Organs, Cultivars, Soil, and Fruit Properties Affect Structure of Endophytic Mycobiota of Pinggu Peach Trees


Abstract
:1. Introduction
2. Materials and Methods
2.1. Collection and Processing of Samples
2.2. DNA Extraction, Amplification of ITS rDNA Region and Sequencing
2.3. Data Processing and Analysis
3. Results
3.1. Overview of the Sequencing Data and Alpha Diversit of Fungal Communities
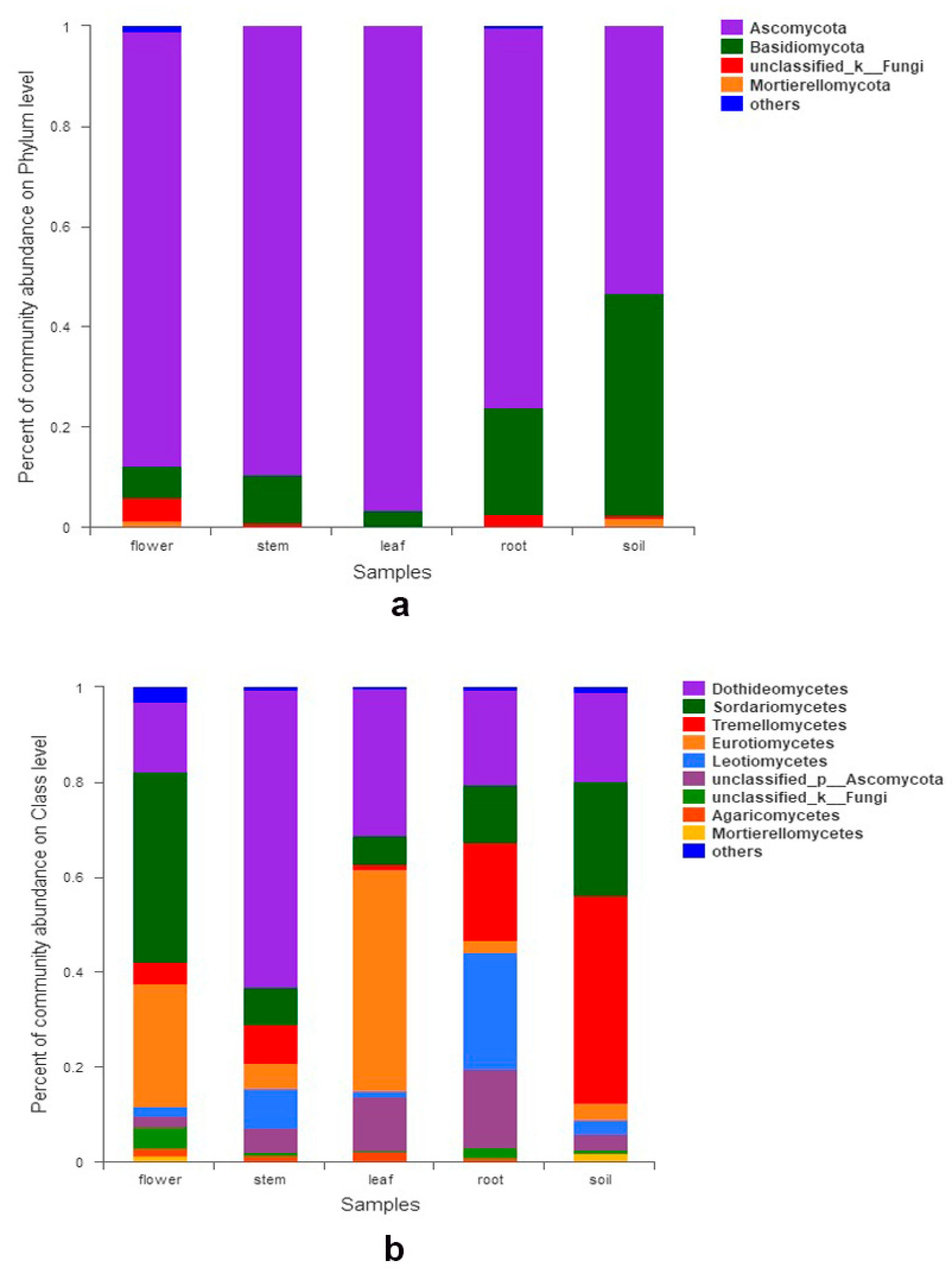
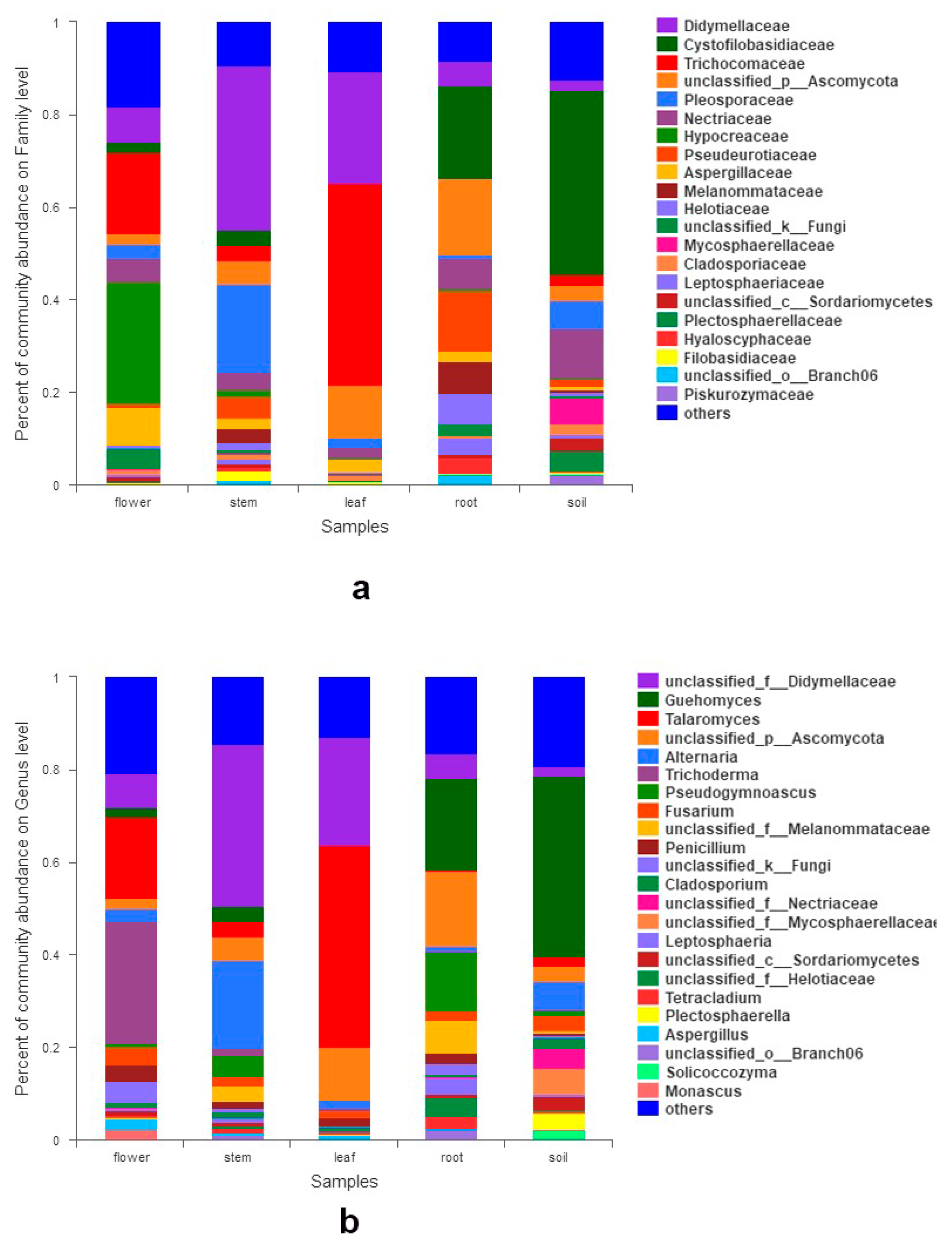
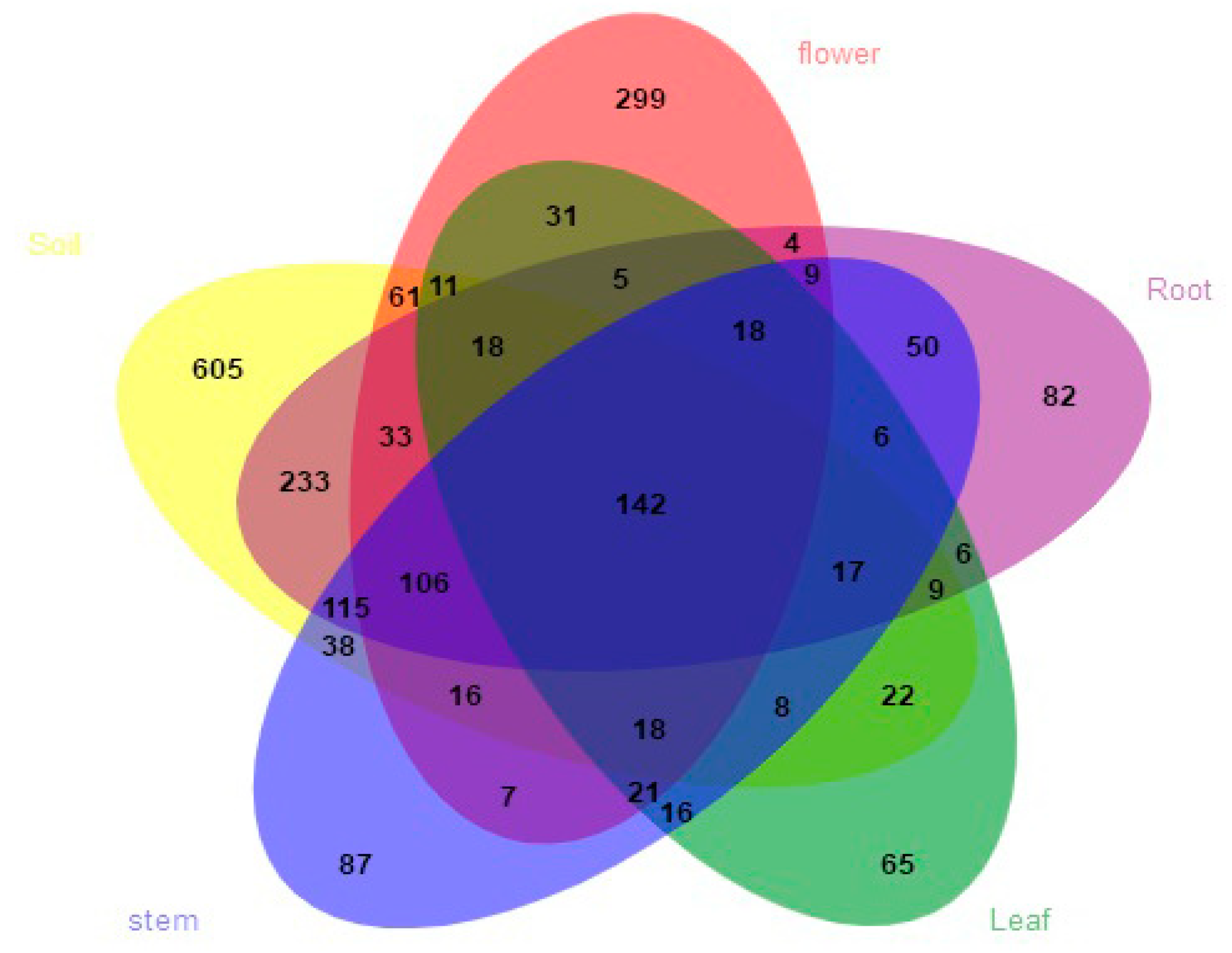
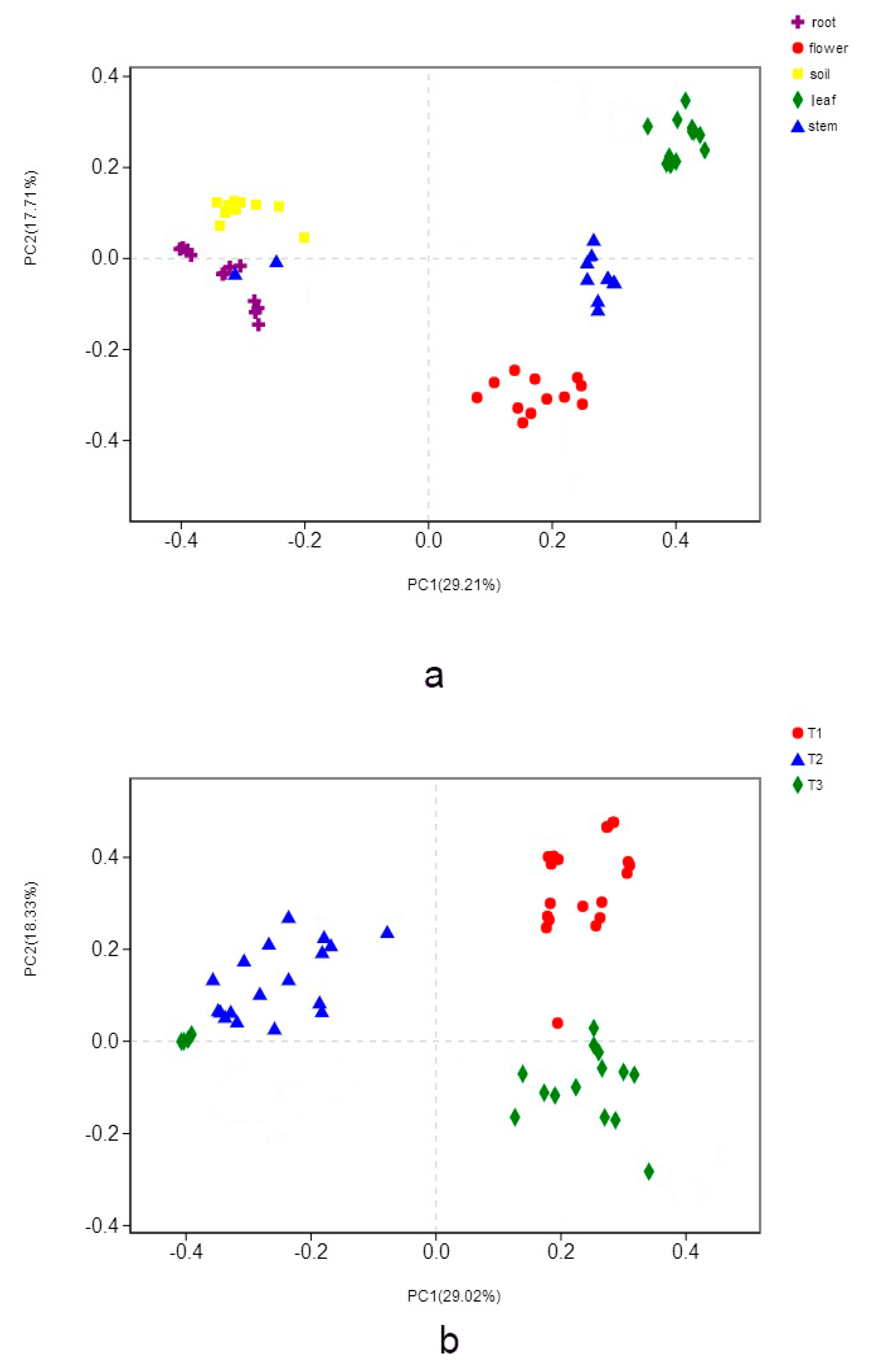
3.2. Fungal Composition and Relative Abundance among Different Organs and Soils
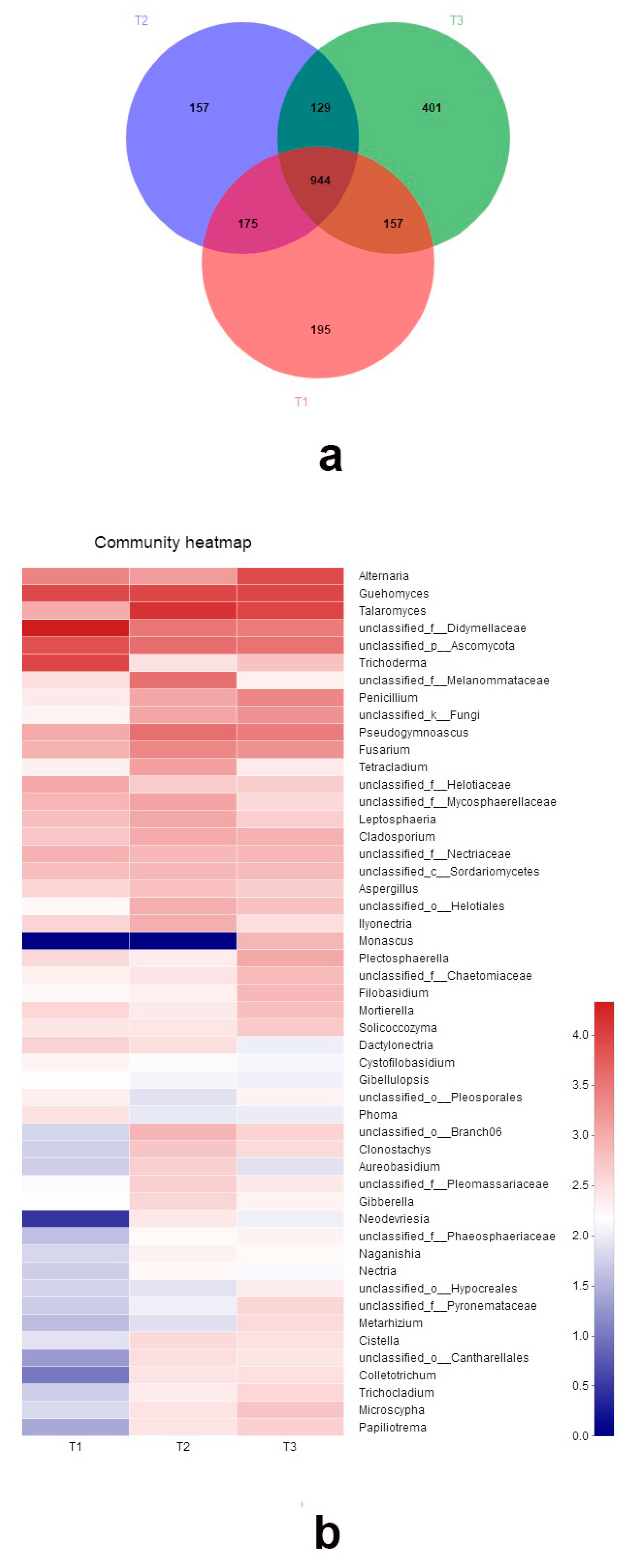
3.3. Fungal Community among Three Peach Cultivars
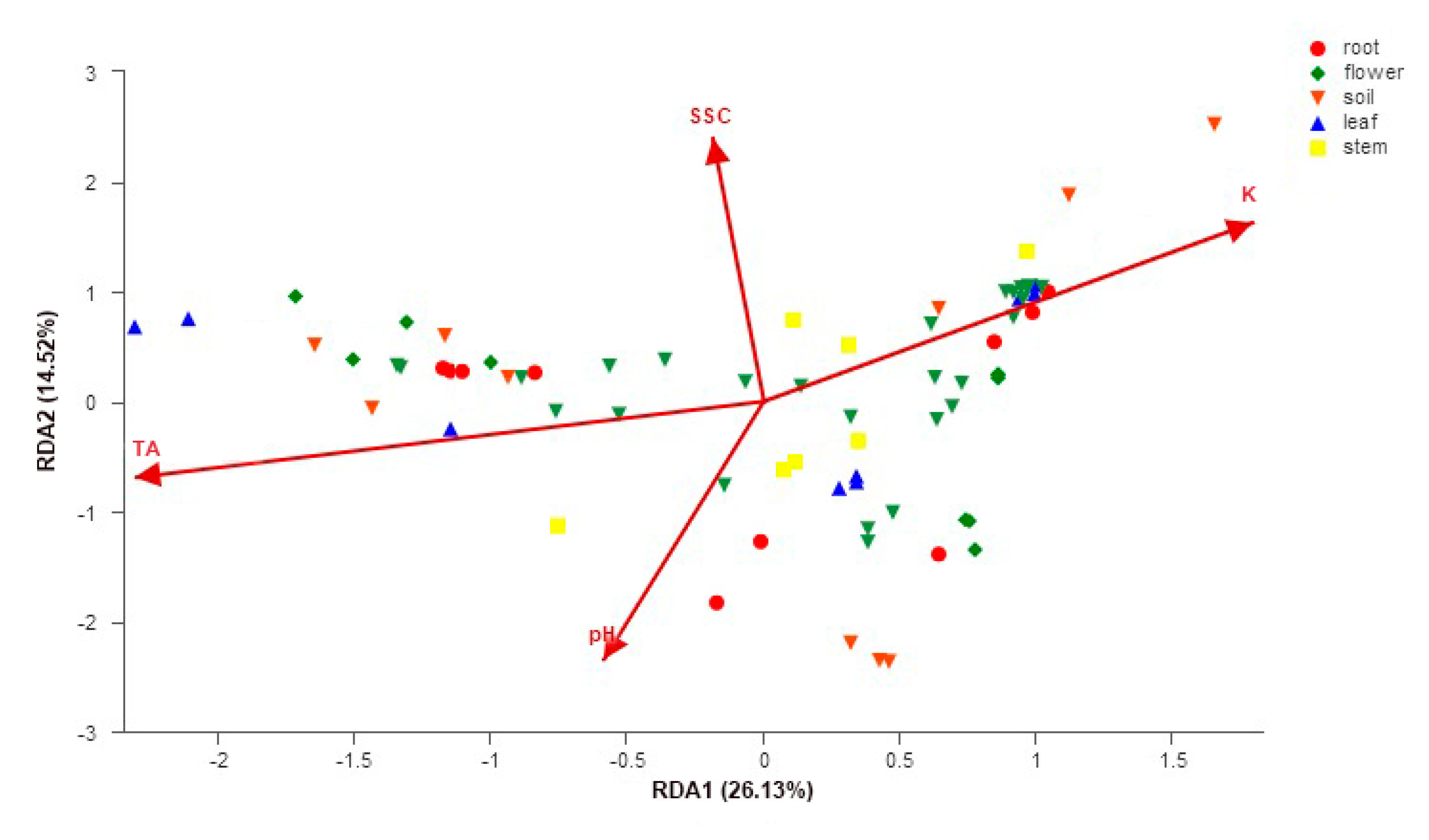
3.4. Impacts of Organs, Peach Cultivars, Soil Properties, and Peach Fruit Properties on Fungal Community Structure
3.5. Culture-Dependent Isolation of Endophytic Fungi of the Pinggu Peach Trees
4. Discussion
4.1. Fungal Communities among Organs
4.2. Fungal Communities of Cultivars
4.3. Correlation between Fungal Communities and Soil and Fruit Properties
4.4. Isolated Endophytic Strains and Future Studies
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Azevedo, J.L.; Maccheroni, W.; Pereira, J.A.; Araujo, W.L. Endophytic microorganisms: A review on insect control and recent advances on tropical plants. Electron. J. Biotechnol. 2000, 3, 15–16. [Google Scholar] [CrossRef]
- Petrini, O.; Stone, J.; Carroll, F.E. Endophytic fungi in evergreen shrubs in western Oregon: A preliminary study. Can. J. Bot. 1982, 60, 789–796. [Google Scholar] [CrossRef]
- Jia, M.; Chen, L.; Xin, H.L.; Zheng, C.J.; Rahman, K.; Han, T.; Qin, L.P. A friendly relationship between endophytic fungi and medicinal plants: A systematic review. Front. Microbiol. 2016, 7, 906. [Google Scholar] [CrossRef]
- Muller, D.B.; Vogel, C.; Bai, Y.; Vorholt, J.A. The plant microbiota: Systems-level insights and perspectives. Annu. Rev. Genet. 2016, 50, 211–234. [Google Scholar] [CrossRef]
- Photita, W.; Lumyong, S.; Lumyong, P.; Hyde, K.D. Endophytic fungi of wild banana (Musa acuminata) at doi Suthep Pui National Park, Thailand. Mycol. Res. 2001, 105, 1508–1513. [Google Scholar] [CrossRef]
- Porras-Alfaro, A.; Bayman, P. Hidden fungi, emergent properties: Endophytes and microbiomes. Annu. Rev. Phytopathol. 2011, 49, 291–315. [Google Scholar] [CrossRef]
- Xu, J.; Ebada, S.S.; Proksch, P. Pestalotiopsis a highly creative genus: Chemistry and bioactivity of secondary metabolites. Fungal Divers. 2010, 44, 15–31. [Google Scholar] [CrossRef]
- Chen, L.; Zhang, Q.Y.; Jia, M.; Ming, Q.L.; Yue, W.; Rahman, K.; Han, T. Endophytic fungi with antitumor activities: Their occurrence and anticancer compounds. Crit. Rev. Microbiol. 2016, 42, 454–473. [Google Scholar] [CrossRef]
- Liao, H.L.; Bonito, G.; Rojas, J.A.; Hameed, K.; Wu, S.; Schadt, C.W. Fungal endophytes of Populus trichocarpa alter host phenotype, gene expression and rhizobiome composition. Mol. Plant-Microbe Interact. 2019, 32, 853. [Google Scholar] [CrossRef]
- Sun, X.; Guo, L.D. Endophytic fungal diversity: Review of traditional and molecular techniques. Mycology 2012, 3, 65–76. [Google Scholar]
- Pinto, C.; Pinho, D.; Sousa, S.; Pinheiro, M.; Egas, C.; Gomes, A.C. Unravelling the diversity of grapevine microbiome. PLoS ONE 2014, 9, e85622. [Google Scholar] [CrossRef]
- Berg, G.; Rybakova, D.; Grube, M.; Koberl, M. The plant microbiome explored: Implications for experimental botany. J. Exp. Bot. 2016, 67, 995–1002. [Google Scholar] [CrossRef]
- Sun, X.; Guo, L.D.; Hyde, K.D. Community composition of endophytic fungi in Acer truncatum and their role in decomposition. Fungal Divers. 2011, 47, 85–95. [Google Scholar] [CrossRef]
- Li, F.Z. Pinggu Da Tao; China Agriculture Press: Beijing, China, 2017; pp. 1–431. (In Chinese) [Google Scholar]
- Wang, Y.H.; Yang, J.G.; Zhang, J.L.; Zhang, B.C.; Xu, S.M. Green control techniques test of Beijing Pinggu large peach disease insect damage. China Plant Prot. 2012, 32, 20–23. (In Chinese) [Google Scholar]
- Larrán, S.; Mónaco, C. Status and progress of research in endophytes from agricultural crops in Argentina. In Management of Fungal Plant Pathogens; Arya, A., Perello, A.E., Eds.; CAB International: Cambridge, Wallingford, UK, 2010; Chapter 12; p. 149. [Google Scholar]
- Russo, M.L.; Pelizza, S.A.; Cabello, M.N.; Stenglein, S.A.; Vianna, M.F.; Scorsetti, A.C. Endophytic fungi from selected varieties of soybean (Glycine max L. Merr.) and corn (Zea mays L.) grown in an agricultural area of Argentina. Rev. Argent. De. Microbiol. 2016, 48, 154–160. [Google Scholar] [CrossRef]
- Bezerra, J.D.; Nascimento, C.C.; Barbosa, R.D.N.; da Silva, D.C.; Svedese, V.M.; Silva–Nogueira, E.B.; Souza–Motta, C.M. Endophytic fungi from medicinal plant Bauhinia forficata: Diversity and biotechnological potential. Braz. J. Microbiol. 2015, 46, 49–57. [Google Scholar] [CrossRef]
- Giordano, L.; Gonthier, P.; Varese, G.C.; Miserere, L.; Nicolotti, G. Mycobiota inhabiting sapwood of healthy and declining Scots pine (Pinus sylvestris L.) trees in the Alps. Fungal Divers. 2009, 38, 69–83. [Google Scholar]
- Kovalchuk, A.; Mukrimin, M.; Zeng, Z.; Raffaello, T.; Liu, M.X.; Kasanen, R.; Sun, H.; Asiegbu, F.O. Mycobiome analysis of asymptomatic and symptomatic Norway spruce trees naturally infected by the conifer pathogens Heterobasidion spp. Environ. Microbiol. Rep. 2018, 10, 532–541. [Google Scholar] [CrossRef]
- Hortova, B.; Novotny, D. Endophytic fungi in branches of sour cherry trees: A preliminary study. Czech Mycol. 2011, 63, 77–82. [Google Scholar] [CrossRef]
- Novotny, D. Studium endofytickych hub zemedelsky vyznamnch rostlin. In Sbornik prispevku z workshopu Micromyco; Novakova, A., Ed.; Ústav půdní biologie: Ceske Budejovice, Czech Republic, 2007; pp. 97–101. [Google Scholar]
- Johnston, P.R. Endophytes of apple and kiwifruit. Proc. N. Z. Plant Prot. Conf. 1994. [Google Scholar] [CrossRef]
- Camatti–Sartori, V.; da Silva–Ribeiro, R.T.; Valdebenito-Sanhueza, R.M.; Pagnocca, F.C.; Echeverrigaray, S.; Azevedo, J.L. Endophytic yeasts and filamentous fungi associated with southern Brazilian apple (Malus domestica) orchards subjected to conventional, integrated or organic cultivation. J. Basic Microb. 2005, 45, 397–402. [Google Scholar] [CrossRef]
- Photita, W.; Lumyong, S.; Lumyong, P.; McKenzie, E.H.C.; Hyde, K.D. Are some endophytes of Musa acuminata latent pathogens? Fungal Divers. 2004, 16, 131–140. [Google Scholar]
- Janisiewicz, W.J.; Jurick, W.M.; Peter, K.A.; Kurtzman, C.P.; Buyer, J.S. Yeasts associated with plums and their potential for controlling brown rot after harvest. Yeast 2014, 31, 207–218. [Google Scholar] [CrossRef]
- Melgarejo, P.; Carrillo, R.; Sagasta, E.M. Mycoflora of peach twigs and flowers and its possible significance in biological control of Monilinia laxa. Trans. Br. Mycol. Soc. 1985, 85, 313–317. [Google Scholar] [CrossRef]
- Vepštaitė–Monstavičė, I.; Lukša, J.; Stanevičienė, R.; Strazdaitė–Žielienė, Ž.; Yurchenko, V.; Serva, S.; Servienė, E. Distribution of apple and blackcurrant microbiota in Lithuania and the Czech Republic. Microbiol. Res. 2018, 206, 1–8. [Google Scholar] [CrossRef]
- Liu, J.; Abdelfattah, A.; Norelli, J.; Burchard, E.; Schena, L.; Droby, S.; Wisniewski, M. Apple endophytic microbiota of different rootstock/scion combinations suggests a genotype–specific influence. Microbiome 2018, 6, 18. [Google Scholar] [CrossRef]
- Ren, F.; Dong, W.; Sun, H.; Yan, D.H. Endophytic Mycobiota of Jingbai Pear Trees in North China. Forests 2019, 10, 260. [Google Scholar] [CrossRef]
- Black, C.A.; Evans, D.D.; Dinauer, R.C. Methods of Soil Analysis; American Society of Agronomy: Madison, WI, USA, 1965; pp. 1–1178. [Google Scholar]
- Wang, L.R.; Zhu, G.R.; Fang, W.C. Descriptors and Data Standard for Peach; China Agriculture Press: Beijing, China, 2005; pp. 1–98. (In Chinese) [Google Scholar]
- Chang, S.; Puryear, J.; Cairney, J. A simple and efficient method for isolating RNA from pine trees. Plant. Mol. Biol. Rep. 1993, 11, 113–116. [Google Scholar] [CrossRef]
- Lanzén, A.; Epelde, L.; Blanco, F.; Martín, I.; Artetxe, U.; Garbisuet, C. Multi–targeted metagenetic analysis of the influence of climate and environmental parameters on soil microbial communities along an elevational gradient. Sci. Rep. 2016, 6, e28257. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogeneics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic: San Diego, CA, USA, 1990; pp. 315–322. [Google Scholar]
- Magoč, T.; Salzberg, S.L. FLASH: Fast length adjustment of short reads to improve genome assemblies. Bioinformatics 2011, 27, 2957–2963. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef]
- Schloss, P.D.; Gevers, D.; Westcott, S.L. Reducing the effects of PCR amplification and sequencing artifacts on 16S rRNA–based studies. PLoS ONE 2011, 6, e27310. [Google Scholar] [CrossRef]
- UNITE Community. UNITE mothur release. Version 01.12.2017. UNITE Community 2017. [Google Scholar] [CrossRef]
- Mothur Calculators. Available online: https://www.mothur.org/wiki/Calculators (accessed on 6 June 2019).
- R Core Team. R: A language and environment for statistical computing. R foundation for Statistical Computing: Vienna, Austria; Available online: http://www.R-project.org. (accessed on 15 July 2019).
- LEfSe. Available online: http://huttenhower.sph.harvard.edu/galaxy/root?tool_id=lefse_upload (accessed on 5 June 2019).
- NCBI Blast. Available online: https://blast.ncbi.nlm.nih.gov/Blast.cgi (accessed on 8 June 2019).
- Wang, H.Y.; Guo, S.Y.; Huang, M.R.; Thorsten, L.H.; Wei, J.C. Ascomycota has a faster evolutionary rate and higher species diversity than Basidiomycota. Sci. China Life Sci. 2010, 53, 1163–1169. [Google Scholar] [CrossRef]
- Botella, L.; Diez, J.J. Phylogenic diversity of fungal endophytes in Spanish stands of Pinus halepensis. Fungal Divers. 2011, 47, 9–18. [Google Scholar] [CrossRef]
- Pawłowska, J.; Wilk, M.; Śliwińska–Wyrzychowska, A.; Mętrak, M.; Wrzosek, M. The diversity of endophytic fungi in the above–ground tissue of two Lycopodium species in Poland. Symbiosis 2014, 63, 87–97. [Google Scholar] [CrossRef]
- Nelsen, M.P.; Lücking, R.; Grube, M.; Mbatchou, J.S.; Muggia, L.; Plata, E.R.; Lumbsch, H.T. Unravelling the phylogenetic relationships of lichenised fungi in Dothideomyceta. Stud. Mycol. 2009, 64, 135–144. [Google Scholar] [CrossRef]
- Higgins, K.L.; Arnold, A.E.; Miadlikowska, J.; Sarvate, S.D.; Lutzoni, F. Phylogenetic relationships, host affinity, and geographic structure of boreal and arctic endophytes from three major plant lineages. Mol. Phylogenet. Evol. 2007, 42, 543–555. [Google Scholar] [CrossRef]
- Maharachchikumbura, S.S.; Hyde, K.D.; Jones, E.G.; McKenzie, E.H.; Huang, S.K.; Abdel–Wahab, M.A.; Hongsanan, S. Towards a natural classification and backbone tree for Sordariomycetes. Fungal Divers. 2015, 72, 199–301. [Google Scholar] [CrossRef]
- Geiser, D.M.; LoBuglio, K.F.; Gueidan, C. Pezizomycotina: Eurotiomycetes Systematics and Evolution; Springer: Berlin/Heidelberg, Germany, 2015; pp. 121–141. [Google Scholar]
- Earl, A.J.; Turner, G.; Croft, J.H. High frequency transfer of species specific mitochondrial DNA sequences between members of the Aspergillaceae. Curr. Genet. 1981, 3, 221–228. [Google Scholar] [CrossRef]
- Kwon, Y.J.; Sohn, M.J.; Zheng, C.J.; Kim, W.G. Fumimycin: A peptide deformylase inhibitor with an unusual skeleton produced by Aspergillus fumisynnematus. Org. Lett. 2007, 9, 2449–2451. [Google Scholar] [CrossRef]
- Houbraken, J.; Samson, R.A. Phylogeny of Penicillium and the segregation of Trichocomaceae into three families. Stud. Mycol. 2011, 70, 1–51. [Google Scholar] [CrossRef]
- Yilmaz, N.; Visagie, C.M.; Frisvad, J.C.; Houbraken, J.; Jacobs, K.; Samson, R.A. Taxonomic re–evaluation of species in Talaromyces section Islandici, using a polyphasic approach. Persoonia 2016, 36, 37. [Google Scholar] [CrossRef]
- Nicoletti, R.; Salvatore, M.M.; Andolfi, A. Secondary Metabolites of Mangrove–Associated Strains of Talaromyces. Marine Drugs 2018, 16, 12. [Google Scholar] [CrossRef]
- Harman, G.E.; Howell, C.R.; Viterbo, A. Trichoderma species—Opportunistic, avirulent plant symbionts. Nat. Rev. Microbiol. 2004, 2, 43. [Google Scholar] [CrossRef]
- Woudenberg, J.H.C.; Groenewald, J.Z.; Binder, M.; Crous, P.W. Alternaria redefined. Stud. Mycol. 2013, 75, 171–212. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, H.X.; Chen, Y.C.; Sun, Z.H.; Li, H.H.; Li, S.N.; Yan, M.L.; Zhang, W.M. Two new metabolites from the endophytic fungus Alternaria sp. A744 derived from Morinda officinalis. Molecules 2017, 22, 765. [Google Scholar] [CrossRef]
- Xiang, L.; Gong, S.; Yang, L.; Hao, J.; Xue, M.; Zeng, F.; Yu, D. Biocontrol potential of endophytic fungi in medicinal plants from Wuhan Botanical Garden in China. Biol. Control 2016, 94, 47–55. [Google Scholar] [CrossRef]
- Summerell, B.A. Resolving Fusarium: Current Status of the Genus. Annu. Rev. Phytopathol. 2019, 57. [Google Scholar] [CrossRef]
- Bensch, K.; Braun, U.; Groenewald, J.Z. The genus cladosporium. Stud. Mycol. 2012, 72, 1–401. [Google Scholar] [CrossRef]
- Aleklett, K.; Hart, M.; Shade, A. The microbial ecology of flowers: An emerging frontier in phyllosphere research. Botany 2014, 92, 253–266. [Google Scholar] [CrossRef]
- Shade, A.; McManus, P.S.; Handelsman, J. Unexpected diversity during community succession in the apple flower microbiome. MBio 2013, 4, e00602-12. [Google Scholar] [CrossRef]
- Varanda, C.M.R.; Materatski, P.; Landum, M. Fungal Communities Associated with Peacock and Cercospora Leaf Spots in Olive. Plants 2019, 8, 169. [Google Scholar] [CrossRef]
- Li, Q.; Guo, R.; Li, Y. Insight into the bacterial endophytic communities of peach cultivars related to crown gall disease resistance. Appl. Environ. Microbiol. 2019, 85, e02931-18. [Google Scholar] [CrossRef]
- Beule, L.; Chen, K.H.; Hsu, C.M. Soil bacterial and fungal communities of six bahiagrass cultivars. Peer J 2019, 7, e7014. [Google Scholar] [CrossRef]
- Kwak, M.J.; Kong, H.G.; Choi, K.; Kwon, S.K.; Song, J.Y.; Lee, J.; An Lee, P.; Choi, S.Y.; Seo, M.; Lee, H.J.; et al. Rhizosphere microbiome structure alters to enable wilt resistance in tomato. Nat. Biotechnol. 2018, 36, 1100. [Google Scholar] [CrossRef]
- Neumann, G.; Bott, S.; Ohler, M.; Mock, H.P.; Lippmann, R.; Grosch, R.; Smalla, K. Root exudation and root development of lettuce (Lactuca sativa L. cv. Tizian) as affected by different soils. Front. Microbiol. 2014, 5, 2. [Google Scholar] [CrossRef]
- Cavaco, T.; Longuinho, C.; Quintas, C. Chemical and microbial changes during the natural fermentation of strawberry tree (Arbutus unedo L.) fruits. J. Food Biochem. 2007, 31, 715–725. [Google Scholar] [CrossRef]
- Aneja, K.R.; Dhiman, R.; Aggarwal, N.K.; Kumar, V.; Kaur, M. Microbes associated with freshly prepared juices of citrus and carrots. Int. J. Food Sci. 2014, e408085. [Google Scholar] [CrossRef]
- Cissé, O.H.; Almeida, J.M.; Fonseca, Á. Genome sequencing of the plant pathogen Taphrina deformans, the causal agent of peach leaf curl. MBio 2013, 4, e00055-13. [Google Scholar] [CrossRef]
- Rutto, K.L.; Mizutani, F. Peach seedling growth in replant and non-replant soils after inoculation with arbuscular mycorrhizal fungi. Soil Biol. Biochem. 2006, 38, 2536–2542. [Google Scholar] [CrossRef]
- Zhou, T.; Northover, J.; Schneider, K.E. Biological control of postharvest diseases of peach with phyllosphere isolates of Pseudomonas syringae. Can. J. Plant Pathol. 1999, 21, 375–381. [Google Scholar] [CrossRef]
- Collado, J.; Platas, G.; Paulus, B.; Bills, G.F. High-throughput culturing of fungi from plant litter by a dilution-to-extinction technique. FEMS Microbiol. Ecol. 2007, 60, 521–533. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Zhu, A.; Tan, H.; Cao, L.; Zhang, R. Engineering banana endosphere microbiome to improve Fusarium wilt resistance in banana. Microbiome 2019, 7, 74. [Google Scholar] [CrossRef]
- Duran, P.; Thiergart, T.; Garrido-Oter, R.; Agler, M.; Kemen, E.; Schulze-Lefert, P. Microbial Interkingdom Interactions in Roots Promote Arabidopsis Survival. Cell 2018, 75, 973–983. [Google Scholar] [CrossRef]
- Bai, Y.; Muller, D.B.; Srinivas, G.; Garrido-Oter, R.; Potthoff, E.; Rott, M.; Dombrowski, N.; Munch, P.C.; Spaepen, S.; Remus-Emsermann, M.; et al. Functional overlap of the Arabidopsis leaf and root microbiota. Nature 2015, 528, 364–369. [Google Scholar] [CrossRef]
- Vannier, N.; Agler, M.; Hacquard, S. Microbiota-mediated disease resistance in plants. PLoS Pathog. 2019, 15, e1007740. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Samples | Sobs (Richness) | Lnvsimpson (Diversity) | Simpsoneven (Evenness) |
|---|---|---|---|
| leaf | 108.25 ± 1.84 | 4.08 ± 0.11 | 0.041 ± 0.001 |
| flower | 207.83 ± 16.16 | 7.86 ± 1.27 | 0.082 ± 0.006 |
| stem | 176.92 ± 4.81 | 7.81 ± 0.59 | 0.041 ± 0.002 |
| root | 310.67 ± 2.84 | 10.30 ± 0.15 | 0.034 ± 0.001 |
| soil | 627.41 ± 13.94 | 8.46 ± 0.31 | 0.014 ± 0.002 |
| Sample Number | Identification | NCBI Number | Percent of Identity |
|---|---|---|---|
| DH2 (5) | Penicillium funiculosum isolate B12 | JN676119 | 99% |
| DGE1 (4) | Trichoderma hamatum strain 347 | KX357867 | 99% |
| DGE3 (6) | Fusarium solani isolate C10-4 | KT876641 | 99% |
| HY1(2) | Meyerozyma guilliermondi | KP764945 | 100% |
| DY4 (4) | Talaromyces stollii | JX965246 | 100% |
| HY4 (4) | Penicillium oxalicum strain QHBC11 | KC880081 | 100% |
| DY2 (3) | Fusarium fujikuroi | NR_111889 | 100% |
| HZ6 (3) | Alternaria alternata | NR_131263 | 99% |
| DY6 (3) | Talaromyces stollii | NR_111781 | 98% |
| DH5 (3) | Penicillium caperatum CBS 443.7 | NR_138333 | 99% |
| HY3 (2) | Chaetomium globosum | NR_144833 | 99% |
| HZ1(2) | Trichoderma sp. isolate yi0319 | MK326900 | 97% |
| HZ4 (3) | Alternaria solani isolate OTA52 | JF491196 | 97% |
| HZ5 (4) | Aspergillus niger | NR_111348 | 99% |
| DY5 (2) | Cladosporium cucumerinum | NR_119841 | 99% |
| HY2 (3) | Alternaria sp. isolate JS8-5 | MF033857 | 99% |
| TY6 (3) | Arthrinium sp. GU071007 | AB471012 | 99% |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ren, F.; Dong, W.; Yan, D.-H. Organs, Cultivars, Soil, and Fruit Properties Affect Structure of Endophytic Mycobiota of Pinggu Peach Trees. Microorganisms 2019, 7, 322. https://doi.org/10.3390/microorganisms7090322
Ren F, Dong W, Yan D-H. Organs, Cultivars, Soil, and Fruit Properties Affect Structure of Endophytic Mycobiota of Pinggu Peach Trees. Microorganisms. 2019; 7(9):322. https://doi.org/10.3390/microorganisms7090322
Chicago/Turabian StyleRen, Fei, Wei Dong, and Dong-Hui Yan. 2019. "Organs, Cultivars, Soil, and Fruit Properties Affect Structure of Endophytic Mycobiota of Pinggu Peach Trees" Microorganisms 7, no. 9: 322. https://doi.org/10.3390/microorganisms7090322
APA StyleRen, F., Dong, W., & Yan, D. -H. (2019). Organs, Cultivars, Soil, and Fruit Properties Affect Structure of Endophytic Mycobiota of Pinggu Peach Trees. Microorganisms, 7(9), 322. https://doi.org/10.3390/microorganisms7090322