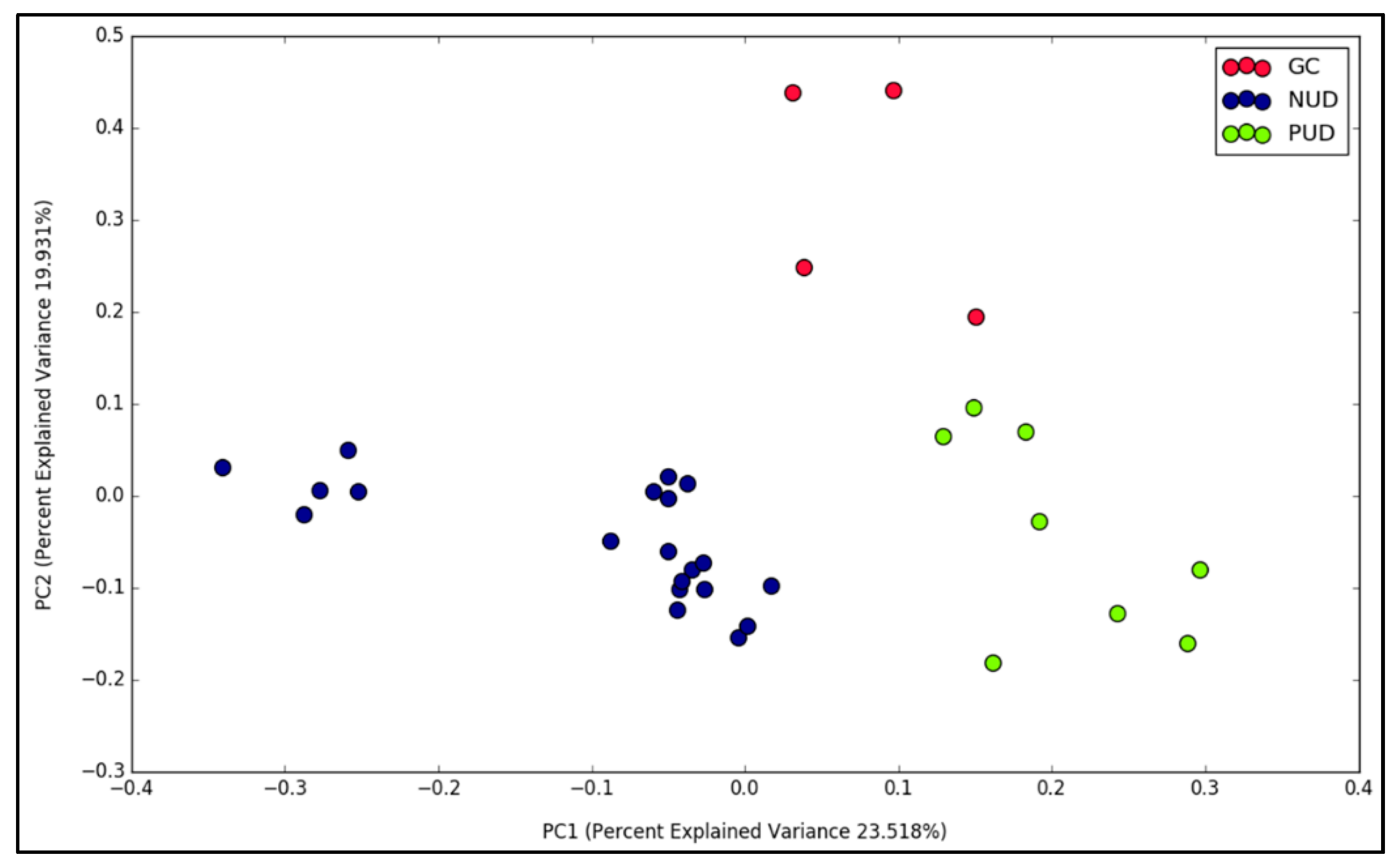{kind=link}
{kind=link}
| Origin | N | Positive | Indeterminate | Negative | Mean Age (years) | Married (%) | Female (%) |
|---|---|---|---|---|---|---|---|
| Temiar | 69 | 3 | 49 | 17 | 33 (16–80) | 85% | 67% |
| Temuan | 56 | 3 | 32 | 21 | 29 (15–56) | 69% | 51% |
| Semai | 45 | 11 | 19 | 15 | 32 (14–60) | 83% | 77% |
| Others (Mah Meri, Iban, Jah Hut and Jakun) | 7 | 1 | 6 | 0 | 39 (27–53) | 43% | 57% |
| Country | Ethnic Group | N | Mean Age (years) | NUD | PUD | GC |
|---|---|---|---|---|---|---|
| Malaysia | Orang Asli (Semai) | 3 | 33.0 | 3 | 0 | 0 |
| Orang Asli (Temiar) | 7 | 32.3 | 7 | 0 | 0 | |
| Chinese | 16 | 47.9 | 7 | 6 | 3 | |
| Indian | 10 | 34.6 | 8 | 1 | 1 | |
| Malay | 6 | 37.5 | 5 | 1 | 0 | |
| Overall | 42 | 39.6 | 30 | 8 | 4 | |
| Myanmar | Burmese | 22 | 48.3 | 5 | 11 | 6 |
| Rakhine | 5 | 47.4 | 2 | 1 | 2 | |
| Karen | 3 | 36.7 | 2 | 0 | 1 | |
| Chin | 2 | 41.5 | 2 | 0 | 0 | |
| Mon | 2 | 31.5 | 2 | 0 | 0 | |
| Others (Kachin, Paoh) | 3 | 61.7 | 0 | 2 | 1 | |
| Overall | 37 | 47.1 | 13 | 14 | 10 |
| Genus | Median Relative Abundance (%) | Adjusted p-Values | ||||
|---|---|---|---|---|---|---|
| OA (n = 10) | MYA (n = 13) | MM (n = 20) | OA vs. MYA | OA vs. MM | MYA vs. MM | |
| Aquabacterium | 0.69 | 0 | 0 | 0.005 | <0.001 | N.S |
| Asticcacaulis | 0.77 | 0 | 0 | 0.004 | <0.001 | N.S |
| Bosea | 1.24 | 0 | 0 | 0.004 | <0.001 | N.S |
| Brevibacillus | 2.09 | 0 | 0 | 0.004 | <0.001 | N.S |
| Brevundimonas | 1.86 | 0.01 | 0 | 0.004 | 0.001 | N.S |
| Caulobacter | 0.72 | 0 | 0 | 0.004 | 0.001 | N.S |
| Emticicia | 3.11 | 0 | 0 | 0.004 | <0.001 | N.S |
| Flavobacterium | 7.7 | 0 | 0 | 0.004 | <0.001 | N.S |
| Ideonella | 2.07 | 0 | 0 | 0.004 | <0.001 | N.S |
| Legionella | 0.84 | 0 | 0 | 0.004 | <0.001 | N.S |
| Massilia | 3.1 | 0 | 0.05 | 0.004 | <0.001 | N.S |
| Methyloversatilis | 0.63 | 0 | 0 | 0.004 | 0.001 | N.S |
| Phenylobacterium | 2.96 | 0 | 0 | 0.004 | <0.001 | N.S |
| Sphingorhabdus | 2.18 | 0 | 0 | 0.004 | <0.001 | N.S |
| Sulfuritalea | 3.96 | 0 | 0 | 0.004 | <0.001 | N.S |
| Tardiphaga | 0.55 | 0 | 0 | 0.004 | <0.001 | N.S |
| Variovorax | 1.49 | 0 | 0 | 0.004 | <0.001 | N.S |
| Helicobacter | 0.01 | 0.49 | 71.69 | N.S | 0.001 | 0.01 |
| Nocardioides | 0.88 | 0 | 0.42 | 0.005 | N.S | 0.001 |
| Agrobacterium | 0.62 | 0 | 0 | 0.005 | N.S | N.S |
| Streptococcus | 0.36 | 11.06 | 1.89 | 0.007 | N.S | N.S |
| Anaerococcus | 0.01 | 0.01 | 0.59 | N.S | 0.004 | N.S |
| Corynebacterium | 0.1 | 0.41 | 1.6 | N.S | 0.001 | N.S |
| Dehalogenimonas | 0.29 | 0 | 0 | N.S | 0.01 | N.S |
| Desulfatiglans | 1.85 | 0 | 0 | N.S | <0.001 | N.S |
| Paludibaculum | 0.71 | 0 | 0 | N.S | 0.006 | N.S |
| Rothia | 0 | 0.44 | 0.33 | N.S | 0.006 | N.S |
| Lactobacillus | 0 | 0.59 | 0 | N.S | N.S | 0.004 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chua, E.G.; Loke, M.F.; Gunaletchumy, S.P.; Gan, H.M.; Thevakumar, K.; Tay, C.Y.; Young, S.; Aye, T.T.; Maw, W.W.; Aye, M.M.; et al. The Influence of Modernization and Disease on the Gastric Microbiome of Orang Asli, Myanmars and Modern Malaysians. Microorganisms 2019, 7, 174. https://doi.org/10.3390/microorganisms7060174
Chua EG, Loke MF, Gunaletchumy SP, Gan HM, Thevakumar K, Tay CY, Young S, Aye TT, Maw WW, Aye MM, et al. The Influence of Modernization and Disease on the Gastric Microbiome of Orang Asli, Myanmars and Modern Malaysians. Microorganisms. 2019; 7(6):174. https://doi.org/10.3390/microorganisms7060174
Chicago/Turabian StyleChua, Eng Guan, Mun Fai Loke, Selva Perumal Gunaletchumy, Han Ming Gan, Kavitha Thevakumar, Chin Yen Tay, Sylvia Young, Than Than Aye, Win Win Maw, Mya Mya Aye, and et al. 2019. "The Influence of Modernization and Disease on the Gastric Microbiome of Orang Asli, Myanmars and Modern Malaysians" Microorganisms 7, no. 6: 174. https://doi.org/10.3390/microorganisms7060174
APA StyleChua, E. G., Loke, M. F., Gunaletchumy, S. P., Gan, H. M., Thevakumar, K., Tay, C. Y., Young, S., Aye, T. T., Maw, W. W., Aye, M. M., Leow, A. H.-R., Azmi, A. N., Kalimuthu, S. G., Dahlan, H. M. A. B. D., Goh, K. L., & Vadivelu, J. (2019). The Influence of Modernization and Disease on the Gastric Microbiome of Orang Asli, Myanmars and Modern Malaysians. Microorganisms, 7(6), 174. https://doi.org/10.3390/microorganisms7060174