Human Polyomaviruses in the Cerebrospinal Fluid of Neurological Patients

 , , , ,
, , , , 

Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Group
2.2. DNA Isolation
2.3. HPyV Quantitative Real-Time PCR (Q-PCR)
2.4. JCPyV, BKPyV and MCPyV Molecular Characterization
2.5. Statistical Analysis
3. Results
3.1. HPyV Detection by Q-PCR
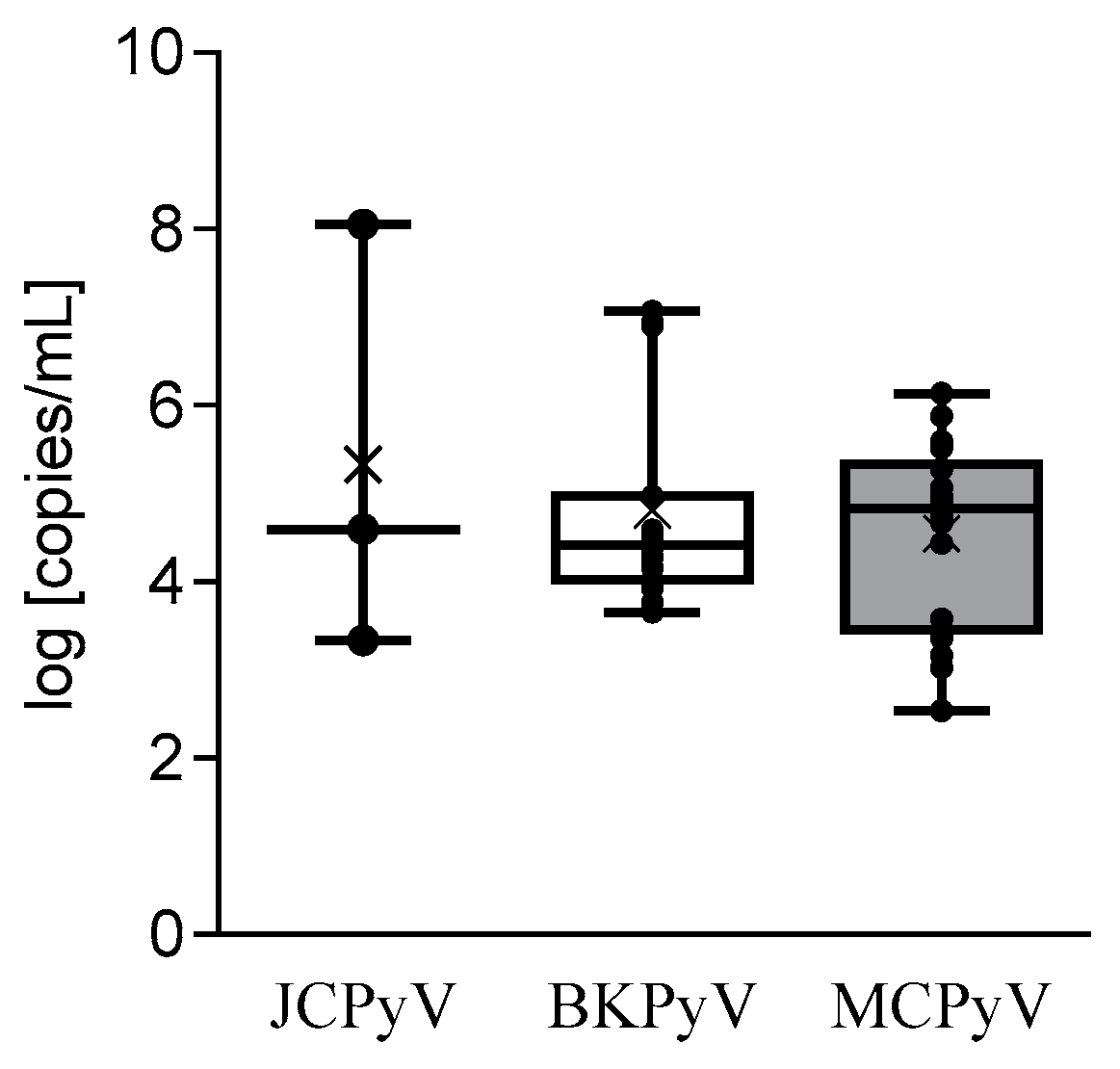
3.2. HPyV Load and Infected Cells Percentage
3.3. HPyV Distribution in the CSF According to the Disease
3.4. HPyV Molecular Characterization
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- White, M.K.; Safak, M.; Khalili, K. Regulation of gene expression in primate polyomaviruses. J. Virol. 2009, 83, 10846–10856. [Google Scholar] [CrossRef] [Green Version]
- Allander, T.; Andreasson, K.; Gupta, S.; Bjerkner, A.; Bogdanovic, G.; Persson, M.A.; Dalianis, T.; Ramqvist, T.; Andersson, B. Identification of a third human polyomavirus. J. Virol. 2007, 81, 4130–4136. [Google Scholar] [CrossRef] [Green Version]
- Gaynor, A.M.; Nissen, M.D.; Whiley, D.M.; Mackay, I.M.; Lambert, S.B.; Wu, G.; Brennan, D.C.; Storch, G.A.; Sloots, T.P.; Wang, D. Identification of a novel polyomavirus from patients with acute respiratory tract infections. PLoS Pathog. 2007, 3, e64. [Google Scholar] [CrossRef]
- Feng, H.; Kwun, H.J.; Liu, X.; Gjoerup, O.; Stolz, D.B.; Chang, Y.; Moore, P.S. Cellular and viral factors regulating Merkel cell polyomavirus replication. PLoS ONE 2011, 6, e22468. [Google Scholar] [CrossRef] [Green Version]
- Feng, H.; Shuda, M.; Chang, Y.; Moore, P.S. Clonal integration of a polyomavirus in human Merkel cell carcinoma. Science 2008, 319, 1096–1100. [Google Scholar] [CrossRef] [Green Version]
- van der Meijden, E.; Janssens, R.W.; Lauber, C.; Bouwes Bavinck, J.N.; Gorbalenya, A.E.; Feltkamp, M.C. Discovery of a new human polyomavirus associated with trichodysplasia spinulosa in an immunocompromized patient. PLoS Pathog. 2010, 6, e1001024. [Google Scholar] [CrossRef] [Green Version]
- Schowalter, R.M.; Pastrana, D.V.; Pumphrey, K.A.; Moyer, A.L.; Buck, C.B. Merkel cell polyomavirus and two previously unknown polyomaviruses are chronically shed from human skin. Cell Host Microbe 2010, 7, 509–515. [Google Scholar] [CrossRef] [Green Version]
- Scuda, N.; Hofmann, J.; Calvignac-Spencer, S.; Ruprecht, K.; Liman, P.; Kühn, J.; Hengel, H.; Ehlers, B. A novel human polyomavirus closely related to the african green monkey-derived lymphotropic polyomavirus. J. Virol. 2011, 85, 4586–4590. [Google Scholar] [CrossRef] [Green Version]
- Siebrasse, E.A.; Reyes, A.; Lim, E.S.; Zhao, G.; Mkakosya, R.S.; Manary, M.J.; Gordon, J.I.; Wang, D. Identification of MW polyomavirus, a novel polyomavirus in human stool. J. Virol. 2012, 86, 10321–10326. [Google Scholar] [CrossRef] [Green Version]
- Buck, C.B.; Phan, G.Q.; Raiji, M.T.; Murphy, P.M.; McDermott, D.H.; McBride, A.A. Complete genome sequence of a tenth human polyomavirus. J. Virol. 2012, 86, 10887. [Google Scholar] [CrossRef] [Green Version]
- Yu, G.; Greninger, A.L.; Isa, P.; Phan, T.G.; Martínez, M.A.; de la Luz Sanchez, M.; Contreras, J.F.; Santos-Preciado, J.I.; Parsonnet, J.; Miller, S.; et al. Discovery of a novel polyomavirus in acute diarrheal samples from children. PLoS ONE 2012, 7, e49449. [Google Scholar] [CrossRef]
- Lim, E.S.; Reyes, A.; Antonio, M.; Saha, D.; Ikumapayi, U.N.; Adeyemi, M.; Stine, O.C.; Skelton, R.; Brennan, D.C.; Mkakosya, R.S.; et al. Discovery of STL polyomavirus, a polyomavirus of ancestral recombinant origin that encodes a unique T antigen by alternative splicing. Virology 2013, 436, 295–303. [Google Scholar] [CrossRef] [Green Version]
- Korup, S.; Rietscher, J.; Calvignac-Spencer, S.; Trusch, F.; Hofmann, J.; Moens, U.; Sauer, I.; Voigt, S.; Schmuck, R.; Ehlers, B. Identification of a novel human polyomavirus in organs of the gastrointestinal tract. PLoS ONE 2013, 8, e58021. [Google Scholar] [CrossRef] [Green Version]
- Mishra, N.; Pereira, M.; Rhodes, R.H.; An, P.; Pipas, J.M.; Jain, K.; Kapoor, A.; Briese, T.; Faust, P.L.; Lipkin, W.I. Identification of a novel polyomavirus in a pancreatic transplant recipient with retinal blindness and vasculitic myopathy. J. Infect. Dis. 2014, 210, 1595–1599. [Google Scholar] [CrossRef] [Green Version]
- Gheit, T.; Dutta, S.; Oliver, J.; Robitaille, A.; Hampras, S.; Combes, J.D.; McKay-Chopin, S.; Le Calvez-Kelm, F.; Fenske, N.; Cherpelis, B.; et al. Isolation and characterization of a novel putative human polyomavirus. Virology 2017, 506, 45–54. [Google Scholar] [CrossRef]
- Feltkamp, M.C.; Kazem, S.; van der Meijden, E.; Lauber, C.; Gorbalenya, A.E. From Stockholm to Malawi: Recent developments in studying human polyomaviruses. J. Gen. Virol. 2013, 94, 482–496. [Google Scholar] [CrossRef]
- Kamminga, S.; van der Meijden, E.; Feltkamp, M.C.W.; Zaaijer, H.L. Seroprevalence of fourteen human polyomaviruses determined in blood donors. PLoS ONE 2018, 13, e0206273. [Google Scholar] [CrossRef] [Green Version]
- Padgett, B.L.; Walker, D.L.; ZuRhein, G.M.; Eckroade, R.J.; Dessel, B.H. Cultivation of papova-like virus from human brain with progressive multifocal leucoencephalopathy. Lancet 1971, 1, 1257–1260. [Google Scholar] [CrossRef]
- Hirsch, H.H.; Knowles, W.; Dickenmann, M.; Passweg, J.; Klimkait, T.; Mihatsch, M.J.; Steiger, J. Prospective study of polyomavirus type BK replication and nephropathy in renal-transplant recipients. N. Engl. J. Med. 2002, 347, 488–496. [Google Scholar] [CrossRef]
- Arthur, R.R.; Shah, K.V.; Baust, S.J.; Santos, G.W.; Saral, R. Association of BK viruria with hemorrhagic cystitis in recipients of bone marrow transplants. N Engl. J. Med. 1986, 315, 230–234. [Google Scholar] [CrossRef]
- Nguyen, K.D.; Lee, E.E.; Yue, Y.; Stork, J.; Pock, L.; North, J.P.; Vandergriff, T.; Cockerell, C.; Hosler, G.A.; Pastrana, D.V.; et al. Human polyomavirus 6 and 7 are associated with pruritic and dyskeratotic dermatoses. J. Am. Acad. Dermatol. 2017, 76, 932–940.e933. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ho, J.; Jedrych, J.J.; Feng, H.; Natalie, A.A.; Grandinetti, L.; Mirvish, E.; Crespo, M.M.; Yadav, D.; Fasanella, K.E.; Proksell, S.; et al. Human polyomavirus 7-associated pruritic rash and viremia in transplant recipients. J. Infect. Dis. 2015, 211, 1560–1565. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.H.; Zhang, J.M.; Han, W.; Chen, H.; Chen, Y.H.; Wang, F.R.; Wang, J.Z.; Zhang, Y.Y.; Mo, X.D.; Chen, Y.; et al. Viral encephalitis after haplo-identical hematopoietic stem cell transplantation: Causative viral spectrum, characteristics, and risk factors. Eur. J. Haematol. 2017, 98, 450–458. [Google Scholar] [CrossRef]
- Bárcena-Panero, A.; Van Ghelue, M.; Khan, M.T.; Echevarría, J.E.; Fedele, G.; Moens, U. BK virus-associated infection in cerebrospinal fluid of neurological patients and mutation analysis of the complete VP1 gene in different patient groups. J. Cell Physiol. 2012, 227, 136–145. [Google Scholar] [CrossRef] [PubMed]
- Barzon, L.; Squarzon, L.; Militello, V.; Trevisan, M.; Porzionato, A.; Macchi, V.; De Caro, R.; Palù, G. WU and KI polyomaviruses in the brains of HIV-positive patients with and without progressive multifocal leukoencephalopathy. J. Infect. Dis. 2009, 200, 1755–1758. [Google Scholar] [CrossRef]
- Bialasiewicz, S.; Whiley, D.M.; Lambert, S.B.; Nissen, M.D.; Sloots, T.P. Detection of BK, JC, WU, or KI polyomaviruses in faecal, urine, blood, cerebrospinal fluid and respiratory samples. J. Clin. Virol. 2009, 45, 249–254. [Google Scholar] [CrossRef]
- Dang, X.; Bialasiewicz, S.; Nissen, M.D.; Sloots, T.P.; Koralnik, I.J.; Tan, C.S. Infrequent detection of KI, WU and MC polyomaviruses in immunosuppressed individuals with or without progressive multifocal leukoencephalopathy. PLoS ONE 2011, 6, e16736. [Google Scholar] [CrossRef]
- Rockett, R.J.; Sloots, T.P.; Bowes, S.; O’Neill, N.; Ye, S.; Robson, J.; Whiley, D.M.; Lambert, S.B.; Wang, D.; Nissen, M.D.; et al. Detection of novel polyomaviruses, TSPyV, HPyV6, HPyV7, HPyV9 and MWPyV in feces, urine, blood, respiratory swabs and cerebrospinal fluid. PLoS ONE 2013, 8, e62764. [Google Scholar] [CrossRef] [Green Version]
- Delbue, S.; Elia, F.; Signorini, L.; Bella, R.; Villani, S.; Marchioni, E.; Ferrante, P.; Phan, T.G.; Delwart, E. Human polyomavirus 6 DNA in the cerebrospinal fluid of an HIV-positive patient with leukoencephalopathy. J. Clin. Virol. 2015, 68, 24–27. [Google Scholar] [CrossRef]
- RUBIN, J.; GIRAUD, G.; PRIFTAKIS, P.; WIDE, K.; GUSTAFSSON, B.; RAMQVIST, T.; DALIANIS, T. No Detection of BK Virus, JC Virus, KI, WU and Merkel Cell Polyomaviruses in Cerebrospinal Fluid of Patients with Neurological Complications after Hematopoetic Stem Cell Transplantation. Anticancer Res. 2011, 31, 3489–3492. [Google Scholar]
- Delbue, S.; Sotgiu, G.; Fumagalli, D.; Valli, M.; Borghi, E.; Mancuso, R.; Marchioni, E.; Maserati, R.; Ferrante, P. A case of a progressive multifocal leukoencephalopathy patient with four different JC virus transcriptional control region rearrangements in cerebrospinal fluid, blood, serum, and urine. J. Neurovirol. 2005, 11, 51–57. [Google Scholar] [CrossRef] [PubMed]
- Tremolada, S.; Delbue, S.; Larocca, S.; Carloni, C.; Elia, F.; Khalili, K.; Gordon, J.; Ferrante, P. Polymorphisms of the BK virus subtypes and their influence on viral in vitro growth efficiency. Virus Res. 2010, 149, 190–196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goh, S.; Lindau, C.; Tiveljung-Lindell, A.; Allander, T. Merkel cell polyomavirus in respiratory tract secretions. Emerg. Infect. Dis 2009, 15, 489–491. [Google Scholar] [CrossRef] [PubMed]
- Bella, R.; Dolci, M.; Ferraresso, M.; Ticozzi, R.; Ghio, L.; Rizzo, J.; Signorini, L.; Villani, S.; Elia, F.; Ferrante, P.; et al. Human herpesvirus-6 and polyomaviruses DNAemia in children and young adult patients after kidney transplantation. Future Virol. 2015, 10, 1275–1284. [Google Scholar] [CrossRef]
- Lo, Y.M.; Tein, M.S.; Lau, T.K.; Haines, C.J.; Leung, T.N.; Poon, P.M.; Wainscoat, J.S.; Johnson, P.J.; Chang, A.M.; Hjelm, N.M. Quantitative analysis of fetal DNA in maternal plasma and serum: Implications for noninvasive prenatal diagnosis. Am. J. Hum. Genet. 1998, 62, 768–775. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Delbue, S.; Ferraresso, M.; Elia, F.; Belingheri, M.; Carloni, C.; Signorini, L.; Carluccio, S.; Dallari, S.; Ghio, L.; Ferrante, P. Investigation of polyomaviruses replication in pediatric patients with nephropathy receiving rituximab. J. Med. Virol. 2012, 84, 1464–1470. [Google Scholar] [CrossRef] [PubMed]
- Hashida, Y.; Higuchi, T.; Matsui, K.; Shibata, Y.; Nakajima, K.; Sano, S.; Daibata, M. Genetic Variability of the Noncoding Control Region of Cutaneous Merkel Cell Polyomavirus: Identification of Geographically Related Genotypes. J. Infect. Dis. 2018, 217, 1601–1611. [Google Scholar] [CrossRef] [Green Version]
- Agostini, H.T.; Ryschkewitsch, C.F.; Stoner, G.L. Genotype profile of human polyomavirus JC excreted in urine of immunocompetent individuals. J. Clin. Microbiol. 1996, 34, 159–164. [Google Scholar]
- Ault, G.S.; Stoner, G.L. Human polyomavirus JC promoter/enhancer rearrangement patterns from progressive multifocal leukoencephalopathy brain are unique derivatives of a single archetypal structure. J. Gen. Virol. 1993, 74, 1499–1507. [Google Scholar] [CrossRef]
- Jensen, P.N.; Major, E.O. A classification scheme for human polyomavirus JCV variants based on the nucleotide sequence of the noncoding regulatory region. J. Neurovirol. 2001, 7, 280–287. [Google Scholar] [CrossRef]
- Jin, L.; Gibson, P.E.; Knowles, W.A.; Clewley, J.P. BK virus antigenic variants: Sequence analysis within the capsid VP1 epitope. J. Med. Virol. 1993, 39, 50–56. [Google Scholar] [CrossRef] [PubMed]
- Siebrasse, E.A.; Bauer, I.; Holtz, L.R.; Le, B.M.; Lassa-Claxton, S.; Canter, C.; Hmiel, P.; Shenoy, S.; Sweet, S.; Turmelle, Y.; et al. Human polyomaviruses in children undergoing transplantation, United States, 2008–2010. Emerg. Infect. Dis. 2012, 18, 1676–1679. [Google Scholar] [CrossRef] [PubMed]
- Chittick, P.; Williamson, J.C.; Ohl, C.A. BK virus encephalitis: Case report, review of the literature, and description of a novel treatment modality. Ann. Pharmacother. 2013, 47, 1229–1233. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.; Kim, Y.J.; Cho, H. BK virus nephropathy and multiorgan involvement in a child with heart transplantation. Clin. Nephrol. 2019, 91, 107–113. [Google Scholar] [CrossRef]
- Jun, J.B.; Choi, Y.; Kim, H.; Lee, S.H.; Jeong, J.; Jung, J. BK polyomavirus encephalitis in a patient with thrombotic microangiopathy after an allogeneic hematopoietic stem cell transplant. Transpl. Infect. Dis. 2016, 18, 950–953. [Google Scholar] [CrossRef] [PubMed]
- Elsner, C.; Dörries, K. Evidence of human polyomavirus BK and JC infection in normal brain tissue. Virology 1992, 191, 72–80. [Google Scholar] [CrossRef]
- Vago, L.; Cinque, P.; Sala, E.; Nebuloni, M.; Caldarelli, R.; Racca, S.; Ferrante, P.; Trabottoni, G.; Costanzi, G. JCV-DNA and BKV-DNA in the CNS tissue and CSF of AIDS patients and normal subjects. Study of 41 cases and review of the literature. J. Acquir. Immune Defic. Syndr. Hum. Retrovirol. 1996, 12, 139–146. [Google Scholar] [CrossRef]
- Barzon, L.; Squarzon, L.; Pacenti, M.; Scotton, P.G.; Palù, G. Detection of WU polyomavirus in cerebrospinal fluid specimen from a patient with AIDS and suspected progressive multifocal leukoencephalopathy. J. Infect. Dis. 2009, 200, 314–315. [Google Scholar] [CrossRef]
- Sadeghi, F.; Salehi-Vaziri, M.; Alizadeh, A.; Ghodsi, S.M.; Bokharaei-Salim, F.; Fateh, A.; Monavari, S.H.; Keyvani, H. Detection of Merkel cell polyomavirus large T-antigen sequences in human central nervous system tumors. J. Med. Virol. 2015, 87, 1241–1247. [Google Scholar] [CrossRef]
- McDonald, C.P.; Roy, A.; Mahajan, P.; Smith, R.; Charlett, A.; Barbara, J.A. Relative values of the interventions of diversion and improved donor-arm disinfection to reduce the bacterial risk from blood transfusion. Vox Sang. 2004, 86, 178–182. [Google Scholar] [CrossRef]
- Bruneau, C.; Perez, P.; Chassaigne, M.; Allouch, P.; Audurier, A.; Gulian, C.; Janus, G.; Boulard, G.; De Micco, P.; Salmi, L.R.; et al. Efficacy of a new collection procedure for preventing bacterial contamination of whole-blood donations. Transfusion 2001, 41, 74–81. [Google Scholar] [CrossRef] [PubMed]
- Del Valle, L.; Piña-Oviedo, S. Human Polyomavirus JCPyV and Its Role in Progressive Multifocal Leukoencephalopathy and Oncogenesis. Front. Oncol. 2019, 9, 711. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
| Age in years, mean (range) | 58 (17–92) |
| Female, number (%) | 117 (50.0%) |
| Suspected diagnosis at hospital admission | Meningitis/encephalitis, 139 (59.4%) Peripheral neuropathy, 30 (13%) Cognitive disorders, 19 (8.1%) Multiple sclerosis, 10 (4.2%) Brain tumor/metastasis, 9 (3.8%) Epilepsy, 5 (2.1%) Confused state, 5 (2.1%) Hydrocephalus, 4 (1.7%) Migraine, 4 (1.7%) Progressive multifocal leukoencephalopathy, 3 (1.3%) Mood disorder, 3 (1.3%) Myelitis, 3 (1.3%) Sjogren syndrome, 1 (0.4%) Guillain–Barré syndrome, 1 (0.4%) Myeloproliferative disease, 1 (0.4%) Angioma, 1 (0.4%) |
| HPyVs Prevalence | |||||
|---|---|---|---|---|---|
| No. of Cases | JCPyV+/tot (%) | BKPyV+/tot (%) | MCPyV+/tot (%) | HPyV-6+/tot (%) | HPyV+/tot (%) |
| 234 | 3/234 * (1.3%) | 15/234 *° (6.4%) | 22/234 *° (9.4%) | 1/234 * (0.4) | 41/234 (17.5) |
| JCPyV+ | BKPyV+ | MCPyV+ | |
|---|---|---|---|
| HSV-1+ | / | 1 | 1 |
| EBV+ | 2 | 2 | / |
| Enteroviruses+ | / | 1 | 3 |
| HIV+ | 2 | / | / |
| HPyVs (no. + Cases) | Disease (Frequency; Percentage) |
|---|---|
| JCPyV + (3) | PML (3/3; 100%) |
| BKPyV + (15) | Meningoencephalitis (9/135; 6.7%) Peripheral neuropathy (3/30; 10%) Cognitive disorder (1/19; 5.3%) Sjogren syndrome (1/1; 100%) Mood disorder (1/3; 33%) |
| MCPyV+ (22) | Meningoencephalitis (15/135; 11.1%) Neuropathy (2/30; 6.7%) Migraine (1/4; 25%) Multiple sclerosis (1/10; 10%) Brain tumor (1/9; 11.1%) Myeloproliferative disease (1/1; 100%) Angioma (1/1; 100%) |
| HPyV6+ (1) | Meningoencephalitis (1/135; 7.4%) |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Delbue, S.; Franciotta, D.; Giannella, S.; Dolci, M.; Signorini, L.; Ticozzi, R.; D’Alessandro, S.; Campisciano, G.; Comar, M.; Ferrante, P.; et al. Human Polyomaviruses in the Cerebrospinal Fluid of Neurological Patients. Microorganisms 2020, 8, 16. https://doi.org/10.3390/microorganisms8010016
Delbue S, Franciotta D, Giannella S, Dolci M, Signorini L, Ticozzi R, D’Alessandro S, Campisciano G, Comar M, Ferrante P, et al. Human Polyomaviruses in the Cerebrospinal Fluid of Neurological Patients. Microorganisms. 2020; 8(1):16. https://doi.org/10.3390/microorganisms8010016
Chicago/Turabian StyleDelbue, Serena, Diego Franciotta, Sara Giannella, Maria Dolci, Lucia Signorini, Rosalia Ticozzi, Sarah D’Alessandro, Giuseppina Campisciano, Manola Comar, Pasquale Ferrante, and et al. 2020. "Human Polyomaviruses in the Cerebrospinal Fluid of Neurological Patients" Microorganisms 8, no. 1: 16. https://doi.org/10.3390/microorganisms8010016
APA StyleDelbue, S., Franciotta, D., Giannella, S., Dolci, M., Signorini, L., Ticozzi, R., D’Alessandro, S., Campisciano, G., Comar, M., Ferrante, P., & Ciotti, M. (2020). Human Polyomaviruses in the Cerebrospinal Fluid of Neurological Patients. Microorganisms, 8(1), 16. https://doi.org/10.3390/microorganisms8010016