Microbial Markers Profile in Anaerobic Mars Analogue Environments Using the LDChip (Life Detector Chip) Antibody Microarray Core of the SOLID (Signs of Life Detector) Platform

 , , , , , , ,
, , , , , , , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Mars Analogues Selection and Sampling
2.2. Immunoprofiles of MASE Sites Using Fluorescence Sandwich Microarray Immunoassays
2.3. Building a Microarray from MASE Sites and Salty Environments
2.4. Testing and Validating the New Antibodies and MASE-Chip
3. Results
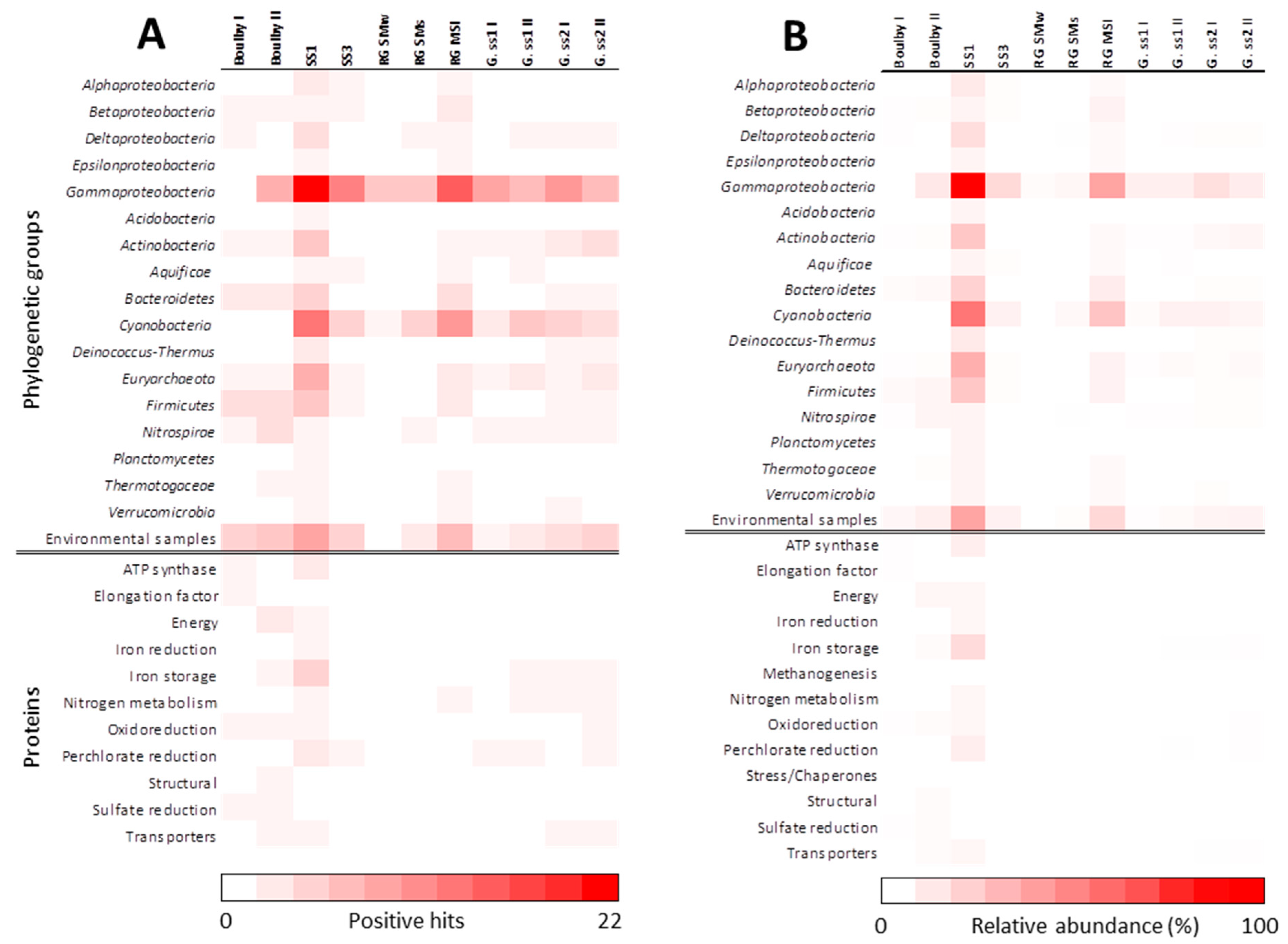
3.1. Biomarker Profiles of MASE Sites
3.2. MASE-Chip
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Cockell, C.S.; Bush, T.; Bryce, C.; Direito, S.; Fox-Powell, M.; Harrison, J.P.; Lammer, H.; Landenmark, H.; Martin-Torres, J.; Nicholson, N.; et al. Habitability: A Review. Astrobiology 2016, 16, 89–117. [Google Scholar] [CrossRef] [PubMed]
- Stoker, C.R.; Zent, A.; Catling, D.C.; Douglas, S.; Marshall, J.R.; Archer, D.; Clark, B.; Kounaves, S.P.; Lemmon, M.T.; Quinn, R.; et al. Habitability of the Phoenix landing site. J. Geophys. Res. 2010, 115. [Google Scholar] [CrossRef]
- Westall, F.; Loizeau, D.; Foucher, F.; Bost, N.; Betrand, M.; Vago, J.; Kminek, G. Habitability on Mars from a Microbial Point of View. Astrobiology 2013, 13, 887–897. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davila, A.F.; Duport, L.G.; Melchiorri, R.; Jänchen, J.; Valea, S.; Rios, A.d.l.; Fairén, A.G.; Möhlmann, D.; McKay, C.P.; Ascaso, C.; et al. Hygroscopic Salts and the Potential for Life on Mars. Astrobiology 2010, 10, 617–628. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davila, A.F.; Fairén, A.G.; Gago-Duport, L.; Stoker, C.; Amils, R.; Bonaccorsi, R.; Zavaleta, J.; Lim, D.; Schulze-Makuch, D.; McKay, C.P. Subsurface formation of oxidants on Mars and implications for the preservation of organic biosignatures. Earth Planet. Sci. Lett. 2008, 272, 456–463. [Google Scholar] [CrossRef]
- Fairén, A.G.; Davila, A.F.; Lim, D.; Bramall, N.; Bonaccorsi, R.; Zavaleta, J.; Uceda, E.R.; Stoker, C.; Wierzchos, J.; Dohm, J.M.; et al. Astrobiology through the Ages of Mars: The Study of Terrestrial Analogues to Understand the Habitability of Mars. Astrobiology 2010, 10, 821–843. [Google Scholar] [CrossRef]
- Horneck, G. The microbial world and the case for Mars. Planet. Space Sci. 2000, 48, 1053–1063. [Google Scholar] [CrossRef]
- Michalski, J.; Cuadros, J.; Niles, P.; Parnell, J.; Deanne Rogers, A.; Wright, S.P. Groundwater activity on Mars and implications for a deep biosphere. Nat. Geosci. 2013, 6, 133–138. [Google Scholar] [CrossRef]
- Schulze-Makuch, D.; Irwin, L.N.; Lipps, J.H.; LeMone, D.; Dohm, J.M.; Fairén, A.G. Scenarios for the evolution of life on Mars. J. Geophys. Res. 2005, 110. [Google Scholar] [CrossRef] [Green Version]
- Tosca, N.J.; Knoll, A.H.; McLennan, S.M. Water Activity and the Challenge for Life on Early Mars. Science 2008, 320, 1204–1207. [Google Scholar] [CrossRef]
- Gu, W.; Li, Y.; Tang, M.; Jia, X.; Ding, X.; Bi, X.; Wang, X. Water uptake and hygroscopicity of perchlorates and implications for the existence of liquid water in some hyperarid environments. Rsc. Adv. 2017, 7, 46866–46873. [Google Scholar] [CrossRef] [Green Version]
- Martín-Torres, F.J.; Zorzano, M.-P.; Valentín-Serrano, P.; Harri, A.-M.; Genzer, M.; Kemppinen, O.; Rivera-Valentin, E.G.; Jun, I.; Wray, J.; Bo Madsen, M.; et al. Transient liquid water and water activity at Gale crater on Mars. Nat. Geosci. 2015, 8, 357. [Google Scholar] [CrossRef]
- McEwen, A.S.; Ojha, L.; Dundas, C.M.; Mattson, S.S.; Byrne, S.; Wray, J.J.; Cull, S.C.; Murchie, S.L.; Thomas, N.; Gulick, V.C. Seasonal Flows on Warm Martian Slopes. Science 2011, 333, 740. [Google Scholar] [CrossRef] [PubMed]
- Bibring, J.-P.; Langevin, Y.; Mustard, J.F.; Poulet, F.; Arvidson, R.; Gendrin, A.; Gondet, B.; Mangold, N.; Pinet, P.; Forget, F.; et al. Global Mineralogical and Aqueous Mars History Derived from OMEGA/Mars Express Data. Science 2006, 312, 400. [Google Scholar] [CrossRef] [PubMed]
- Carr, M.H.; Head, J.W. Geologic history of Mars. Earth Planet. Sci. Lett. 2010, 294, 185–203. [Google Scholar] [CrossRef] [Green Version]
- McCauley, J.F. Mariner 9 evidence for wind erosion in the equatorial and mid-latitude regions of Mars. J. Geophys. Res. (1896–1977) 1973, 78, 4123–4137. [Google Scholar] [CrossRef]
- McEwen, A.S.; Eliason, E.M.; Bergstrom, J.W.; Bridges, N.T.; Hansen, C.J.; Delamere, W.A.; Grant, J.A.; Gulick, V.C.; Herkenhoff, K.E.; Keszthelyi, L.; et al. Mars Reconnaissance Orbiter’s High Resolution Imaging Science Experiment (HiRISE). J. Geophys. Res. 2007, 112. [Google Scholar] [CrossRef]
- Ojha, L.; Wilhelm, M.B.; Murchie, S.L.; McEwen, A.S.; Wray, J.J.; Hanley, J.; Massé, M.; Chojnacki, M. Spectral evidence for hydrated salts in recurring slope lineae on Mars. Nat. Geosci. 2015, 8, 829. [Google Scholar] [CrossRef]
- Stillman, D.E.; Michaels, T.I.; Grimm, R.E.; Harrison, K.P. New observations of martian southern mid-latitude recurring slope lineae (RSL) imply formation by freshwater subsurface flows. Icarus 2014, 233, 328–341. [Google Scholar] [CrossRef]
- Bryanskaya, A.; Rozanov, A.; Malup, T.; Aleshina, T.; Lazareva, E.; Taran, O.; Goryachkovskaya, T.; Ivanisenko, V.; Peltek, S. An Integrated Study to Analyze Salt Lake Microbial Community Structure (Novosibirsk Oblast, Russia). Acta Geol. Sin. Engl. Ed. 2014, 88, 61–62. [Google Scholar] [CrossRef]
- Litchfielld, C.D. Survival strategies for microorganisms in hypersaline environments and their relevance to life on early Mars. Meteorit. Planet. Sci. 1998, 33, 813–819. [Google Scholar] [CrossRef] [Green Version]
- Oren, A.; Elevi Bardavid, R.; Mana, L.J.E. Perchlorate and halophilic prokaryotes: implications for possible halophilic life on Mars. Extremophiles 2014, 18, 75–80. [Google Scholar] [CrossRef] [PubMed]
- Leuko, S.; Rothschild, L.J.; Burns, B.P. Halophilic Archaea and the Search for Extinct and Extant Life on Mars. J. Cosmol. 2010, 5, 940–950. [Google Scholar]
- Clark, B.C.; Baird, A.K.; Rose, H.J.; Toulmin, P.; Keil, K.; Castro, A.J.; Kelliher, W.C.; Rowe, C.D.; Evans, P.H. Inorganic Analyses of Martian Surface Samples at the Viking Landing Sites. Science 1976, 194, 1283–1288. [Google Scholar] [CrossRef] [PubMed]
- Clark, B.C.; Baird, A.K.; Weldon, R.J.; Tsusaki, D.M.; Schnabel, L.; Candelaria, M.P. Chemical composition of Martian fines. J. Geophys. Res. Solid Earth 1982, 87, 10059–10067. [Google Scholar] [CrossRef]
- Wänke, H.; Brückner, J.; Dreibus, G.; Rieder, R.; Ryabchikov, I. Chemical Composition of Rocks and Soils at the Pathfinder Site. Space Sci. Rev. 2001, 96, 317–330. [Google Scholar] [CrossRef]
- Brückner, J.; Dreibus, G.; Rieder, R.; Wänke, H. Refined data of Alpha Proton X-ray Spectrometer analyses of soils and rocks at the Mars Pathfinder site: Implications for surface chemistry. J. Geophys. Res. Planets 2003, 108. [Google Scholar] [CrossRef]
- Michalski, J.R.; Onstott, T.C.; Mojzsis, S.J.; Mustard, J.; Chan, Q.H.S.; Niles, P.B.; Johnson, S.S. The Martian subsurface as a potential window into the origin of life. Nat. Geosci. 2018, 11, 21–26. [Google Scholar] [CrossRef]
- Stamenković, V.; Ward, L.M.; Mischna, M.; Fischer, W.W. O2 solubility in Martian near-surface environments and implications for aerobic life. Nat. Geosci. 2018, 11, 905–909. [Google Scholar] [CrossRef] [Green Version]
- Cockell, C.S.; Schwendner, P.; Perras, A.; Rettberg, P.; Beblo-Vranesevic, K.; Bohmeier, M.; Rabbow, E.; Moissl-Eichinger, C.; Wink, L.; Marteinsson, V.; et al. Anaerobic microorganisms in astrobiological analogue environments: from field site to culture collection. Int. J. Astrobiol. 2018, 17, 314–328. [Google Scholar] [CrossRef]
- Perras, A.K. Grappling extremes: Molecular methods combined with cultivation reveal the composition and biology of space-relevant microbial communities. Ph.D. Thesis, University of Regensburg, Regensburg, Germany, 15 January 2018. [Google Scholar]
- Beblo-Vranesevic, K.; Bohmeier, M.; Perras, A.K.; Schwendner, P.; Rabbow, E.; Moissl-Eichinger, C.; Cockell, C.S.; Pukall, R.; Vannier, P.; Marteinsson, V.T.; et al. The responses of an anaerobic microorganism, Yersinia intermedia MASE-LG-1 to individual and combined simulated Martian stresses. PLoS ONE 2017, 12, e0185178. [Google Scholar] [CrossRef] [PubMed]
- Gaboyer, F.; Le Milbeau, C.; Bohmeier, M.; Schwendner, P.; Vannier, P.; Beblo-Vranesevic, K.; Rabbow, E.; Foucher, F.; Gautret, P.; Guégan, R.; et al. Mineralization and Preservation of an extremotolerant Bacterium Isolated from an Early Mars Analog Environment. Sci. Rep. 2017, 7, 8775. [Google Scholar] [CrossRef] [PubMed]
- Beblo-Vranesevic, K.; Bohmeier, M.; Perras, A.K.; Schwendner, P.; Rabbow, E.; Moissl-Eichinger, C.; Cockell, C.S.; Vannier, P.; Marteinsson, V.T.; Monaghan, E.P.; et al. Lack of correlation of desiccation and radiation tolerance in microorganisms from diverse extreme environments tested under anoxic conditions. FEMS Microbiol. Lett. 2018, 365, fny044. [Google Scholar] [CrossRef] [PubMed]
- Schwendner, P.; Bohmeier, M.; Rettberg, P.; Beblo-Vranesevic, K.; Gaboyer, F.; Moissl-Eichinger, C.; Perras, A.K.; Vannier, P.; Marteinsson, V.T.; Garcia-Descalzo, L.; et al. Beyond Chloride Brines: Variable Metabolomic Responses in the Anaerobic Organism Yersinia intermedia MASE-LG-1 to NaCl and MgSO4 at Identical Water Activity. Front. Microbiol. 2018, 9. [Google Scholar] [CrossRef] [PubMed]
- Parro, V.; Rodríguez-Manfredi, J.A.; Briones, C.; Compostizo, C.; Herrero, P.L.; Vez, E.; Sebastián, E.; Moreno-Paz, M.; García-Villadangos, M.; Fernández-Calvo, P.; et al. Instrument development to search for biomarkers on mars: Terrestrial acidophile, iron-powered chemolithoautotrophic communities as model systems. Planet. Space Sci. 2005, 53, 729–737. [Google Scholar] [CrossRef]
- Sims, M.R.; Cullen, D.C.; Bannister, N.P.; Grant, W.D.; Henry, O.; Jones, R.; McKnight, D.; Thompson, D.P.; Wilson, P.K. The specific molecular identification of life experiment (SMILE). Planet. Space Sci. 2005, 53, 781–791. [Google Scholar] [CrossRef]
- Parro, V.; Diego-Castilla, G.d.; Rodríguez-Manfredi, J.A.; Rivas, L.A.; Blanco-López, Y.; Sebastián, E.; Romeral, J.; Compostizo, C.; Herrero, P.L.; García-Marín, A.; et al. SOLID3: A Multiplex Antibody Microarray-Based Optical Sensor Instrument for In Situ Life Detection in Planetary Exploration. Astrobiology 2011, 11, 15–28. [Google Scholar] [CrossRef] [PubMed]
- Sephton, M.A.; Sims, M.R.; Court, R.W.; Luong, D.; Cullen, D.C. Searching for biomolecules on Mars: Considerations for operation of a life marker chip instrument. Planet. Space Sci. 2013, 86, 66–74. [Google Scholar] [CrossRef]
- Parro, V.; Diego-Castilla, G.d.; Moreno-Paz, M.; Blanco, Y.; Cruz-Gil, P.; Rodríguez-Manfredi, J.A.; Fernández-Remolar, D.; Gómez, F.; Gómez, M.J.; Rivas, L.A.; et al. A Microbial Oasis in the Hypersaline Atacama Subsurface Discovered by a Life Detector Chip: Implications for the Search for Life on Mars. Astrobiology 2011, 11, 969–996. [Google Scholar] [CrossRef]
- Parro, V.; Rivas, L.A.; Gómez-Elvira, J. Protein Microarrays-Based Strategies for Life Detection in Astrobiology. In Strategies of Life Detection; Botta, O., Bada, J.L., Gomez-Elvira, J., Javaux, E., Selsis, F., Summons, R., Eds.; Springer: Boston, MA, USA, 2008; Volume 25, pp. 293–311. [Google Scholar]
- Blanco, Y.; Gallardo-Carreño, I.; Ruiz-Bermejo, M.; Puente-Sánchez, F.; Cavalcante-Silva, E.; Quesada, A.; Prieto-Ballesteros, O.; Parro Garcia, V. Critical Assessment of Analytical Techniques in the Search for Biomarkers on Mars: A Mummified Microbial Mat from Antarctica as a Best-Case Scenario. Astrobiology 2017, 17. [Google Scholar] [CrossRef]
- Parro, V.; Blanco, Y.; Puente-Sánchez, F.; Rivas, L.A.; Moreno-Paz, M.; Echeverría, A.; Chong-Díaz, G.; Demergasso, C.; Cabrol Nathalie, A. Biomarkers and Metabolic Patterns in the Sediments of Evolving Glacial Lakes as a Proxy for Planetary Lake Exploration. Astrobiology 2018, 18, 586–606. [Google Scholar] [CrossRef] [PubMed]
- Parro, V.; Fernández-Remolar, D.; Rodríguez-Manfredi, J.A.; Cruz-Gil, P.; Rivas, L.A.; Ruiz-Bermejo, M.; Moreno-Paz, M.; García-Villadangos, M.; Gómez-Ortiz, D.; Blanco-López, Y.; et al. Classification of Modern and Old Río Tinto Sedimentary Deposits Through the Biomolecular Record Using a Life Marker Biochip: Implications for Detecting Life on Mars. Astrobiology 2011, 11, 29–44. [Google Scholar] [CrossRef] [PubMed]
- Puente-Sánchez, F.; Sánchez-Román, M.; Amils, R.; Parro Garcia, V. Tessaracoccus lapidicaptus sp. nov., an actinobacterium isolated from the deep subsurface of the Iberian pyrite belt. Int. J. Syst. Evol. Microbiol. 2014, 64, 3546–3552. [Google Scholar] [CrossRef] [Green Version]
- Puente-Sánchez, F.; Arce Rodríguez, A.; Oggerin, M.; García-Villadangos, M.; Moreno-Paz, M.; Blanco, Y.; Rodríguez, N.; Bird, L.; Lincoln, S.; Tornos, F.; et al. Viable cyanobacteria in the deep continental subsurface. Proc. Natl. Acad. Sci. USA 2018, 115. [Google Scholar] [CrossRef]
- Blanco, Y.; Prieto-Ballesteros, O.; Gómez, M.J.; Moreno-Paz, M.; García-Villadangos, M.; Rodríguez-Manfredi, J.A.; Cruz-Gil, P.; Sánchez-Román, M.; Rivas, L.A.; Parro, V. Prokaryotic communities and operating metabolisms in the surface and the permafrost of Deception Island (Antarctica). Environ. Microbiol. 2012, 14, 2495–2510. [Google Scholar] [CrossRef] [PubMed]
- Parro, V.; Puente-Sánchez, F.; Cabrol, N.A.; Gallardo-Carreño, I.; Moreno-Paz, M.; Blanco, Y.; García-Villadangos, M.; Tambley, C.; Tilot, V.C.; Thompson, C.; et al. Microbiology and Nitrogen Cycle in the Benthic Sediments of a Glacial Oligotrophic Deep Andean Lake as Analog of Ancient Martian Lake-Beds. Front. Microbiol. 2019, 10. [Google Scholar] [CrossRef]
- Woods, P.J.E. The geology of Boulby Mine. Econ. Geol. 1979, 74, 409–418. [Google Scholar] [CrossRef]
- McGenity, T.J.; Gemmell, R.T.; Grant, W.D.; Stan-Lotter, H. Origins of halophilic microorganisms in ancient salt deposits. Environ. Microbiol. 2000, 2, 243–250. [Google Scholar] [CrossRef]
- Cockell, C.S.; Payler, S.; Paling, S.; McLuckie, D. Boulby International Subsurface Astrobiology LaboratoryBoulby Mine. Astron. Geophys. 2013, 54, 2.25–2.27. [Google Scholar] [CrossRef]
- Payler, S.J.; Biddle, J.F.; Coates, A.J.; Cousins, C.R.; Cross, R.E.; Cullen, D.C.; Downs, M.T.; Direito, S.O.L.; Edwards, T.; Gray, A.L.; et al. Planetary science and exploration in the deep subsurface: Results from the MINAR Program, Boulby Mine, UK. Int. J. Astrobiol. 2017, 16, 114–129. [Google Scholar] [CrossRef]
- Thorarinsson, S. Grænavatn and Gestsstadavatn. Geogr. Tidsskr. 1953, 52, 292–302. [Google Scholar]
- Bhattacharya, J.P.; Payenberg, T.H.D.; Lang, S.C.; Bourke, M. Dynamic river channels suggest a long-lived Noachian crater lake on Mars. Geophys. Res. Lett. 2005, 32. [Google Scholar] [CrossRef] [Green Version]
- Ehlmann, B.L.; Mustard, J.F.; Fassett, C.I.; Schon, S.C.; Head Iii, J.W.; Des Marais, D.J.; Grant, J.A.; Murchie, S.L. Clay minerals in delta deposits and organic preservation potential on Mars. Nat. Geosci. 2008, 1, 355. [Google Scholar] [CrossRef]
- Rudolph, C.; Wanner, G.; Huber, R. Natural Communities of Novel Archaea and Bacteria Growing in Cold Sulfurous Springs with a String-of-Pearls-Like Morphology. Appl. Environ. Microbiol. 2001, 67, 2336–2344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Henneberger, R.; Moissl, C.; Amann, T.; Rudolph, C.; Huber, R. New insights into the lifestyle of the cold-loving SM1 euryarchaeon: natural growth as a monospecies biofilm in the subsurface. Appl. Environ. Microbiol. 2006, 72, 192–199. [Google Scholar] [CrossRef]
- Moissl, C.; Rudolph, C.; Huber, R. Natural communities of novel archaea and bacteria with a string-of-pearls-like morphology: molecular analysis of the bacterial partners. Appl. Environ. Microbiol. 2002, 68, 933–937. [Google Scholar] [CrossRef]
- Lemcke, K. Übertiefe Grundwässer im süddeutschen Alpenvorland. Buu Ver Schweiz Pet. -Geol U -Ing 1976, 42, 9–18. [Google Scholar]
- Probst, A.J.; Birarda, G.; Holman, H.-Y.N.; DeSantis, T.Z.; Wanner, G.; Andersen, G.L.; Perras, A.K.; Meck, S.; Völkel, J.; Bechtel, H.A.; et al. Coupling genetic and chemical microbiome profiling reveals heterogeneity of archaeome and bacteriome in subsurface biofilms that are dominated by the same archaeal species. PLoS ONE 2014, 9, e99801. [Google Scholar] [CrossRef]
- Gaillard, F.; Michalski, J.; Berger, G.; Maclennan, S.M.; Scaillet, B. Geochemical Reservoirs and Timing of Sulfur Cycling on Mars. Space Sci. Rev. 2013, 174, 251–300. [Google Scholar] [CrossRef]
- Rivas, L.A.; García-Villadangos, M.; Moreno-Paz, M.; Cruz-Gil, P.; Gómez-Elvira, J.; Parro, V. A 200-Antibody Microarray Biochip for Environmental Monitoring: Searching for Universal Microbial Biomarkers through Immunoprofiling. Anal. Chem. 2008, 80, 7970–7979. [Google Scholar] [CrossRef] [Green Version]
- Blanco, Y.; Moreno-Paz, M.; Aguirre, J.; Parro, V. Multiplex Fluorescent Antibody Microarrays and Antibody Graphsfor Microbial and Biomarker Detection in the Environment. In Hydrocarbon and Lipid Microbiology Protocol; McGenity, T.J., Timmis, K.N., Fernandez, B.N., Eds.; Springer: Berlin/Heidelberg, Germany, 2015. [Google Scholar]
- Nocker, A.; Sossa-Fernandez, P.; Burr, M.D.; Camper, A.K. Use of propidium monoazide for live/dead distinction in microbial ecology. Appl. Environ. Microbiol. 2007, 73, 5111–5117. [Google Scholar] [CrossRef] [PubMed]
- Barth, V.C.; Cattani, F.; Ferreira, C.A.S.; de Oliveira, S.D. Sodium chloride affects propidium monoazide action to distinguish viable cells. Anal. Biochem. 2012, 428, 108–110. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-García, L.; Aeppli, C.; Parro, V.; Fernández-Remolar, D.; García-Villadangos, M.; Chong-Diaz, G.; Blanco, Y.; Carrizo, D.J.B. Molecular biomarkers in the subsurface of the Salar Grande (Atacama, Chile) evaporitic deposits. Biogeochemistry 2018, 140, 31–52. [Google Scholar] [CrossRef] [Green Version]
- Blanco, Y.; Rivas, L.A.; García-Moyano, A.; Aguirre, J.; Cruz-Gil, P.; Palacín, A.; van Heerden, E.; Parro, V. Deciphering the prokaryotic community and metabolisms in South African deep-mine biofilms through antibody microarrays and graph theory. PLoS ONE 2014, 9, e114180. [Google Scholar] [CrossRef] [PubMed]
- García-Descalzo, L.; García-López, E.; Alcázar, A.; Baquero, F.; Cid, C. Proteomic analysis of the adaptation to warming in the Antarctic bacteria Shewanella frigidimarina. Biochim. Et Biophys. Acta (Bba) Proteins Proteom. 2014, 1844, 2229–2240. [Google Scholar] [CrossRef]
- Feller, G. Cryosphere and Psychrophiles: Insights into a Cold Origin of Life? Life (Basel) 2017, 7, 25. [Google Scholar] [CrossRef] [PubMed]
- Vorobyova, E.; Soina, V.; Gorlenko, M.; Minkovskaya, N.; Zalinova, N.; Mamukelashvili, A.; Gilichinsky, D.; Rivkina, E.; Vishnivetskaya, T. The deep cold biosphere: facts and hypothesis. Fems Microbiol. Rev. 1997, 20, 277–290. [Google Scholar] [CrossRef]
- Antony, R.; Krishnan, K.P.; Laluraj, C.M.; Thamban, M.; Dhakephalkar, P.K.; Engineer, A.S.; Shivaji, S. Diversity and physiology of culturable bacteria associated with a coastal Antarctic ice core. Microbiol. Res. 2012, 167, 372–380. [Google Scholar] [CrossRef] [PubMed]
- Marx, J.G.; Carpenter, S.D.; Deming, J.W. Production of cryoprotectant extracellular polysaccharide substances (EPS) by the marine psychrophilic bacterium Colwellia psychrerythraea strain 34H under extreme conditions. Can. J. Microbiology. 2009, 55, 63–72. [Google Scholar] [CrossRef]
- Cousins, C. Volcanogenic Fluvial-Lacustrine Environments in Iceland and Their Utility for Identifying Past Habitability on Mars. Life (Basel) 2015, 5, 568–586. [Google Scholar] [CrossRef] [Green Version]
- Johnston, C.A. Sediment and nutrient retention by freshwater wetlands: Effects on surface water quality. Crit. Rev. Environ. Control 1991, 21, 491–565. [Google Scholar] [CrossRef]
- Knox, A.K.; Dahlgren, R.A.; Tate, K.W.; Atwill, E.R. Efficacy of Natural Wetlands to Retain Nutrient, Sediment and Microbial Pollutants. J. Environ. Qual. 2008, 37, 1837–1846. [Google Scholar] [CrossRef] [PubMed]
- Blanco, Y.; de Diego-Castilla, G.; Viúdez-Moreiras, D.; Cavalcante-Silva, E.; Rodríguez-Manfredi, J.A.; Davila, A.F.; McKay, C.P.; Parro, V. Effects of Gamma and Electron Radiation on the Structural Integrity of Organic Molecules and Macromolecular Biomarkers Measured by Microarray Immunoassays and Their Astrobiological Implications. Astrobiology 2018, 18, 1497–1516. [Google Scholar] [CrossRef] [PubMed]
- Juncker, D.; Bergeron, S.; Laforte, V.; Li, H. Cross-reactivity in antibody microarrays and multiplexed sandwich assays: shedding light on the dark side of multiplexing. Curr. Opin. Chem. Biol. 2014, 18, 29–37. [Google Scholar] [CrossRef] [PubMed]
- Rivas, L.A.; Aguirre, J.; Blanco, Y.; González-Toril, E.; Parro, V. Graph-based deconvolution analysis of multiplex sandwich microarray immunoassays: applications for environmental monitoring. Environ. Microbiol. 2011, 13, 1421–1432. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ab Name/ID | Immunogen (Sample/Strain) | References |
|---|---|---|
| Bou. I | Boulby Mine I (wet and pink salt) | - |
| Bou. II | Boulby Mine II | - |
| MSIs | Mülbach Islinger-Regensburg (cold spring-sediment) | - |
| SMs | Sippenauer Moore-Regensburg (cold spring-sediment) | - |
| IS. SS1 | Grænavatn Lake-Iceland (sediment) | - |
| MASE-BB-1 | Halanaerobium sp. (isolate from Boulby) | [30] |
| MASE-IM-5 | Trichococcus sp. 37AN3 (Mülbach Islinger) | [30] |
| MASE-IM-4 | Clostridium sp. DSM632 (Mülbach Islinger) | [30] |
| MASE-SM-3 | Hafnia sp. (Sippenauer Moore) | [30] |
| MASE-SM-2 | Clostridium sp. (Sippenauer Moore) | [30] |
| MASE-SM-1 | Methanomethylovorans sp. (Sippenauer Moore) | [30] |
| MASE-IM-7 | Desulfovibrio sp. (Mülbach Islinger) | [30] |
| MASE-LG-2 | Pelosinus sp. (Grænavatn Lake) | [30] |
| ET2 | Bacteroides xylanoliticus X5-1 (Grænavatn Lake) | up |
| MASE-Glacier-SS3 | Rhanella sp. (Kaunertal Glacier) | [30] |
| IVE7C1 | Halothiobacillus neapolitanus | [40] |
| IVG5C1 | Sulfobacillus acidophilus | [40] |
| IVI12C1 | Geobacter metallireducens | [62] |
| IVI20C1 | Salinibacter ruber M8 | [40] |
| IVI21C1 | Salinibacter ruber PR1 | [40] |
| IVI24C1 | Thessaracoccus lapidicapta | [66] |
| IVJ1C1 | Haloferax mediterranei | [62] |
| IVJ8C1 | Halorubrum sp. | [40] |
| IVJ9C1 | Halobacterium sp. | [40] |
| IVK19C1 | Chroococcidiopsis O29 | up |
| VD2BF | Biofilm from Mansimongs Mines Southafrica | up |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
García-Descalzo, L.; Parro, V.; García-Villadangos, M.; Cockell, C.S.; Moissl-Eichinger, C.; Perras, A.; Rettberg, P.; Beblo-Vranesevic, K.; Bohmeier, M.; Rabbow, E.; et al. Microbial Markers Profile in Anaerobic Mars Analogue Environments Using the LDChip (Life Detector Chip) Antibody Microarray Core of the SOLID (Signs of Life Detector) Platform. Microorganisms 2019, 7, 365. https://doi.org/10.3390/microorganisms7090365
García-Descalzo L, Parro V, García-Villadangos M, Cockell CS, Moissl-Eichinger C, Perras A, Rettberg P, Beblo-Vranesevic K, Bohmeier M, Rabbow E, et al. Microbial Markers Profile in Anaerobic Mars Analogue Environments Using the LDChip (Life Detector Chip) Antibody Microarray Core of the SOLID (Signs of Life Detector) Platform. Microorganisms. 2019; 7(9):365. https://doi.org/10.3390/microorganisms7090365
Chicago/Turabian StyleGarcía-Descalzo, Laura, Victorino Parro, Miriam García-Villadangos, Charles S. Cockell, Christine Moissl-Eichinger, Alex Perras, Petra Rettberg, Kristina Beblo-Vranesevic, Maria Bohmeier, Elke Rabbow, and et al. 2019. "Microbial Markers Profile in Anaerobic Mars Analogue Environments Using the LDChip (Life Detector Chip) Antibody Microarray Core of the SOLID (Signs of Life Detector) Platform" Microorganisms 7, no. 9: 365. https://doi.org/10.3390/microorganisms7090365
APA StyleGarcía-Descalzo, L., Parro, V., García-Villadangos, M., Cockell, C. S., Moissl-Eichinger, C., Perras, A., Rettberg, P., Beblo-Vranesevic, K., Bohmeier, M., Rabbow, E., Westall, F., Gaboyer, F., Amils, R., Malki, M., Marteinsson, V., Vannier, P., Ehrenfreund, P., Monaghan, E., Riedo, A., ... Gómez, F. G. (2019). Microbial Markers Profile in Anaerobic Mars Analogue Environments Using the LDChip (Life Detector Chip) Antibody Microarray Core of the SOLID (Signs of Life Detector) Platform. Microorganisms, 7(9), 365. https://doi.org/10.3390/microorganisms7090365