Development of Salmonellosis as Affected by Bioactive Food Compounds


Abstract
:1. Introduction
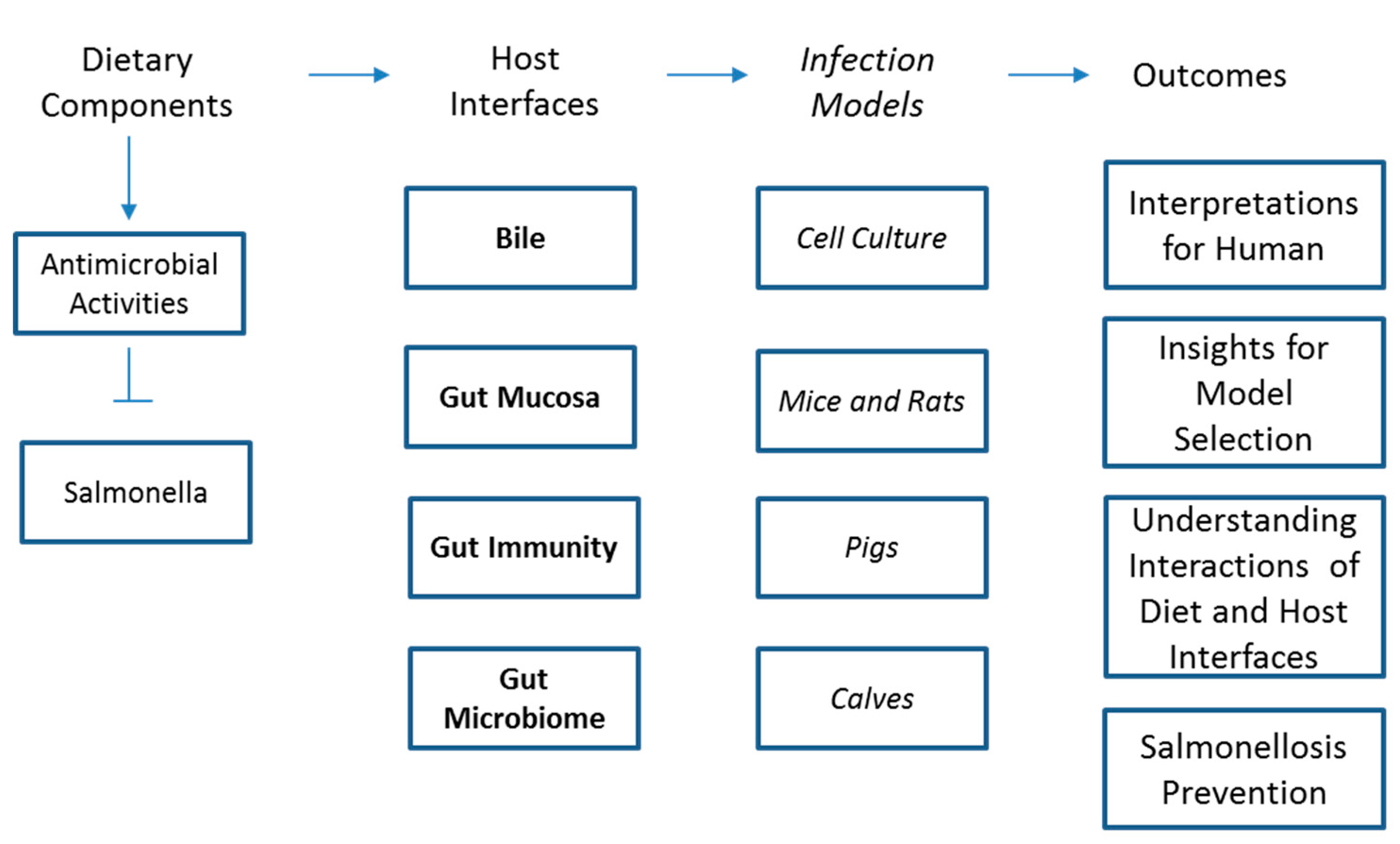
2. Current Status of Knowledge
2.1. Effect of Dietary Components against Salmonella: In-Vitro Models
2.2. Summary of Effect of Dietary Components on Salmonella Infection in Rodent Models
2.3. Summary of Effect of Dietary Components on Salmonella Infection in Pig Models
2.4. Summary of Effect of Dietary Components on Salmonella Infection in Calf Models
2.5. Summary of Dietary Interventions for Salmonella Infection in Humans
3. Potential Mechanisms of Protection against Salmonella Infections
3.1. Alteration in Bile Quality and Quantity
3.2. Gut Mucosa
3.3. Antimicrobial Activities
3.4. Gut Microbiome
3.5. Gut Immunity
4. Conclusions
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Herrick, R.L.; Buchberger, S.G.; Clark, R.M.; Kupferle, M.; Murray, R.; Succop, P. A Markov model to estimate Salmonella morbidity, mortality, illness duration, and cost. Health Econ. 2012, 21, 1169–1182. [Google Scholar] [CrossRef] [PubMed]
- Fouladkhah, A.; Geornaras, I.; Yang, H.; Belk, K.; Nightingale, K.K.; Woerner, D.; Smith, G.C.; Sofos, J.N. Sensitivity of Shiga Toxin-Producing Escherichia coli, Multidrug Resistant Salmonella, and Antibiotic Susceptible Salmonella to Lactic Acid on Inoculated Beef Trimmings. J. Food Prot. 2012, 75, 1751–1758. [Google Scholar] [CrossRef] [PubMed]
- Fouladkhah, A.; Geornaras, I.; Sofos, J. Biofilm Formation of O157 and Non-O157 Shiga Toxin-Producing Escherichia coli and Multidrug-Resistant and Susceptible Salmonella Typhimurium and Newport and Their Inactivation by Sanitizers. J. Food Sci. 2013, 78, M880–M886. [Google Scholar] [CrossRef] [PubMed]
- Fouladkhah, A.; Geornaras, I.; Yang, H.; Sofos, J. Lactic Acid Resistance of Shiga Toxin-Producing Escherichia coli and Multidrug-Resistant and Susceptible Salmonella Typhimurium and Salmonella Newport in Meat Homogenate. Food Microbiol. 2013, 36, 260–266. [Google Scholar] [CrossRef] [PubMed]
- Scallan, E.; Hoekstra, R.M.; Angulo, F.J.; Tauxe, R.V.; Widdowson, M.A.; Roy, S.L.; Jones, J.L.; Griffin, P.M. Foodborne Illness Acquired in the United States—major Pathogens. J. Emerg. Infect. Dis. 2011, 17, 7–15. [Google Scholar] [CrossRef] [PubMed]
- Scallan, E.; Hoekstra, R.M.; Mahon, B.E.; Jones, T.F.; Griffin, P.M. An Assessment of the Human Health Impact of Seven Leading Foodborne Pathogens in the United States Using Disability Adjusted Life Years. Epidemiol. Infect. 2015, 143, 2795–2804. [Google Scholar] [CrossRef] [PubMed]
- Bosilevac, J.M.; Arthur, T.M.; Bono, J.L.; Brichta-Harhay, D.M.; Kalchayanad, N.; King, D.A.; Shackelford, S.D.; Wheeler, M.L.; Koohmaraie, M. Prevalence and Enumeration of Escherichia coli O157: H7 and Salmonella in U.S. Abattoirs That Process Fewer than 1000 Head of Cattle per Day. J. Food Prot. 2009, 72, 1272–1278. [Google Scholar] [CrossRef] [PubMed]
- Zhao, S.; Blickenstaff, K.; Glenn, A.; Ayers, S.L.; Friedman, S.L.; Abbott, J.W.; McDermott, P.F. Lactam Resistance in Salmonella Strains Isolated from Retail Meats in the United States by the National Antimicrobial Resistance Monitoring System between 2002 and 2006. Appl. Environ. Microbiol. 2009, 75, 7624–7630. [Google Scholar] [CrossRef]
- Centers for Disease Control and Prevention (US). Antibiotic Resistance Threats in the United States; Centers for Disease Control and Prevention (US): Antlanta, GA, USA, 2013.
- Ao, T.T.; Feasey, N.A.; Gordon, M.A.; Keddy, K.H.; Angulo, F.J.; Crump, J.A. Global Burden of Invasive Nontyphoidal Salmonella Disease, 2010 (1). Emerg. Infect. Dis. 2015, 21, 941. [Google Scholar] [CrossRef]
- World Health Organization. Global Burden of Food Safety. Available online: Http://Www.Who.Int/Foodsafety/Areas_work/Foodborne-Diseases/Ferg/En/ (accessed on 1 January 2019).
- Grimont, P.A.; Weill, F.-X. Antigenic Formulae of the Salmonella Serovars. WHO Collab. Cent. Ref. Res. Salmonella 2007, 9, 1–166. [Google Scholar]
- Kearney, K. Food consumption trends and drivers. Philos. Trans. R. Soc. B 2010, 365, 2793–2807. [Google Scholar] [CrossRef] [PubMed]
- Cohen, M.L. Changing patterns of infectious disease. Nature 2000, 406, 762–767. [Google Scholar] [CrossRef] [PubMed]
- Thiennimitr, P.; Winter, S.E.; Winter, M.G.; Xavier, M.N.; Tolstikov, V.; Huseby, D.L.; Sterzenbach, T.; Tsolis, R.M.; Roth, J.R.; Bäumler, A.J. Intestinal inflammation allows Salmonella to use ethanolamine to compete with the microbiota. Proc. Natl. Acad. Sci. USA 2011, 108, 17480–17485. [Google Scholar] [CrossRef] [PubMed]
- Winter, S.E.; Thiennimitr, P.; Winter, M.G.; Butler, B.P.; Huseby, D.L.; Crawford, R.W.; Russell, J.M.; Bevins, C.L.; Adams, L.G.; Tsolis, R.M.; et al. Gut inflammation provides a respiratory electron acceptor for Salmonella. Nature 2010, 467, 426–429. [Google Scholar] [CrossRef] [PubMed]
- Stecher, B.; Robbiani, R.; Walker, A.W.; Westendorf, A.M.; Barthel, M.; Kremer, M.; Chaffron, S.; Macpherson, A.J.; Buer, J.; Parkhill, J.; et al. Salmonella enterica serovar typhimurium exploits inflammation to compete with the intestinal microbiota. PLoS Biol. 2007, 5, 2177–2189. [Google Scholar] [CrossRef] [PubMed]
- Calder, P.C.; Kew, S. The immune system: A target for functional foods? Br. J. Nutr. 2002, 88, S165–S177. [Google Scholar] [CrossRef]
- Harrison, L.M.; Balan, K.V.; Babu, U.S. Dietary fatty acids and immune response to food-borne bacterial infections. Nutrients 2013, 5, 1801–1822. [Google Scholar] [CrossRef] [PubMed]
- Taylor, A.K.; Cao, W.; Vora, K.P.; Cruz, J.D.L.; Shieh, W.J.; Zaki, S.R.; Katz, J.M.; Sambhara, S.; Gangappa, S. Protein energy malnutrition decreases immunity and increases susceptibility to influenza infection in mice. J. Infect. Dis. 2012, 207, 501–510. [Google Scholar] [CrossRef] [PubMed]
- Hekmatdoost, A.; Wu, X.; Morampudi, V.; Innis, S.M.; Jacobson, K. Dietary oils modify the host immune response and colonic tissue damage following Citrobacter rodentium infection in mice. Am. J. Physiol. 2013, 304, G917–G928. [Google Scholar] [CrossRef]
- Chandra, R.K. Nutrition, immunity and infection: From basic knowledge of dietary manipulation of immune responses to practical application of ameliorating suffering and improving survival. Proc. Natl. Acad. Sci. USA 1996, 93, 14304–14307. [Google Scholar] [CrossRef] [Green Version]
- He, R.R.; Wang, M.; Wang, C.Z.; Chen, B.T.; Lu, C.N.; Yao, X.S.; Chen, J.X.; Kurihara, H. Protective effect of apple polyphenols against stress-provoked influenza viral infection in restraint mice. J. Agric. Food Chem. 2011, 59, 3730–3737. [Google Scholar] [CrossRef]
- Daglia, M. Polyphenols as antimicrobial agents. Curr. Opin. Biotechnol. 2012, 23, 174–181. [Google Scholar] [CrossRef] [PubMed]
- Roberts, C.L.; Keita, Å.V.; Parsons, B.N.; Prorok-Hamon, M.; Knight, P.; Winstanley, C.; Niamh, O.; Söderholm, J.D.; Rhodes, J.M.; Campbell, B.J. Soluble plantain fibre blocks adhesion and M-cell translocation of intestinal pathogens. J. Nutr. Biochem. 2013, 24, 97–103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Long, K.Z.; Santos, J.I.; Rosado, J.L.; Estrada-Garcia, T.; Haas, M.; Al Mamun, A.; DuPont, H.L.; Nanthakumar, N.N. Vitamin A supplementation modifies the association between mucosal innate and adaptive immune responses and resolution of enteric pathogen infections. Am. J. Clin. Nutr. 2011, 93, 578–585. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dhaliwal, W.; Shawa, T.; Khanam, M.; Jagatiya, P.; Simuyandi, M.; Ndulo, N.; Bevins, C.L.; Sanderson, I.R.; Kelly, P. Intestinal antimicrobial gene expression: Impact of micronutrients in malnourished adults during a randomized trial. J. Infect. Dis. 2010, 202, 971–978. [Google Scholar] [CrossRef] [PubMed]
- Hung, C.C.; Garner, C.D.; Slauch, J.M.; Dwyer, Z.W.; Lawhon, S.D.; Frye, J.G.; McClelland, M.; Ahmer, B.M.; Altier, C. The intestinal fatty acid propionate inhibits Salmonella invasion through the post-translational control of HilD. Mol. Microbiol. 2013, 87, 1045–1060. [Google Scholar] [CrossRef] [PubMed]
- Agerberth, B.; Bergman, P.; Gudmundsson, G.H. Helping the host: Induction of antimicrobial peptides as a novel therapeutic strategy against infections. In Antimicrobial Peptides and Innate Immunity; Springer: Basel, Switzerland, 2013; Volume 14, pp. 359–375. [Google Scholar]
- Ulluwishewa, D.; Anderson, R.C.; McNabb, W.C.; Moughan, P.J.; Wells, J.M.; Roy, N.C. Regulation of tight junction permeability by intestinal bacteria and dietary components. J. Nutr. 2011, 141, 769–776. [Google Scholar] [CrossRef] [PubMed]
- Ramalingam, A.; Wang, X.; Gabello, M.; Valenzano, M.C.; Soler, A.P.; Ko, A.; Morin, P.J.; Mullin, J.M. Dietary methionine restriction improves colon tight junction barrier function and alters claudin expression pattern. Am. J. Physiol. Cell Physiol. 2010, 299, C1028–C1035. [Google Scholar] [CrossRef] [PubMed]
- Kau, A.L.; Ahern, P.P.; Griffin, N.W.; Goodman, A.L.; Gordon, J.I. Human nutrition, the gut microbiome and the immune system. Nature 2011, 474, 327–336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chai, S.J.; White, P.L.; Lathrop, S.L.; Solghan, S.M.; Medus, C.; McGlinchey, B.M.; Tobin-D’Angelo, M.; Marcus, R.; Mahon, B.E. Salmonella enterica Serotype Enteritidis: Increasing Incidence of Domestically Acquired Infections. Clin. Infect. Dis. 2012, 54, S488–S497. [Google Scholar] [CrossRef] [PubMed]
- Arguello, H.; Alvarez-Ordonez, A.; Carvajal, A.; Rubio, P.; Prieto, M. Role of Slaughtering in Salmonella Spreading and Control in Pork Production. J. Food Prot. 2013, 76, 899–911. [Google Scholar] [CrossRef] [PubMed]
- Metcalf, E.S.; Almond, G.W.; Routh, P.A.; Horton, J.R.; Dillman, R.C.; Orndorff, P.E. Experimental Salmonella typhi infection in the domestic pig, Sus scrofa domestica. Microb. Pathog. 2000, 29, 121–126. [Google Scholar] [CrossRef] [PubMed]
- Santos, R.L.; Zhang, S.; Tsolis, R.M.; Kingsley, R.A.; Adams, L.G.; Bäumler, A.J. Animal models of Salmonella infections: Enteritis versus typhoid fever. Microbes Infect. 2001, 3, 1335–1344. [Google Scholar] [CrossRef]
- Tsolis, R.M.; Kingsley, R.A.; Townsend, S.M.; Ficht, T.A.; Adams, L.G.; Bäumler, A.J. Of mice, calves, and men - Comparison of the mouse typhoid model with other Salmonella infections. Adv. Exp. Med. Biol. 1999, 473, 261–274. [Google Scholar] [PubMed]
- Oussalah, M.; Caillet, S.; Saucier, L.; Lacroix, M. Inhibitory effects of selected plant essential oils on the growth of four pathogenic bacteria: E. coli O157: H7, Salmonella Typhimurium, Staphylococcus aureus and Listeria monocytogenes. Food Control 2007, 18, 414–420. [Google Scholar] [CrossRef]
- Vikram, A.; Jesudhasan, P.R.; Jayaprakasha, G.K.; Pillai, S.D.; Jayaraman, A.; Patil, B.S. Citrus flavonoid represses Salmonella pathogenicity island 1 and motility in S. typhimurium LT2. Int. J. Food Microbiol. 2011, 145, 28–36. [Google Scholar] [CrossRef] [PubMed]
- Ebani, V.V.; Nardoni, S.; Bertelloni, F.; Tosi, G.; Massi, P.; Pistelli, L.; Mancianti, F. In Vitro Antimicrobial Activity of Essential Oils against Salmonella enterica Serotypes Enteritidis and Typhimurium Strains Isolated from Poultry. Molecules 2019, 24, 900. [Google Scholar] [CrossRef]
- Chen, C.Y.; Tsen, H.Y.; Lin, C.L.; Lin, C.K.; Chuang, L.T.; Chen, C.S.; Chiang, Y.C. Enhancement of the immune response against Salmonella infection of mice by heat-killed multispecies combinations of lactic acid bacteria. J. Med. Microbiol. 2013, 62, 1657–1664. [Google Scholar] [CrossRef]
- Forbes, S.J.; Eschmann, M.; Mantis, N.J. Inhibition of Salmonella enterica serovar Typhimurium motility and entry into epithelial cells by a protective antilipopolysaccharide monoclonal immunoglobulin a antibody. Infect. Immun. 2008, 76, 4137–4144. [Google Scholar] [CrossRef]
- Hansen-Wester, I.; Hensel, M. Salmonella pathogenicity islands encoding type III secretion systems. Microbes Infect. 2001, 3, 549–559. [Google Scholar] [CrossRef]
- Bovee-Oudenhoven, I.M.J.; ten Bruggencate, S.J.M.; Lettink-Wissink, M.L.G.; van der Meer, R. Dietary fructo-oligosaccharides and lactulose inhibit intestinal colonisation but stimulate translocation of Salmonella in rats. Gut 2003, 52, 1572–1578. [Google Scholar] [CrossRef] [PubMed]
- Hitchins, A.D.; Wells, P.; McDonough, F.E.; Wong, N.P. Amelioration of the adverse effect of a gastrointestinal challenge with Salmonella enteritidis on weanling rats by a yogurt diet. Am. J. Clin. Nutr. 1985, 41, 92–100. [Google Scholar] [CrossRef]
- Kim, S.P.; Moon, E.; Nam, S.H.; Friedman, M. Composition of Herba Pogostemonis Water Extract and Protection of Infected Mice against Salmonella typhimurium-Induced Liver Damage and Mortality by Stimulation of Innate Immune Cells. J. Agric. Food Chem. 2012, 60, 12122–12130. [Google Scholar] [CrossRef] [PubMed]
- Gabriel, P.O.; Aribisala, J.O.; Oladunmoye, M.K.; Arogunjo, A.O.; Ajayi-Moses, O.B. Therapeutic Effect of Goya Extra Virgin Olive Oil in Albino Rat Orogastricallly Dosed with Salmonella typhi. South Asian J. Res. Microbiol. 2019, 3, 1–9. [Google Scholar]
- Kelly, P.; Shawa, T.; Mwanamakondo, S.; Soko, R.; Smith, G.; Barclay, G.R.; Sanderson, I.R. Gastric and intestinal barrier impairment in tropical enteropathy and HIV: Limited impact of micronutrient supplementation during a randomised controlled trial. BMC Gastroenterol. 2010, 10, 72. [Google Scholar] [CrossRef]
- Tennant, S.M.; Hartland, E.L.; Phumoonna, T.; Lyras, D.; Rood, J.I.; Robins-Browne, R.M.; van Driel, I.R. Influence of gastric acid on susceptibility to infection with ingested bacterial pathogens. Infect. Immun. 2008, 76, 639–645. [Google Scholar] [CrossRef]
- Lucas, J.S.; Cochrane, S.A.; Warner, J.O.; Hourihane, J.O. The effect of digestion and pH on the allergenicity of kiwifruit proteins. Pediatr. Allergy Immunol.: Off. Publ. Eur. Soc. Pediatr. Allergy Immunol. 2008, 19, 392–398. [Google Scholar] [CrossRef]
- Henderson, L.M.; Brewer, G.J.; Dressman, J.B.; Swidan, S.Z.; DuRoss, D.J.; Adair, C.H.; Barnett, J.L.; Berardi, R.R. Effect of intragastric pH on the absorption of oral zinc acetate and zinc oxide in young healthy volunteers. JPEN J. Parenter. Enter. Nutr. 1995, 19, 393–397. [Google Scholar] [CrossRef]
- Kararli, T.T. Comparison of the Gastrointestinal Anatomy, Physiology, and Biochemistry of Humans and Commonly Used Laboratory-Animals. Biopharm. Drug Dispos. 1995, 16, 351–380. [Google Scholar] [CrossRef]
- Guilloteau, P.; Zabielski, R.; Hammon, H.M.; Metges, C.C. Nutritional programming of gastrointestinal tract development. Is the pig a good model for man? Nutr. Res. Rev. 2010, 23, 4–22. [Google Scholar] [CrossRef] [Green Version]
- Michiels, J.; Missotten, J.; Rasschaert, G.; Dierick, N.; Heyndrickx, M.; De Smet, S. Effect of organic acids on Salmonella colonization and shedding in weaned piglets in a seeder model. J. Food Prot. 2012, 75, 1974–1983. [Google Scholar] [CrossRef]
- Rajtak, U.; Boland, F.; Leonard, N.; Bolton, D.; Fanning, S. Roles of diet and the acid tolerance response in survival of common Salmonella serotypes in feces of finishing pigs. Appl. Environ. Microbiol. 2012, 78, 110–119. [Google Scholar] [CrossRef] [PubMed]
- Boyen, F.; Haesebrouck, F.; Vanparys, A.; Volf, J.; Mahu, M.; Van Immerseel, F.; Rychlik, I.; Dewulf, J.; Ducatelle, R.; Pasmans, F. Coated fatty acids alter virulence properties of Salmonella typhimurium and decrease intestinal colonization of pigs. Vet. Microbiol. 2008, 132, 319–327. [Google Scholar] [CrossRef]
- Chen, Y.; Chen, D.; Tian, G.; He, J.; Mao, X.; Mao, Q.; Yu, B. Dietary arginine supplementation alleviates immune challenge induced by Salmonella enterica serovar Choleraesuis bacterin potentially through the Toll-like receptor 4-myeloid differentiation factor 88 signalling pathway in weaned piglets. Br. J. Nutr. 2012, 108, 1069–1076. [Google Scholar] [CrossRef] [PubMed]
- Omonijo, F.A.; Ni, L.; Gong, J.; Wang, Q.; Lahaye, L.; Yang, C. Essential oils as alternatives to antibiotics in swine production. Anim. Nutr. 2018, 4, 126–136. [Google Scholar] [CrossRef] [PubMed]
- Boyen, F.; Haesebrouck, F.; Maes, D.; Van Immerseel, F.; Ducatelle, R.; Pasmans, F. Non-typhoidal Salmonella infections in pigs: A closer look at epidemiology, pathogenesis and control. Vet. Microbiol. 2008, 130, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Eberhard, M.; Hennig, U.; Kuhla, S.; Brunner, R.M.; Kleessen, B.; Metges, C.C. Effect of inulin supplementation on selected gastric, duodenal, and caecal microbiota and short chain fatty acid pattern in growing piglets. Arch. Anim. Nutr. 2007, 61, 235–246. [Google Scholar] [CrossRef] [PubMed]
- Scharek, L.; Tedin, K. The porcine immune system--differences compared to man and mouse and possible consequences for infections by Salmonella serovars. Berl. Und Munch. Tierarztl. Wochenschr. 2007, 120, 347–354. [Google Scholar]
- Higginson, E.E.; Simon, R.; Tennant, S.M. Animal models for salmonellosis: Applications in vaccine research. Clin. Vaccine Immunol. 2016, 23, 746–756. [Google Scholar] [CrossRef] [PubMed]
- Costa, L.F.; Paixao, T.A.; Tsolis, R.M.; Baumler, A.J.; Santos, R.L. Salmonellosis in cattle: Advantages of being an experimental model. Res. Vet. Sci. 2012, 93, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Hill, T.M.; VandeHaar, M.J.; Sordillo, L.M.; Catherman, D.R.; Bateman Ii, H.G.; Schlotterbeck, R.L. Fatty acid intake alters growth and immunity in milk-fed calves. J. Dairy Sci. 2011, 94, 3936–3948. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hofmann, R.R. Evolutionary Steps of Ecophysiological Adaptation and Diversification of Ruminants - a Comparative View of Their Digestive-System. Oecologia 1989, 78, 443–457. [Google Scholar] [CrossRef] [PubMed]
- Li, R.W.; Connor, E.E.; Li, C.J.; Baldwin, R.L.; Sparks, M.E. Characterization of the rumen microbiota of pre-ruminant calves using metagenomic tools. Environ. Microbiol. 2012, 14, 129–139. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Vargas, F.M.; Abu-El-Haija, M.A.; Gomez-Duarte, O.G. Salmonella infections: An update on epidemiology, management, and prevention. Travel Med. Infect. Dis. 2011, 9, 263–277. [Google Scholar] [CrossRef] [PubMed]
- Connor, B.A.; Schwartz, E. Typhoid and paratyphoid fever in travellers. Lancet Infect. Dis 2005, 5, 623–628. [Google Scholar] [CrossRef]
- Hohmann, E.L. Nontyphoidal salmonellosis. Clin. Infect. Dis. 2001, 32, 263–269. [Google Scholar] [PubMed]
- Centers for Disease Control and Prevention. National Outbreak Reporting System (NORS), 2018. Available online: https://www.cdc.gov/nors/index.html (accessed on 1 January 2019).
- Glenn, L.M.; Lindsey, R.L.; Folster, J.P.; Pecic, G.; Boerlin, P.; Gilmour, M.W. Antimicrobial resistance genes in multidrug-resistant Salmonella enterica isolated from animals, retail meats, and humans in the United States and Canada. Microb. Drug Resist. 2013, 19, 175–184. [Google Scholar] [CrossRef] [PubMed]
- Di Cagno, R.; De Angelis, M.; De Pasquale, I.; Ndagijimana, M.; Vernocchi, P.; Ricciuti, P.; Gagliardi, F.; Laghi, L.; Crecchio, C.; Guerzoni, M.E.; et al. Duodenal and faecal microbiota of celiac children: Molecular, phenotype and metabolome characterization. BMC Microbiol. 2011, 11, 219. [Google Scholar] [CrossRef]
- Lara-Villoslada, F.; Sierra, S.; Boza, J.; Xaus, J.; Olivares, M. Beneficial effects of consumption of a dairy product containing two probiotic strains, Lactobacillus coryniformis CECT5711 and Lactobacillus gasseri CECT5714 in healthy children. Nutr. Hosp. 2007, 22, 496–502. [Google Scholar]
- Rabbani, G.H.; Teka, T.; Zaman, B.; Majid, N.; Khatun, M.; Fuchs, G.J. Clinical studies in persistent diarrhea: Clinical studies in persistent diarrhea: Dietary management with green banana or pectin in Bangladeshi children. Gastroenterology 2001, 121, 554–560. [Google Scholar] [CrossRef]
- Kingamkono, R.; Sjogren, E.; Svanberg, U. Enteropathogenic bacteria in faecal swabs of young children fed on lactic acid-fermented cereal gruels. Epidemiol. Infect. 1999, 122, 23–32. [Google Scholar] [CrossRef] [PubMed]
- Szabó, I.; Wieler, L.H.; Tedin, K.; Scharek-Tedin, L.; Taras, D.; Hensel, A.; Appel, B.; Nöckler, K. Influence of a probiotic strain of Enterococcus faecium on Salmonella enterica serovar Typhimurium DT104 infection in a porcine animal infection model. Appl. Environ. Microbiol. 2009, 75, 2621–2628. [Google Scholar]
- Pasetti, M.F.; Levine, M.M.; Sztein, M.B. Animal models paving the way for clinical trials of attenuated Salmonella enterica serovar Typhi live oral vaccines and live vectors. Vaccine 2003, 21, 401–418. [Google Scholar] [CrossRef]
- Hofmann, A.F.; Hagey, R.L. Bile acids: Chemistry, pathochemistry, biology, pathobiology, and therapeutics. Cell Mol. Life Sci. 2008, 65, 2461–2483. [Google Scholar] [CrossRef]
- Prieto, A.I.; Ramos-Morales, F.; Casadesus, J. Bile-induced DNA damage in Salmonella enterica. Genetics 2004, 168, 1787–1794. [Google Scholar] [CrossRef]
- Merritt, M.E.; Donaldson, J.R. Effect of bile salts on the DNA and membrane integrity of enteric bacteria. J. Med. Microbiol. 2009, 58, 1533–1541. [Google Scholar] [CrossRef] [Green Version]
- Begley, M.; Gahan, C.G.M.; Hill, C. The interaction between bacteria and bile. FEMS Microbiol. Rev. 2005, 29, 625–651. [Google Scholar] [CrossRef] [Green Version]
- Antunes, L.C.M.; Andersen, S.K.; Menendez, A.; Arena, E.T.; Han, J.; Ferreira, R.B.; Borchers, C.H.; Finlay, B.B. Metabolomics Reveals Phospholipids as Important Nutrient Sources during Salmonella Growth in Bile in Vitro and in Vivo. J. Bacteriol. 2011, 193, 4719–4725. [Google Scholar] [CrossRef]
- Kollanoor-Johny, A.; Mattson, T.; Baskaran, S.A.; Amalaradjou, M.A.R.; Hoagland, T.A.; Darre, M.J.; Khan, M.I.; Schreiber, D.T.; Donoghue, A.M.; Venkitanarayanan, K. Caprylic acid reduces Salmonella enteritidis populations in various segments of digestive tract and internal organs of 3- and 6-week-old broiler chickens, therapeutically. Poult. Sci. 2012, 91, 1686–1694. [Google Scholar] [CrossRef]
- Inagaki, T.; Moschetta, A.; Lee, Y.K.; Peng, L.; Zhao, G.; Downes, M.; Ruth, T.Y.; Shelton, J.M.; Richardson, J.A.; Repa, J.J.; et al. Regulation of antibacterial defense in the small intestine by the nuclear bile acid receptor. Proc. Natl. Acad. Sci. USA 2006, 103, 3920–3925. [Google Scholar] [CrossRef] [Green Version]
- Xu, Q.; Xue, C.; Zhang, Y.; Liu, Y.; Wang, J.; Yu, X.; Zhang, X.; Zhang, R.; Yang, X.; Guo, C. Medium-Chain Fatty Acids Enhanced the Excretion of Fecal Cholesterol and Cholic Acid in C57BL/6J Mice Fed a Cholesterol-Rich Diet. Biosci. Biotechnol. Biochem. 2013, 77, 1390–1396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kato, T.; Hayashi, H.; Sugiyama, Y. Short- and medium-chain fatty acids enhance the cell surface expression and transport capacity of the bile salt export pump (BSEP/ABCB11). Biochim. Biophys. Acta 2010, 1801, 1005–1012. [Google Scholar] [CrossRef] [PubMed]
- Costarelli, V.; Sanders, T.A. Acute effects of dietary fat composition on postprandial plasma bile acid and cholecystokinin concentrations in healthy premenopausal women. Br. J. Nutr. 2001, 86, 471–477. [Google Scholar] [CrossRef]
- Yang, Q.; Lan, T.; Chen, Y.; Dawson, P.A. Dietary fish oil increases fat absorption and fecal bile acid content without altering bile acid synthesis in 20-d-old weanling rats following massive ileocecal resection. Pediatr. Res. 2012, 72, 38–42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prouty, A.M.; Brodsky, I.E.; Manos, J.; Belas, R.; Falkow, S.; Gunn, J.S. Transcriptional regulation of Salmonella enterica serovar typhimurium genes by bile. FEMS Immunol. Med. Microbiol. 2004, 41, 177–185. [Google Scholar] [CrossRef] [PubMed]
- Prouty, A.M.; Gunn, J.S. Salmonella enterica serovar typhimurium invasion is repressed in the presence of bile. Infect. Immun. 2000, 68, 6763–6769. [Google Scholar] [CrossRef]
- Ye, W.; Li, Y.; Zhou, Z.; Wang, X.; Yao, J.; Liu, J.; Wang, C. Synthesis and antibacterial activity of new long-chain-alkyl bile acid-based amphiphiles. Bioorg. Chem. 2013, 51, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Antunes, L.C.; Wang, M.; Andersen, S.K.; Ferreira, R.B.; Kappelhoff, R.; Han, J.; Borchers, C.H.; Finlay, B.B. Repression of Salmonella enterica phoP expression by small molecules from physiological bile. J. Bacteriol. 2012, 194, 2286–2296. [Google Scholar] [CrossRef]
- Matsumoto, K.; Yokoyama, S.; Gato, N. Bile acid-binding activity of young persimmon (Diospyros kaki) fruit and its hypolipidemic effect in mice. Phytother. Res. 2010, 24, 205–210. [Google Scholar] [CrossRef]
- Andersson, K.E.; Immerstrand, T.; Swärd, K.; Bergenståhl, B.; Lindholm, M.W.; Öste, R.; Hellstrand, P. Effects of oats on plasma cholesterol and lipoproteins in C57BL/6 mice are substrain specific. Br. J. Nutr. 2010, 103, 513–521. [Google Scholar] [CrossRef]
- Chen, H.L.; Lin, Y.M.; Wang, Y.C. Comparative effects of cellulose and soluble fibers (pectin, konjac glucomannan, inulin) on fecal water toxicity toward Caco-2 cells, fecal bacteria enzymes, bile acid, and short-chain fatty acids. J. Agric. Food Chem. 2010, 58, 10277–10281. [Google Scholar] [CrossRef]
- Gunness, P.; Gidley, M.J. Mechanisms underlying the cholesterol-lowering properties of soluble dietary fibre polysaccharides. Food Funct. 2010, 1, 149–155. [Google Scholar] [CrossRef] [PubMed]
- Hofmann, A.F.; Eckmann, L. How bile acids confer gut mucosal protection against bacteria. Proc. Natl. Acad. Sci. USA 2006, 103, 4333–4334. [Google Scholar] [CrossRef] [Green Version]
- Zarepour, M.; Bhullar, K.; Montero, M.; Ma, C.; Huang, T.; Velcich, A.; Xia, L.; Vallance, B.A. The mucin Muc2 limits pathogen burdens and epithelial barrier dysfunction during Salmonella enterica serovar Typhimurium Colitis. Infect. Immun. 2013, 81, 3672–3683. [Google Scholar] [CrossRef] [PubMed]
- Hansson, G.C. Role of mucus layers in gut infection and inflammation. Curr. Opin. Microbiol. 2012, 15, 57–62. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.S.; Ho, S.B. Intestinal goblet cells and mucins in health and disease: Recent insights and progress. Curr. Gastroenterol. Rep. 2010, 12, 319–330. [Google Scholar] [CrossRef]
- Kim, J.; Khan, W. Goblet Cells and Mucins: Role in Innate Defense in Enteric Infections. Pathogens 2013, 2, 55–70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Willemsen, L.E.; Koetsier, M.A.; van Deventer, S.J.; van Tol, E.A. Short chain fatty acids stimulate epithelial mucin 2 expression through differential effects on prostaglandin E (1) and E (2) production by intestinal myofibroblasts. Gut 2003, 52, 1442–1447. [Google Scholar] [CrossRef]
- Hino, S.; Takemura, N.; Sonoyama, K.; Morita, A.; Kawagishi, H.; Aoe, S.; Morita, T. Small intestinal goblet cell proliferation induced by ingestion of soluble and insoluble dietary fiber is characterized by an increase in sialylated mucins in rats. J. Nutr. 2012, 142, 1429–1436. [Google Scholar] [CrossRef]
- Kleessen, B.; Blaut, M. Modulation of gut mucosal biofilms. Br. J. Nutr. 2005, 93, S35–S40. [Google Scholar] [CrossRef] [Green Version]
- Morita, T.; Tanabe, H.; Takahashi, K.; Sugiyama, K. Ingestion of resistant starch protects endotoxin influx from the intestinal tract and reduces D-galactosamine-induced liver injury in rats. J. Gastroenterol. Hepatol. 2004, 19, 303–313. [Google Scholar] [CrossRef] [PubMed]
- Kosińska, A.; Andlauer, W. Modulation of Tight Junction Integrity by Food Components. Food Res. Int. 2013, 54, 951–960. [Google Scholar]
- Liu, J.H.; Xu, T.T.; Liu, Y.J.; Zhu, W.Y.; Mao, S.Y. A high-grain diet causes massive disruption of ruminal epithelial tight junctions in goats. Am. J. Psychol. Regul. Integr. Comp. Physiol. 2013, 305, R232–R241. [Google Scholar] [CrossRef] [PubMed]
- Nofrarias, M.; Martinez-Puig, D.; Pujols, J.; Majo, N.; Perez, J.F. Long-term intake of resistant starch improves colonic mucosal integrity and reduces gut apoptosis and blood immune cells. Nutrition 2007, 23, 861–870. [Google Scholar] [CrossRef] [PubMed]
- Park, E.J.; Thomson, A.B.; Clandinin, M.T. Protection of intestinal occludin tight junction protein by dietary gangliosides in lipopolysaccharide-induced acute inflammation. J. Pediatr. Gastroenterol. Nutr. 2010, 50, 321–328. [Google Scholar] [CrossRef]
- Suzuki, T.; Hara, H. Quercetin Enhances Intestinal Barrier Function through the Assembly of Zonnula Occludens-2, Occludin, and Claudin-1 and the Expression of Claudin-4 in Caco-2 Cells. J. Nutr. 2009, 139, 965–974. [Google Scholar] [CrossRef] [PubMed]
- Das, S.; Devaraj, S.N. Protective role of Hemidesmus indicus R. Br. root extract against Salmonella typhimurium-induced cytotoxicity in Int 407 cell line. Phytother. Res. 2007, 21, 1209–1216. [Google Scholar] [CrossRef] [PubMed]
- Willing, B.P.; Vacharaksa, A.; Croxen, M.; Thanachayanont, T.; Finlay, B.B. Altering Host Resistance to Infections through Microbial Transplantation. PLoS ONE 2011, 6, e26988. [Google Scholar] [CrossRef] [PubMed]
- Croswell, A.; Amir, E.; Teggatz, P.; Barman, M.; Salzman, N.H. Prolonged impact of antibiotics on intestinal microbial ecology and susceptibility to enteric Salmonella infection. Infect. Immun. 2009, 77, 2741–2753. [Google Scholar] [CrossRef]
- Kamada, N.; Chen, G.Y.; Inohara, N.; Nunez, G. Control of pathogens and pathobionts by the gut microbiota. Nat. Immunol. 2013, 14, 685–690. [Google Scholar] [CrossRef]
- Stelter, C.; Käppeli, R.; König, C.; Krah, A.; Hardt, W.D.; Stecher, B.; Bumann, D. Salmonella-induced mucosal lectin RegIIIbeta kills competing gut microbiota. PLoS ONE 2011, 6, e20749. [Google Scholar] [CrossRef]
- Gill, N.; Ferreira, R.B.; Antunes, L.C.M.; Willing, B.P.; Sekirov, I.; Al-Zahrani, F.; Hartmann, M.; Finlay, B.B. Neutrophil elastase alters the murine gut microbiota resulting in enhanced Salmonella colonization. PLoS ONE 2012, 7, e49646. [Google Scholar] [CrossRef]
- Castillo, N.A.; de Moreno de LeBlanc, A.; Galdeano, C.M.; Perdigón, G. Probiotics: An alternative strategy for combating salmonellosis: Immune mechanisms involved. Food Res. Int. 2012, 45, 831–841. [Google Scholar] [CrossRef]
- Burkholder, K.M.; Bhunia, A.K. Salmonella enterica serovar typhimurium adhesion and cytotoxicity during epithelial cell stress is reduced by Lactobacillus rhamnosus GG. Gut Pathog. 2009, 1, 14. [Google Scholar] [CrossRef]
- Castillo, N.A.; Perdigon, G.; de Moreno de LeBlanc, A. Oral administration of a probiotic Lactobacillus modulates cytokine production and TLR expression improving the immune response against Salmonella enterica serovar typhimurium infection in mice. BMC Microbiol. 2011, 11, 177. [Google Scholar] [CrossRef] [PubMed]
- Bayoumi, M.A.; Griffiths, M.W. Probiotics down-regulate genes in Salmonella enterica serovar typhimurium pathogenicity islands 1 and 2. J. Food Prot. 2010, 73, 452–460. [Google Scholar] [CrossRef]
- McNulty, N.P.; Wu, M.; Erickson, A.R.; Pan, C.; Erickson, B.; Martens, E.C.; Pudlo, N.A.; Muegge, B.D.; Henrissat, B.; Hettich, R.L.; et al. Effects of Diet on Resource Utilization by a Model Human Gut Microbiota Containing Bacteroides cellulosilyticus WH2, a Symbiont with an Extensive Glycobiome. PLoS Biol. 2013, 11, e1001637. [Google Scholar] [CrossRef] [PubMed]
- Turnbaugh, P.J.; Ridaura, V.K.; Faith, J.J.; Rey, F.E.; Knight, R.; Gordon, J.I. The Effect of Diet on the Human Gut Microbiome: A Metagenomic Analysis in Humanized Gnotobiotic Mice. Sci. Transl. Med. 2009, 1, 6–14. [Google Scholar] [CrossRef] [PubMed]
- Toward, R.; Montandon, S.; Walton, G.; Gibson, G.R. Effect of prebiotics on the human gut microbiota of elderly persons. Gut Microbes 2012, 3, 57–60. [Google Scholar] [CrossRef] [Green Version]
- Laparra, J.M.; Sanz, Y. Interactions of gut microbiota with functional food components and nutraceuticals. Pharmacol. Res.: Off. J. Ital. Pharmacol. Soc. 2010, 61, 219–225. [Google Scholar] [CrossRef] [Green Version]
- Yuan, Q.; Walker, W.A. Innate immunity of the gut: Mucosal defense in health and disease. J. Pediatr. Gastroenterol. Nutr. 2004, 38, 463–473. [Google Scholar] [CrossRef] [PubMed]
- Kawai, T.; Akira, S. The role of pattern-recognition receptors in innate immunity: Update on Toll-like receptors. Nat. Immunol. 2010, 11, 373–384. [Google Scholar] [CrossRef] [PubMed]
- Abreu, M.T. Toll-like receptor signalling in the intestinal epithelium: How bacterial recognition shapes intestinal function. Nat. Rev. Immunol. 2010, 10, 131–143. [Google Scholar] [CrossRef] [PubMed]
- Gordon, S. Pattern recognition receptors: Doubling up for the innate immune response. Cell 2002, 111, 927–930. [Google Scholar] [CrossRef]
- Macpherson, A.J.; Mccoy, K.D.; Johansen, F.E.; Brandtzaeg, P. The immune geography of IgA induction and function. Mucosal Immunol. 2008, 1, 11–22. [Google Scholar] [CrossRef] [PubMed]
- Fagarasan, S.; Honjo, T. Intestinal IgA synthesis: Regulation of front-line body defences. Nat. Rev. Immunol. 2003, 3, 63–72. [Google Scholar] [CrossRef] [PubMed]
- Nanton, M.R.; Way, S.S.; Shlomchik, M.J.; McSorley, S.J. Cutting edge: B cells are essential for protective immunity against Salmonella independent of antibody secretion. J. Immunol. 2012, 189, 5503–5507. [Google Scholar] [CrossRef] [PubMed]
- Cross, M.L. Microbes versus microbes: Immune signals generated by probiotic lactobacilli and their role in protection against microbial pathogens. Fems Immunol. Med. Microbiol. 2002, 34, 245–253. [Google Scholar] [CrossRef] [PubMed]
- Jouanguy, E.; DÖffinger, R.; Dupuis, S.; Pallier, A.; Altare, F.; Casanova, J.L. IL-12 and IFN-gamma in host defense against mycobacteria and salmonella in mice and men. Curr. Opin. Immunol. 1999, 11, 346–351. [Google Scholar] [CrossRef]
- Bao, S.; Beagley, K.W.; France, M.P.; Shen, J.; Husband, A.J. Interferon-gamma plays a critical role in intestinal immunity against Salmonella typhimurium infection. Immunology 2000, 99, 464–472. [Google Scholar] [CrossRef]
- Arnold, J.W.; Niesel, D.W.; Annable, C.R.; Hess, C.B.; Asuncion, M.; Cho, Y.J.; Peterson, J.W.; Klimpel, G.R. Tumor necrosis factor-alpha mediates the early pathology in Salmonella infection of the gastrointestinal tract. Microb. Pathog. 1993, 14, 217–227. [Google Scholar] [CrossRef] [PubMed]
- Kirby, A.C.; Yrlid, U.; Wick, M.J. The innate immune response differs in primary and secondary Salmonella infection. J. Immunol. 2002, 169, 4450–4459. [Google Scholar] [CrossRef] [PubMed]
- Mastroeni, P.; Harrison, J.A.; Chabalgoity, J.A.; Hormaeche, C.E. Effect of interleukin 12 neutralization on host resistance and gamma interferon production in mouse typhoid. Infect. Immun. 1996, 64, 189–196. [Google Scholar] [PubMed]
- Lapaque, N.; Walzer, T.; Meresse, S.; Vivier, E.; Trowsdale, J. Interactions between Human NK Cells and Macrophages in Response to Salmonella Infection. J. Immunol. 2009, 182, 4339–4348. [Google Scholar] [CrossRef]
- Mittrucker, H.W.; Kaufmann, S.H.E. Immune response to infection with Salmonella typhimurium in mice. J. Leukoc. Biol. 2000, 67, 457–463. [Google Scholar] [CrossRef] [PubMed]
- Michetti, P.; Mahan, M.J.; Slauch, J.M.; Mekalanos, J.J.; Neutra, M.R. Monoclonal Secretory Immunoglobulin-a Protects Mice against Oral Challenge with the Invasive Pathogen Salmonella-typhimurium. Infect. Immun. 1992, 60, 1786–1792. [Google Scholar] [PubMed]
- Schley, P.D.; Field, C.J. The immune-enhancing effects of dietary fibres and prebiotics. Br. J. Nutr. 2002, 87, 221–230. [Google Scholar] [CrossRef] [PubMed]
- Galdeano, C.M.; Núñez, I.N.; de Moreno de LeBlanc, A.; Carmuega, E.; Weill, R. Perdigón Impact of a probiotic fermented milk in the gut ecosystem and in the systemic immunity using a non-severe protein-energy-malnutrition model in mice. BMC Gastroenterol. 2011, 11, 64. [Google Scholar] [CrossRef] [PubMed]
- Guerrant, R.L.; Oria, R.B.; Moore, S.R.; Oria, M.O.B.; Lima, A.A.M. Malnutrition as an enteric infectious disease with long-term effects on child development. Nutr. Rev. 2008, 66, 487–505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghoneum, M.; Matsuura, M. Augmentation of macrophage phagocytosis by modified arabinoxylan rice bran (MGN-3/biobran). Int. J. Immunopathol. Pharm. 2004, 17, 283–292. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Niu, X.; Du, X.; Smith, D.; Meydani, S.N.; Wu, D. Dietary supplementation with white button mushrooms augments the protective immune response to salmonella vaccine in mice. J. Nutr. 2014, 144, 98–105. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Infection Model | Strength | Weaknesses |
|---|---|---|
| Salmonella Culture (Non-Host) | Direct interaction with the pathogen without confounders | Does not represent the interaction of dietary components with the host |
| Co-culture of Salmonella with host cell | Increased complexity of interaction compared to only pathogen culture, represents effects of intervention on the pathogen as well as the host | Does not represent involvement of all the host cell types that simultaneously happen together in human |
| Rodent Models | Represent a complex living system, very economical and convenient, ease in genetic manipulation to know mechanistic pathways | No diarrhea and vomiting, different intestinal immunity, different gastric environment, and anatomical structures |
| Pig Models | Similar to humans in body composition, cardiovascular, renal, nutritional, immunological, metabolic, and gastrointestinal aspects | Different than humans in Salmonella colonization pattern, gastric acidity, bile quantities, mucus thickness, immune system, not economical, not convenient |
| Calf Models | Develop similar clinical and pathological features such as diarrhea and enteritis | Stomach structure is different, not economical |
| Clinical Trials | The ideal model | Difficult to study the preventive effects of interventions due to ethical considerations |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kumar, A.; Allison, A.; Henry, M.; Scales, A.; Fouladkhah, A.C. Development of Salmonellosis as Affected by Bioactive Food Compounds. Microorganisms 2019, 7, 364. https://doi.org/10.3390/microorganisms7090364
Kumar A, Allison A, Henry M, Scales A, Fouladkhah AC. Development of Salmonellosis as Affected by Bioactive Food Compounds. Microorganisms. 2019; 7(9):364. https://doi.org/10.3390/microorganisms7090364
Chicago/Turabian StyleKumar, Ajay, Abimbola Allison, Monica Henry, Anita Scales, and Aliyar Cyrus Fouladkhah. 2019. "Development of Salmonellosis as Affected by Bioactive Food Compounds" Microorganisms 7, no. 9: 364. https://doi.org/10.3390/microorganisms7090364
APA StyleKumar, A., Allison, A., Henry, M., Scales, A., & Fouladkhah, A. C. (2019). Development of Salmonellosis as Affected by Bioactive Food Compounds. Microorganisms, 7(9), 364. https://doi.org/10.3390/microorganisms7090364