American Crows as Carriers of Extra Intestinal Pathogenic E. coli and Avian Pathogenic-Like E. coli and Their Potential Impact on a Constructed Wetland

 , ,
, ,
Abstract
:1. Introduction
2. Material and Methods
2.1. Sample Collections
2.2. Isolation of E. coli
2.3. Antibiotic Susceptibility Testing
2.4. DNA Isolation and PCR
2.5. Virulence Gene Detection
2.6. Detection of Antibiotic Resistance Genes and Plasmid Incompatibility (INC) Sites
2.7. Mating Experiments
2.8. Phylogenetic Studies
2.9. Statistical Analysis
3. Results
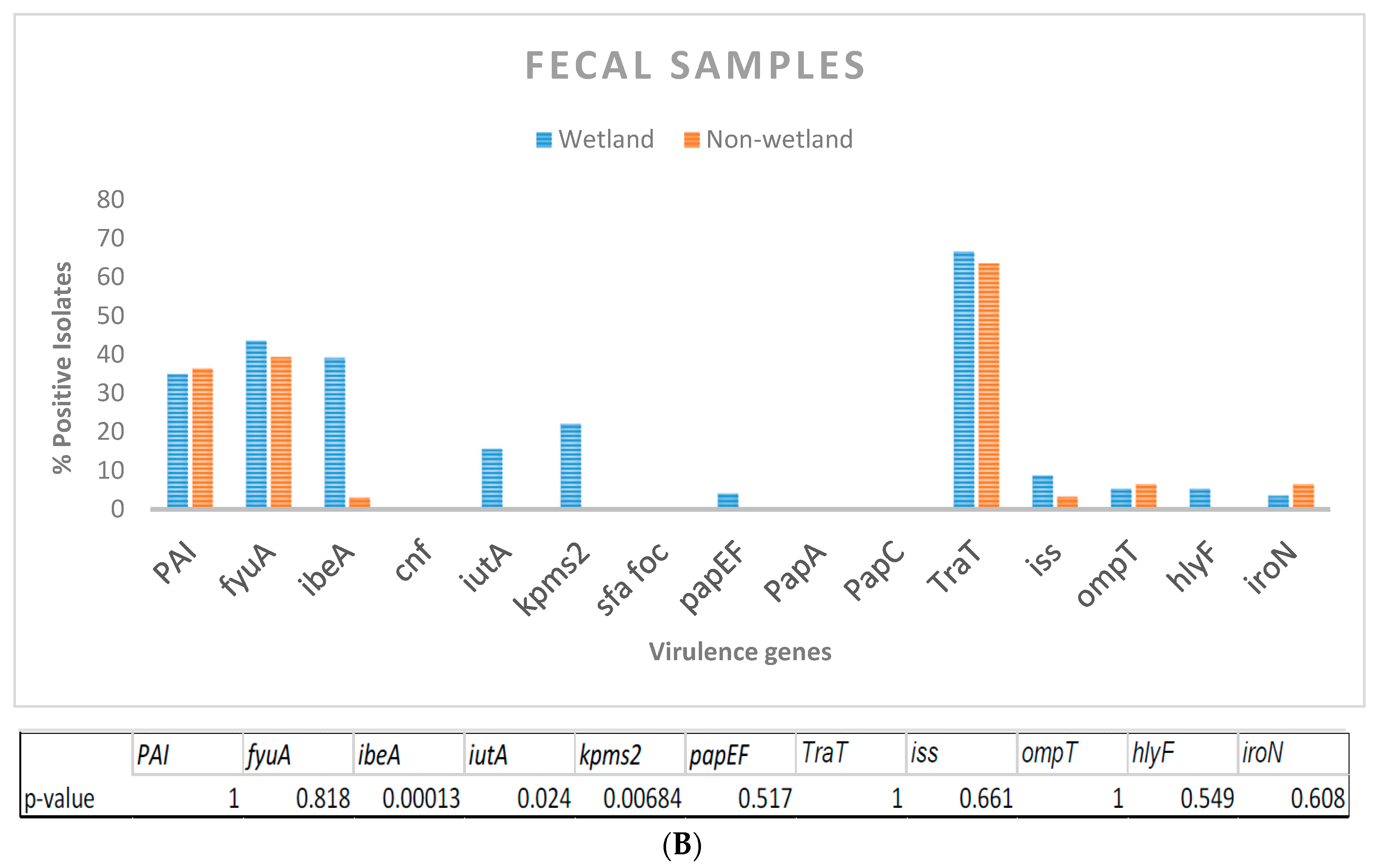
3.1. Relative Abundance of Virulence Genes in Crow Feces and Water Isolates
3.2. Identification of EXPEC, IPEC, STEC Strains
3.3. Identification of APEC Strains and Phylogenetic Types of the Isolates
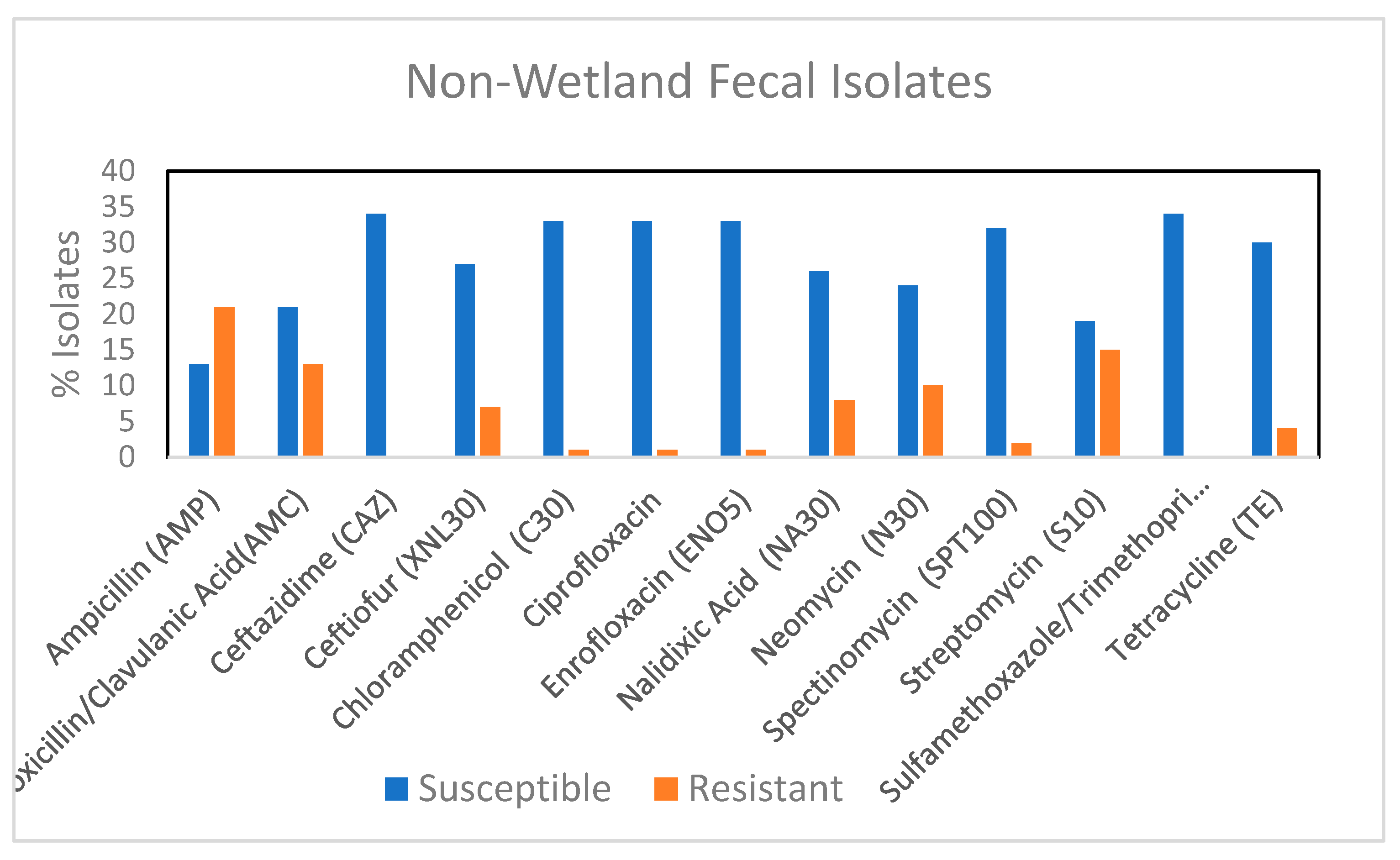
3.4. Comparison of Antibiotic Resistance of Fecal Isolates from Wetland and Non-Wetland Areas
3.5. Replicon Typing and Transferability of Plasmid
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Reed, K.D.; Meece, J.K.; Henkel, J.S.; Shukla, S.K. Birds, migration and emerging zoonoses: West nile virus, lyme disease, influenza A and enteropathogens. Clin. Med. Res. 2003, 1, 5–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benskin, C.M.; Wilson, K.; Jones, K.; Hartley, I.R. Bacterial pathogens in wild birds: A review of the frequency and effects of infection. Biol. Rev. Camb. Philos. Soc. 2009, 84, 349–373. [Google Scholar] [CrossRef] [PubMed]
- al-Sallami, S. A possible role of crows in the spread of diarrhoeal diseases in Aden. J. Egypt. Public Health Assoc. 1991, 66, 441–449. [Google Scholar] [PubMed]
- Ejidokun, O.O.; Walsh, A.; Barnett, J.; Hope, Y.; Ellis, S.; Sharp, M.W.; Paiba, G.A.; Logan, M.; Willshaw, G.A.; Cheasty, T. Human Vero cytotoxigenic Escherichia coli (VTEC) O157 infection linked to birds. Epidemiol. Infect. 2006, 134, 421–423. [Google Scholar] [CrossRef] [PubMed]
- Hinton, M.G.; Reisen, W.K.; Wheeler, S.S.; Townsend, A.K. West nile virus activity in a winter roost of American crows (Corvus brachyrhynchos): Is bird-to-bird transmission important in persistence and amplification? J. Med. Entomol. 2015, 52, 683–692. [Google Scholar] [CrossRef] [Green Version]
- Soderlund, R.; Skarin, H.; Borjesson, S.; Sanno, A.; Jernberg, T.; Aspan, A.; Agren, E.O.; Hansson, I. Prevalence and genomic characteristics of zoonotic gastro-intestinal pathogens and ESBL/pAmpC producing Enterobacteriaceae among Swedish corvid birds. Infect. Ecol. Epidemiol. 2019, 9, 1701399. [Google Scholar] [CrossRef] [Green Version]
- Croxen, M.A.; Law, R.J.; Scholz, R.; Keeney, K.M.; Wlodarska, M.; Finlay, B.B. Recent advances in understanding enteric pathogenic Escherichia coli. Clin. Microbiol. Rev. 2013, 26, 822–880. [Google Scholar] [CrossRef] [Green Version]
- van Elsas, J.D.; Semenov, A.V.; Costa, R.; Trevors, J.T. Survival of Escherichia coli in the environment: Fundamental and public health aspects. ISME J. 2011, 5, 173–183. [Google Scholar] [CrossRef] [Green Version]
- Mokady, D.; Gophna, U.; Ron, E.Z. Extensive Gene Diversity in Septicemic Escherichia coli Strains. J. Clin. Microbiol. 2005, 43, 66–73. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez-Siek, K.E.; Giddings, C.W.; Doetkott, C.; Johnson, T.J.; Nolan, L.K. Characterizing the APEC pathotype. Vet. Res. 2005, 36, 241–256. [Google Scholar] [CrossRef] [Green Version]
- Johnson, T.J.; Wannemuehler, Y.; Johnson, S.J.; Stell, A.L.; Doetkott, C.; Johnson, J.R.; Kim, K.S.; Spanjaard, L.; Nolan, L.K. Comparison of extraintestinal pathogenic Escherichia coli strains from human and avian sources reveals a mixed subset representing potential zoonotic pathogens. Appl. Environ. Microbiol. 2008, 74, 7043–7050. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tivendale, K.A.; Logue, C.M.; Kariyawasam, S.; Jordan, D.; Hussein, A.; Li, G.; Wannemuehler, Y.; Nolan, L.K. Avian-pathogenic Escherichia coli strains are similar to neonatal meningitis E. coli strains and are able to cause meningitis in the rat model of human disease. Infect. Immun. 2010, 78, 3412–3419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, T.J.; Kariyawasam, S.; Wannemuehler, Y.; Mangiamele, P.; Johnson, S.J.; Doetkott, C.; Skyberg, J.A.; Lynne, A.M.; Johnson, J.R.; Nolan, L.K. The genome sequence of avian pathogenic Escherichia coli strain O1:K1:H7 shares strong similarities with human extraintestinal pathogenic E. coli genomes. J. Bacteriol. 2007, 189, 3228–3236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, T.J.; Logue, C.M.; Wannemuehler, Y.; Kariyawasam, S.; Doetkott, C.; DebRoy, C.; White, D.G.; Nolan, L.K. Examination of the source and extended virulence genotypes of Escherichia coli contaminating retail poultry meat. Foodborne Pathog. Dis. 2009, 6, 657–667. [Google Scholar] [CrossRef] [Green Version]
- Zhu Ge, X.; Jiang, J.; Pan, Z.; Hu, L.; Wang, S.; Wang, H.; Leung, F.C.; Dai, J.; Fan, H. Comparative genomic analysis shows that avian pathogenic Escherichia coli isolate IMT5155 (O2:K1:H5; ST complex 95, ST140) shares close relationship with ST95 APEC O1:K1 and human ExPEC O18:K1 strains. PLoS ONE 2014, 9, e112048. [Google Scholar] [CrossRef] [Green Version]
- Johnson, J.R.; Murray, A.C.; Gajewski, A.; Sullivan, M.; Snippes, P.; Kuskowski, M.A.; Smith, K.E. Isolation and molecular characterization of nalidixic acid-resistant extraintestinal pathogenic Escherichia coli from retail chicken products. Antimicrob. Agents Chemother. 2003, 47, 2161–2168. [Google Scholar] [CrossRef] [Green Version]
- Johnson, T.J.; Wannemuehler, Y.; Doetkott, C.; Johnson, S.J.; Rosenberger, S.C.; Nolan, L.K. Identification of minimal predictors of avian pathogenic Escherichia coli virulence for use as a rapid diagnostic tool. J. Clin. Microbiol. 2008, 46, 3987–3996. [Google Scholar] [CrossRef] [Green Version]
- Johnson, T.J.; Jordan, D.; Kariyawasam, S.; Stell, A.L.; Bell, N.P.; Wannemuehler, Y.M.; Alarcon, C.F.; Li, G.; Tivendale, K.A.; Logue, C.M.; et al. Sequence analysis and characterization of a transferable hybrid plasmid encoding multidrug resistance and enabling zoonotic potential for extraintestinal Escherichia coli. Infect. Immun. 2010, 78, 1931–1942. [Google Scholar] [CrossRef] [Green Version]
- Sen, K.; Berglund, T.; Soares, M.A.; Taheri, B.; Ma, Y.; Khalil, L.; Fridge, M.; Lu, J.; Turner, R.J. Antibiotic resistance of E. coli isolated from a constructed wetland dominated by a crow roost, with emphasis on ESBL and AmpC containing E. coli. Front. Microbiol. 2019, 10, 1034. [Google Scholar] [CrossRef] [Green Version]
- Pitout, J.D. Extraintestinal pathogenic Escherichia coli: A combination of virulence with antibiotic resistance. Front. Microbiol. 2012, 3, 9. [Google Scholar] [CrossRef] [Green Version]
- Sen, K.; J, L.S.; Boczek, L.; Rice, E.W. Development of a sensitive detection method for stressed E. coli O157:H7 in source and finished drinking water by culture-qPCR. Environ. Sci. Technol. 2011, 45, 2250–2256. [Google Scholar] [CrossRef] [PubMed]
- Johnson, J.R.; Stell, A.L. Extended virulence genotypes of Escherichia coli strains from patients with urosepsis in relation to phylogeny and host compromise. J. Infect. Dis. 2000, 181, 261–272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, T.J.; Wannemuehler, Y.M.; Johnson, S.J.; Logue, C.M.; White, D.G.; Doetkott, C.; Nolan, L.K. Plasmid replicon typing of commensal and pathogenic Escherichia coli isolates. Appl. Environ. Microbiol. 2007, 73, 1976–1983. [Google Scholar] [CrossRef] [Green Version]
- Carattoli, A.; Bertini, A.; Villa, L.; Falbo, V.; Hopkins, K.L.; Threlfall, E.J. Identification of plasmids by PCR-based replicon typing. J. Microbiol. Methods 2005, 63, 219–228. [Google Scholar] [CrossRef] [PubMed]
- Lyimo, B.; Buza, J.; Subbiah, M.; Temba, S.; Kipasika, H.; Smith, W.; Call, D.R. IncF plasmids are commonly carried by antibiotic resistant Escherichia coli isolated from drinking water sources in Northern Tanzania. Int. J. Microbiol. 2016, 2016, 3103672. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clermont, O.; Christenson, J.K.; Denamur, E.; Gordon, D.M. The Clermont Escherichia coli phylo-typing method revisited: Improvement of specificity and detection of new phylo-groups. Environ. Microbiol. Rep. 2013, 5, 58–65. [Google Scholar] [CrossRef] [PubMed]
- Ewers, C.; Antao, E.M.; Diehl, I.; Philipp, H.C.; Wieler, L.H. Intestine and environment of the chicken as reservoirs for extraintestinal pathogenic Escherichia coli strains with zoonotic potential. Appl. Environ. Microbiol. 2009, 75, 184–192. [Google Scholar] [CrossRef] [Green Version]
- Najafi, S.; Rahimi, M.; Nikousefat, Z. Extra-intestinal pathogenic Escherichia coli from human and avian origin: Detection of the most common virulence-encoding genes. Vet. Res. Forum Int. Q. J. 2019, 10, 43–49. [Google Scholar]
- Bauchart, P.; Germon, P.; Brée, A.; Oswald, E.; Hacker, J.; Dobrindt, U. Pathogenomic comparison of human extraintestinal and avian pathogenic Escherichia coli—Search for factors involved in host specificity or zoonotic potential. Microb. Pathog. 2010, 49, 105–115. [Google Scholar] [CrossRef]
- Vandekerchove, D.; Vandemaele, F.; Adriaensen, C.; Zaleska, M.; Hernalsteens, J.P.; De Baets, L.; Butaye, P.; Van Immerseel, F.; Wattiau, P.; Laevens, H.; et al. Virulence-associated traits in avian Escherichia coli: Comparison between isolates from colibacillosis-affected and clinically healthy layer flocks. Vet. Microbiol. 2005, 108, 75–87. [Google Scholar] [CrossRef]
- Wang, S.; Niu, C.; Shi, Z.; Xia, Y.; Yaqoob, M.; Dai, J.; Lu, C. Effects of ibeA deletion on virulence and biofilm formation of avian pathogenic Escherichia coli. Infect. Immun. 2011, 79, 279–287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bergholz, P.W.; Noar, J.D.; Buckley, D.H. Environmental patterns are imposed on the population structure of Escherichia coli after fecal deposition. Appl. Environ. Microbiol. 2011, 77, 211–219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, T.J.; Logue, C.M.; Johnson, J.R.; Kuskowski, M.A.; Sherwood, J.S.; Barnes, H.J.; DebRoy, C.; Wannemuehler, Y.M.; Obata-Yasuoka, M.; Spanjaard, L.; et al. Associations between multidrug resistance, plasmid content, and virulence potential among extraintestinal pathogenic and commensal Escherichia coli from humans and poultry. Foodborne Pathog. Dis. 2012, 9, 37–46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hanson, N.D.; Moland, E.S.; Hong, S.G.; Propst, K.; Novak, D.J.; Cavalieri, S.J. Surveillance of community-based reservoirs reveals the presence of CTX-M, imported AmpC, and OXA-30 beta-lactamases in urine isolates of Klebsiella pneumoniae and Escherichia coli in a U.S. community. Antimicrob. Agents Chemother. 2008, 52, 3814–3816. [Google Scholar] [CrossRef] [Green Version]
- Qin, X.; Zerr, D.M.; Weissman, S.J.; Englund, J.A.; Denno, D.M.; Klein, E.J.; Tarr, P.I.; Kwong, J.; Stapp, J.R.; Tulloch, L.G.; et al. Prevalence and mechanisms of broad-spectrum beta-lactam resistance in Enterobacteriaceae: A children’s hospital experience. Antimicrob. Agents Chemother. 2008, 52, 3909–3914. [Google Scholar] [CrossRef] [Green Version]
- Johnson, T.J.; Giddings, C.W.; Horne, S.M.; Gibbs, P.S.; Wooley, R.E.; Skyberg, J.; Olah, P.; Kercher, R.; Sherwood, J.S.; Foley, S.L.; et al. Location of increased serum survival gene and selected virulence traits on a conjugative R plasmid in an avian Escherichia coli isolate. Avian Dis. 2002, 46, 342–352. [Google Scholar] [CrossRef]
- Fricke, W.F.; McDermott, P.F.; Mammel, M.K.; Zhao, S.; Johnson, T.J.; Rasko, D.A.; Fedorka-Cray, P.J.; Pedroso, A.; Whichard, J.M.; Leclerc, J.E.; et al. Antimicrobial resistance-conferring plasmids with similarity to virulence plasmids from avian pathogenic Escherichia coli strains in Salmonella enterica serovar Kentucky isolates from poultry. Appl. Environ. Microbiol. 2009, 75, 5963–5971. [Google Scholar] [CrossRef] [Green Version]
- Skyberg, J.A.; Johnson, T.J.; Johnson, J.R.; Clabots, C.; Logue, C.M.; Nolan, L.K. Acquisition of avian pathogenic Escherichia coli plasmids by a commensal E. coli isolate enhances its abilities to kill chicken embryos, grow in human urine, and colonize the murine kidney. Infect. Immun. 2006, 74, 6287–6292. [Google Scholar] [CrossRef] [Green Version]
- Le Gall, T.; Clermont, O.; Gouriou, S.; Picard, B.; Nassif, X.; Denamur, E.; Tenaillon, O. Extraintestinal virulence is a coincidental by-product of commensalism in B2 phylogenetic group Escherichia coli strains. Mol. Biol. Evol. 2007, 24, 2373–2384. [Google Scholar] [CrossRef] [Green Version]
- Picard, B.; Garcia, J.S.; Gouriou, S.; Duriez, P.; Brahimi, N.; Bingen, E.; Elion, J.; Denamur, E. The link between phylogeny and virulence in Escherichia coli extraintestinal infection. Infect. Immun. 1999, 67, 546–553. [Google Scholar] [CrossRef] [Green Version]
- Schierack, P.; Walk, N.; Ewers, C.; Wilking, H.; Steinruck, H.; Filter, M.; Wieler, L.H. ExPEC-typical virulence-associated genes correlate with successful colonization by intestinal E. coli in a small piglet group. Environ. Microbiol. 2008, 10, 1742–1751. [Google Scholar] [CrossRef] [PubMed]
- Dissanayake, D.R.A.; Octavia, S.; Lan, R. Population structure and virulence content of avian pathogenic Escherichia coli isolated from outbreaks in Sri Lanka. Vet. Microbiol. 2014, 168, 403–412. [Google Scholar] [CrossRef] [PubMed]
- Jamborova, I.; Dolejska, M.; Zurek, L.; Townsend, A.K.; Clark, A.B.; Ellis, J.C.; Papousek, I.; Cizek, A.; Literak, I. Plasmid-mediated resistance to cephalosporins and quinolones in Escherichia coli from American crows in the USA. Environ. Microbiol. 2017, 19, 2025–2036. [Google Scholar] [CrossRef] [PubMed]
- Verbeek, N.A.M. American Crow (Corvus brachyrhynchos). The Birds of North America; 2002 edition; Rodewald, P.G., Ed.; Cornell Lab of Orinthology: Ithaca, NY, USA, 2002. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | PAI | fyuA | ibeA | cnf | iutA | kpmsT II/KPs1 | Sfa/foc | papEF | papA | papC | traT | iss | ompT | hlyF | iroN | chuA | cva | Potential Pathotype | Phylotype | Replicon Type |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| FECAL | ||||||||||||||||||||
| F11.1 (R2) | 1 | 1 | 0 | 0 | 1 | 1 | 0 | 1 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 1 | 0 | ExPEC | B2 | FIB |
| F11.2 (R2) | 1 | 1 | 0 | 0 | 1 | 1 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 1 | 0 | ExPEC | B2 | FIB |
| F13.1 (R2) | 1 | 1 | 0 | 0 | 1 | 1 | 0 | 1 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 1 | 0 | ExPEC | B2 | FIB |
| F13.2 (R2) | 1 | 1 | 0 | 0 | 1 | 1 | 0 | 1 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 1 | 0 | ExPEC | B2 | FIB |
| F15.2 (R2) | 1 | 1 | 1 | 0 | 1 | 1 | 0 | 0 | 0 | 0 | 1 | 1 | 0 | 0 | 0 | 1 | 0 | ExPEC | B2 | FIB |
| F16.2 (R2) | 1 | 1 | 1 | 0 | 1 | 1 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 1 | 0 | ExPEC | B2 | FIB |
| F39.2 (R3) | 0 | 0 | 1 | 0 | 1 | 1 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | ExPEC | B1 | FIC |
| F42.2 (R4) | 1 | 1 | 1 | 0 | 1 | 1 | 0 | 0 | 0 | 0 | 1 | 0 | 1 | 0 | 0 | 0 | 0 | ExPEC | B1 | B/O |
| F43.1 (R4) | 0 | 1 | 1 | 0 | 1 | 0 | 1 | 0 | 0 | 1 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | ExPEC | A | B/O, A/C |
| F46.2 (R4) | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 0 | 0 | 0 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | ExPEC/APEC | B2 | FIB |
| F47.1 (R4) | 0 | 0 | 0 | 0 | 1 | 1 | 0 | 0 | 0 | 0 | 1 | 1 | 0 | 0 | 1 | 0 | 0 | APEC | C | I1, FIB |
| F53.1 (R5) | 0 | 0 | 0 | 0 | 1 | 1 | 0 | 0 | 0 | 0 | 1 | 1 | 0 | 1 | 1 | 0 | 0 | APEC | B1 | I1 |
| FS 7 (R6) | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | APEC | cryptic clade | FIB |
| FS1 (R6) | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 0 | 0 | 1 | 1 | 1 | APEC | F | FIB |
| FS4 (R6) | 0 | 0 | 1 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 1 | 1 | 1 | APEC | F | FIB |
| WATER | ||||||||||||||||||||
| NC5.1ctx (R2) | 1 | 1 | 0 | 0 | 1 | 1 | 0 | 1 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 1 | 0 | ExPEC | B2 | FIB, Frep |
| NC6.4ctx (R2) | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 1 | 1 | 1 | 0 | 1 | APEC | B1 | FIB |
| SW2.1 (R3) | 0 | 0 | 1 | 0 | 1 | 0 | 0 | 1 | 1 | 1 | 0 | 1 | 0 | 1 | 1 | 1 | 1 | ExPEC/APEC | B2 | FIB, I1 |
| RP3.2 (R4) | 0 | 0 | 1 | 0 | 0 | 1 | 1 | 1 | 1 | 0 | 0 | 1 | 1 | 0 | 1 | 0 | 0 | ExPEC/APEC | B1 | B/0; I1 |
| SW2.26/22/16 (No Rain) | 0 | 1 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 1 | 1 | 0 | 1 | 1 | APEC | B2 | I1, FIB, FIA |
| RS1.1 6/22/16 (No Rain) | 1 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | ExPEC | B2 | FIB |
| RS1.3 6/22/16 (No Rain) | 1 | 0 | 0 | 1 | 1 | 1 | 0 | 0 | 1 | 0 | 1 | 0 | 1 | 1 | 1 | 1 | 0 | EXPEC/APEC | B2 | FIB |
| RS2.2 6/22/16 (No Rain) | 1 | 1 | 1 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 1 | 0 | 1 | 1 | APEC | B2 | FIB |
| NC6.1 6/22/16 (No Rain) | 0 | 1 | 1 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 1 | 1 | 0 | 1 | 1 | APEC | B2 | FIB, FIA |
| NC6.2 6/22/16 (No Rain) | 1 | 1 | 1 | 0 | 1 | 1 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 1 | 0 | 1 | 1 | ExPEC/APEC | B2 | FIB, FIA |
| NC1.1 7/9/16 (No Rain) | 1 | 1 | 1 | 0 | 0 | 1 | 1 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 0 | ExPEC | B2 | No ID |
| NC1.2 10/3/16 (No rain) | 1 | 1 | 0 | 1 | 0 | 1 | 0 | 0 | 0 | 0 | 1 | 0 | 1 | 0 | 1 | 1 | 0 | APEC | B2 | FIB |
| NC1.3 10/3/16 (No rain) | 1 | 1 | 1 | 0 | 1 | 0 | 1 | 1 | 1 | 1 | 1 | 0 | 0 | 0 | 0 | 1 | 0 | ExPEC | B2 | FIB |
| RS2.3 10/3/16 (No Rain) | 1 | 1 | 1 | 0 | 1 | 0 | 1 | 1 | 1 | 1 | 1 | 0 | 0 | 0 | 0 | 1 | 0 | ExPEC | B2 | FIB |
| SW8.1 10/4/16 (Rain) | 0 | 1 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 0 | 0 | 1 | 0 | 0 | APEC | B1 | FIB |
| RS1.1 1/31/17 (Rain) | 1 | 1 | 0 | 0 | 1 | 0 | 0 | 1 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | ExPEC | A | I1 |
| NC6.1 1/31/17 (Rain) | 0 | 1 | 1 | 0 | 1 | 0 | 1 | 1 | 0 | 0 | 0 | 1 | 1 | 1 | 0 | 1 | 0 | ExPEC/APEC | B2 | FIB, FIA |
| RS2.2 1/31/17 (Rain) | 1 | 1 | 0 | 0 | 1 | 0 | 0 | 1 | 0 | 0 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | APEC | B2 | FIB |
| NC1.2 1/31/17 (Rain) | 1 | 1 | 1 | 0 | 0 | 1 | 1 | 1 | 0 | 0 | 1 | ND | ND | ND | ND | 1 | 0 | ExPEC | B2 | No ID |
| RS2.3 1/31/17 (Rain) | 1 | 1 | 0 | 0 | 1 | 0 | 0 | 1 | 0 | 0 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | ExPEC/APEC | B2 | FIB |
| NC1.3 1/31/17 (Rain) | 1 | 1 | 0 | 0 | 0 | 0 | 1 | 1 | 0 | 0 | 1 | 1 | ND | ND | ND | 0 | - | ExPEC | B1 | FIB |
| SW2.21/31/17 (Rain) | 1 | 1 | 1 | 1 | 0 | 0 | 0 | 0 | 1 | 1 | 1 | 0 | 1 | 1 | 0 | 1 | 0 | ExPEC/APEC | B2 | FIB |
| Replicon Type | Number of Isolates | % |
|---|---|---|
| FIB | 35 | 59.32 |
| I1 | 15 | 25.42 |
| FIC | 3 | 5.08 |
| F1A | 8 | 13.56 |
| P | 1 | 1.69 |
| B/0 | 7 | 11.86 |
| FIIA | 1 | 1.69 |
| FIB, I1, F1C | 1 | 1.69 |
| F1A, FIB | 5 | 8.47 |
| F11A, FIB, B/O | 1 | 1.69 |
| I1, FIB, IA | 1 | 1.69 |
| FIB, B/O | 2 | 3.39 |
| A/C | 1 | 1.69 |
| P, FIC | 1 | 1.69 |
| Strain ID | Resistance Phenotype * | AR Genes * | Mating | Replicon Type | Trans Conjugant |
|---|---|---|---|---|---|
| Fecal | Donor | Donor | |||
| F11.1 | AMP-CTX-CF-XNL-S-SXT-T | blactx, strA, sul1, tet(A), tet(M) | 1 | FIB | blactx-M, strB |
| F11.2 | AMP-CAZ-CF-XNL-S-SXT-T | blactx, strA, strB, sul1, tet(A), tet(B), tet(M) | 1 | FIB | bla ctx-M, - tetA, strB |
| F13.1 | AMP-CAZ-CF-XNL-NA-S-SPT-SXT-T | blactx, strA, strB, sul1, tet(A) | 1 | FIB | bla ctx-M, strB |
| F15.2 | AMP-CAZ-XNL-N--S-SXT-SPT-T | blactx-M, strA, strB, sul1, tet(A), tet(M), tetB | 1 | FIB, | bla ctx-M |
| F16.2 | AMP-CAZ-CF-XNL-NA-SXT-T | blactx-M, blaCMY-2, strA, strB, sul1, tet(A) | 1 | FIB | bla ctx, strB |
| F35.2 ctx | AMP-CAZ | blaCMY2, strB, | 1 | I1 | blaCMY-2 |
| F 42.2 | AMP-AMC-CAZ-XNL-NA-N-S | blaCMY-2, strB | 1 | B/O | bla-CMY 2, strB |
| F43.1 | AMP-AMC-XNL-C-NA-N-S-T | strB, tet(B), tet(M) | 0 | B/O, A/C | - |
| F46.2 | AMP-CAZ-NA-N-S-T | blaCMY-2, strB, tet(B) | 1 | FIB | FIB, iss, iutA, iroN |
| F47.2 | AMP-AMC-XNL-ENO-NA-N-S | blatem, strB, IS133 | 1 | FIB, I1, | strB |
| F39.2 | AMP, S, CIP, CAZ, T | blaCMY-2, tetA, strA | 1 | F1C, P | blaCMY-2 |
| F53.1 | AMP | blaCMY2, strB | 1 | I1 | I1, blaCMY-2, strB |
| FS1 | AMP, N, T | blaCMY2, tet (A) | ND | FIB | - |
| FS7 | AMP, T | blaCMY2 | 1 | FIB | blaCMY2, cvaC |
| FS4 | AMP, T | blaCMY2 | ND | FIB | - |
| Water | |||||
| NC5.1ctx 9/17/14 | AMP-CAZ-XNL-S-SXT--T | blactx, strB, sul1, tet(A), tet(M) | 1 | FIB, Frep | bla ctx-M, strB |
| NC6.4ctx 9/17/14 | T | tet(A) | 1 | FIB, FIC | tet(A), iroN, iss, iutA |
| SW2.1 1/21/15 | Amp-NA-S | blaCMY-2, strB | 1 | FIB, FI1 | blaCMY-2 |
| RP3.2 2/27/15 | AMP-AMC-CTX-XNL-ENO-NA-N | blaCMY-2 | 0 | I1, B/O | - |
| RS 1.1 6/22/16 | AMP-S-T | blaCMY-2, tet(A), strA | 1 | FIB | blaCMY-2 |
| RS1.3 6/22/16 | AMP-CAZ-CF-XNL-S-SXT-C | bla(CTX-M), tet(B), strA | 0 | FIB | strA |
| SW2.2 6/22/16 | T (tet) | tet(A) | ND | I1, FIB, FIA | ND |
| RS2.2 6/22/16 | AMP-S-T | strA, tet(M), tet(A) | 1 | FIB, | tet(A), ompT, hlyF |
| NC6.1 6/22/16 | AMP-T | blaCMY-2, tet(A) | 1 | FIB, FIA | tet(A), iss, ompT, hlyF |
| NC6.2 6/22/16 | AMP-T | blactx, tet(A) | 1 | FIB, FIA | tet(A), iss, ompT, hlyF |
| NC1.1 7/9/16 | AMP, S | blaCMY-2 | 1 | No ID | blaCMY-2 |
| NC1.2 10/3/16 | S | strB | 0 | FIB | - |
| NC1.3 1 10/3/16 | Amp-S-SXT | blaCMY-2 | 1 | No ID | blaCMY2 |
| RS2.3 10/3/16 | S | strB | ND | FIB | ND |
| SW8.1 10/4/16 | AMP-S-SXT | strA | ND | FIB | ND |
| RS1.1 1/31/17 | SXT-T | tet(A), tet(D) | ND | I1 | ND |
| NC6.1 1/31/17 | T (tet) | tet(A) | 1 | FIB, FIA | tet(A) |
| RS2.2 1/31/17 | AMP-CTX-S, T | tet(A), tet(B), aadA strB | 1 | FIB | tet(A), tet(B) |
| NC1.2 1/31/17 | AMP-T | bla-CMY-2, tet(M) | 1 | ND | blaCMY-2 |
| RS2.3 1/31/17 | AMP-S-SXT-T | strB, aadE, tet(B) | 1 | FIB | tet(B) |
| NC1.3 1/31/17 | AMP-S-SXT-T | blaCMY-2, tet(B) | 1 | FIB | blaCMY-2, tet(B) |
| SW2.2 1-31-17 | SXT-T | tet(A), tet(D) | 0 | FIB | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sen, K.; Shepherd, V.; Berglund, T.; Quintana, A.; Puim, S.; Tadmori, R.; J. Turner, R.; Khalil, L.; Soares, M.A. American Crows as Carriers of Extra Intestinal Pathogenic E. coli and Avian Pathogenic-Like E. coli and Their Potential Impact on a Constructed Wetland. Microorganisms 2020, 8, 1595. https://doi.org/10.3390/microorganisms8101595
Sen K, Shepherd V, Berglund T, Quintana A, Puim S, Tadmori R, J. Turner R, Khalil L, Soares MA. American Crows as Carriers of Extra Intestinal Pathogenic E. coli and Avian Pathogenic-Like E. coli and Their Potential Impact on a Constructed Wetland. Microorganisms. 2020; 8(10):1595. https://doi.org/10.3390/microorganisms8101595
Chicago/Turabian StyleSen, Keya, Vaughn Shepherd, Tanner Berglund, Alexa Quintana, Shnia Puim, Rama Tadmori, Robert J. Turner, Laura Khalil, and Marilia A. Soares. 2020. "American Crows as Carriers of Extra Intestinal Pathogenic E. coli and Avian Pathogenic-Like E. coli and Their Potential Impact on a Constructed Wetland" Microorganisms 8, no. 10: 1595. https://doi.org/10.3390/microorganisms8101595
APA StyleSen, K., Shepherd, V., Berglund, T., Quintana, A., Puim, S., Tadmori, R., J. Turner, R., Khalil, L., & Soares, M. A. (2020). American Crows as Carriers of Extra Intestinal Pathogenic E. coli and Avian Pathogenic-Like E. coli and Their Potential Impact on a Constructed Wetland. Microorganisms, 8(10), 1595. https://doi.org/10.3390/microorganisms8101595