Bacterial Community Dynamics Distinguish Poultry Compost from Dairy Compost and Non-Amended Soils Planted with Spinach

 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Field Experimental Design
2.2. Compost Treatments
2.3. Agronomic Practices
2.4. Soil Sampling
2.5. Bacterial and Fungal Community Composition
2.6. Microbial Ecoenzymatic Activity
2.7. Statistical Analysis
3. Results
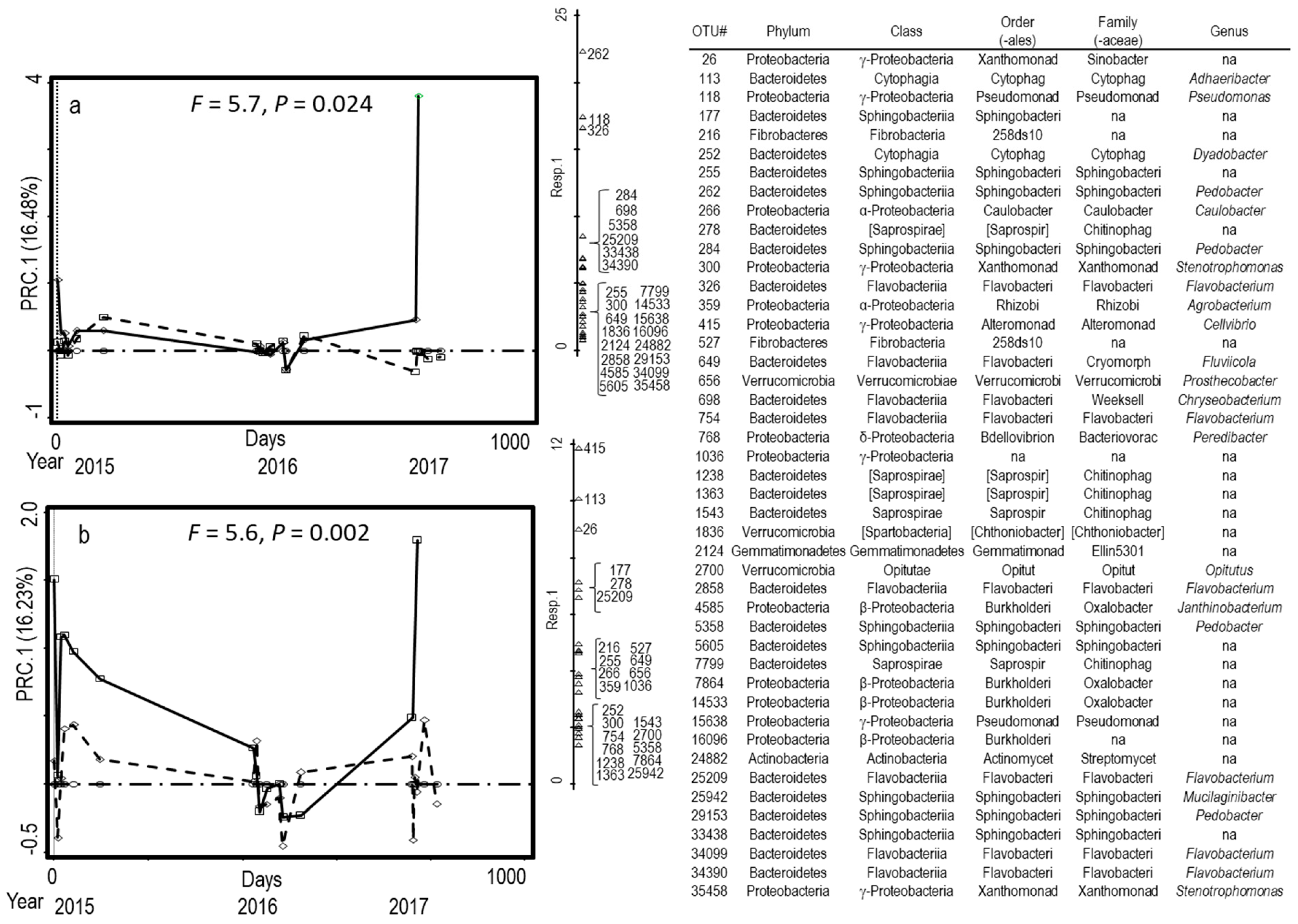
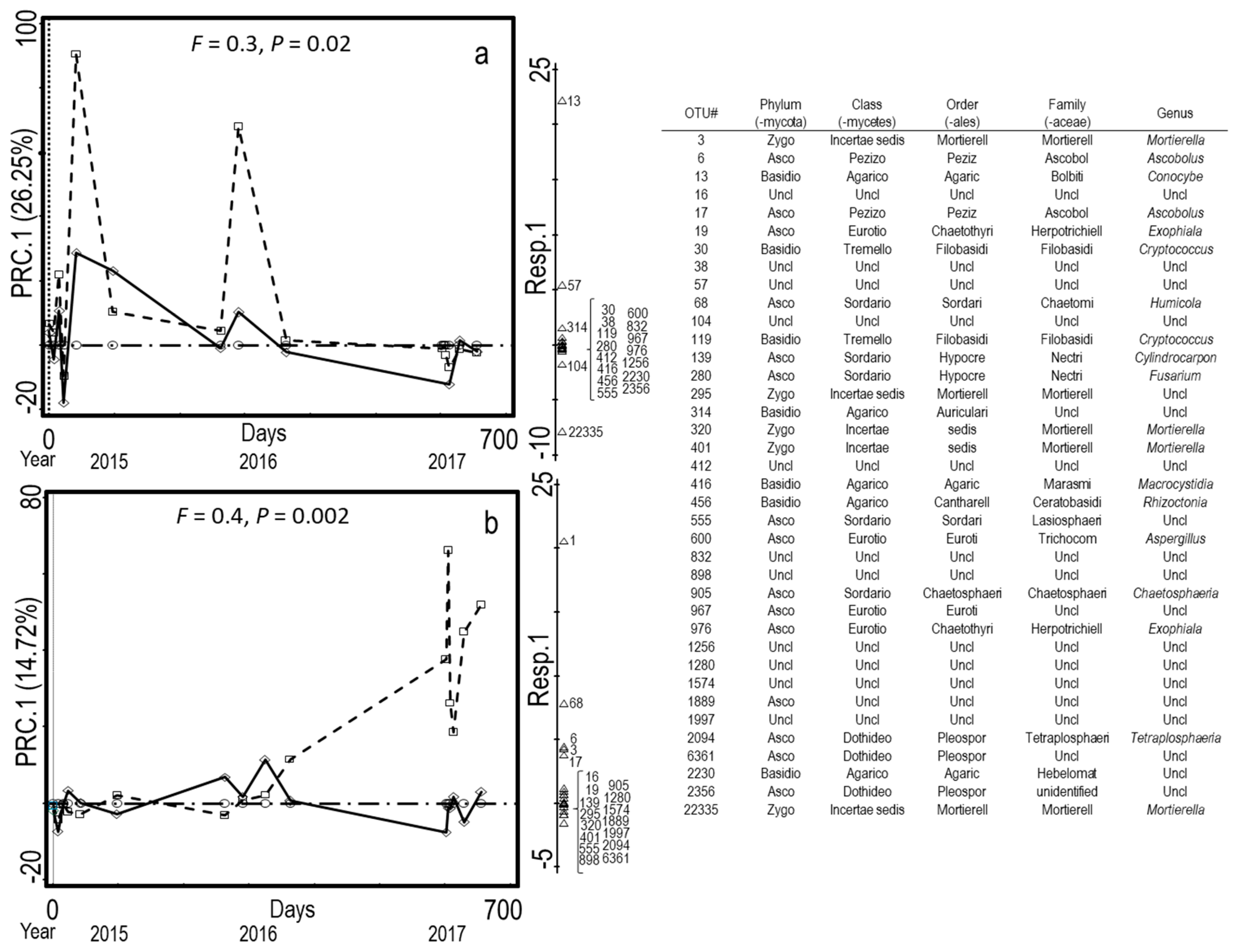
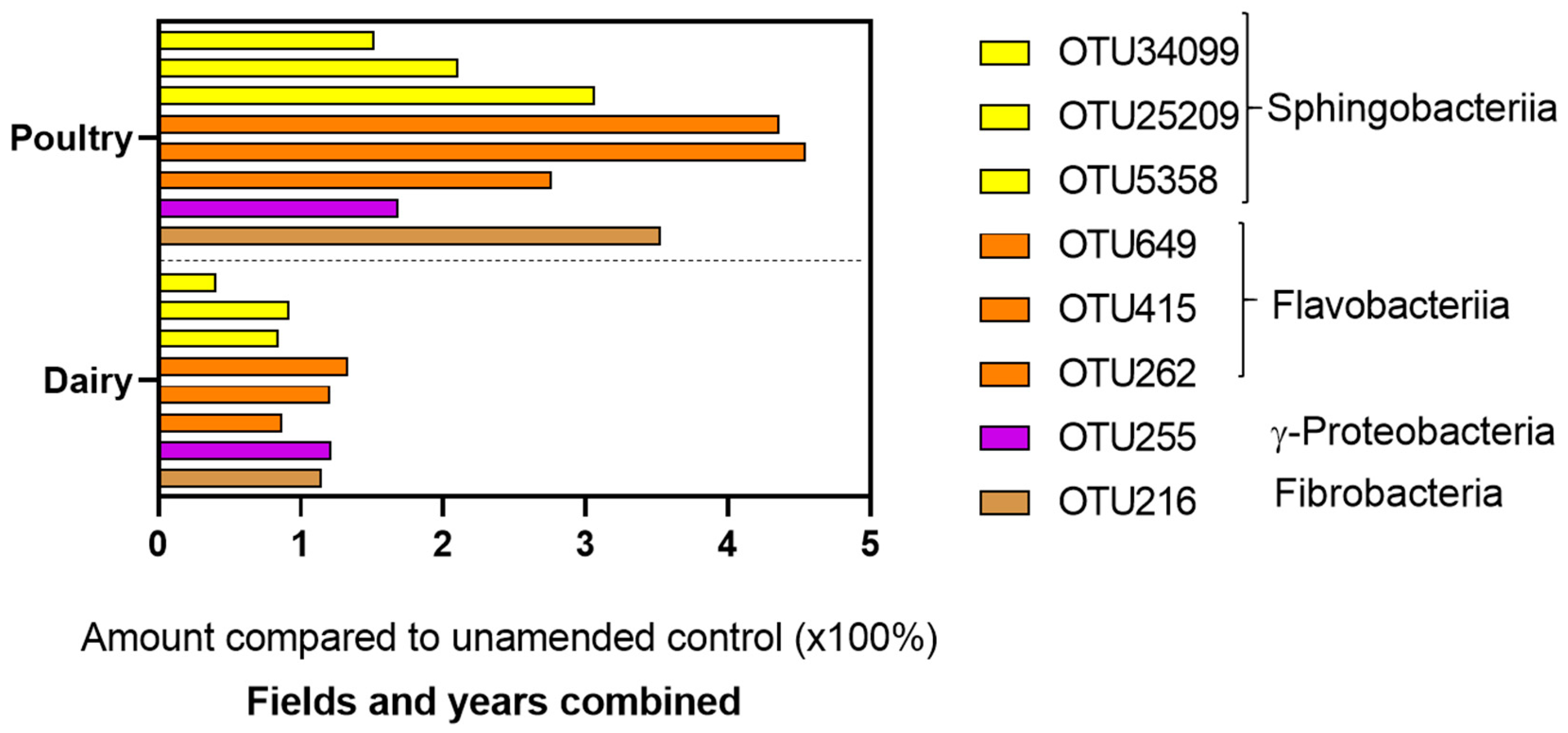
3.1. Microbial Composition of Dairy and Poultry Based Composts
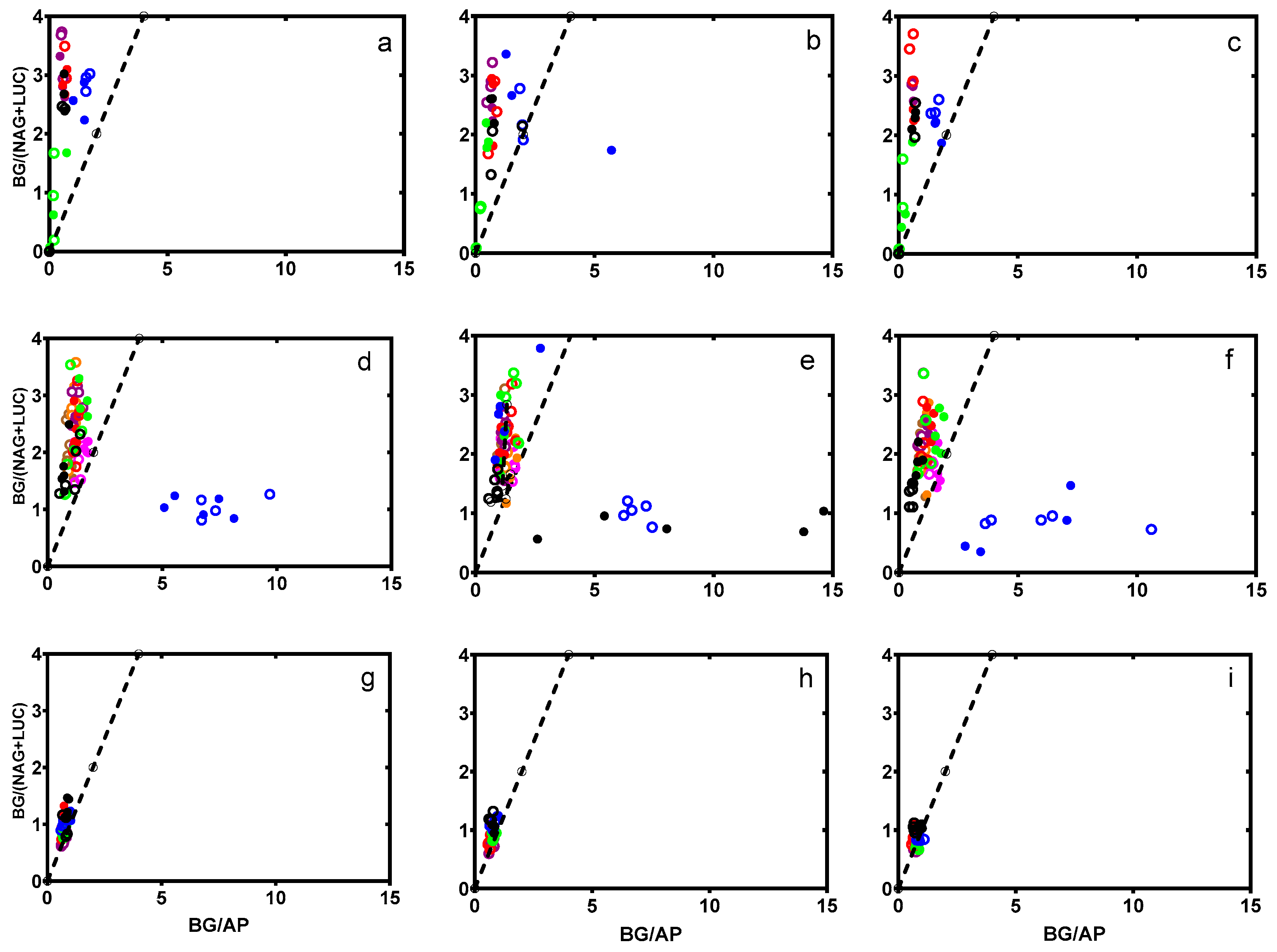
3.2. Microbial Nutrient Acquisition
4. Discussion
4.1. Microbial Composition of Dairy and Poultry Manure-Based Composts
4.2. Microbial Nutrient Acquisition
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Astill, G.; Minor, T.; Thornsbury, S. Changes in U.S. produce grower food safety practices from 1999 to 2016. Food Control 2019, 104, 326–332. [Google Scholar] [CrossRef]
- Poultry Litter. Available online: https://www.casella.com/products/earthlife-products/fertilizer/poultry-litter (accessed on 6 November 2019).
- Kyakuwaire, M.; Olupot, G.; Amoding, A.; Nkedi-Kizza, P.; Ateenyi Basamba, T. How safe is chicken litter for land application as an organic fertilizer?: A review. Int. J. Environ. Res. Public Health 2019, 16, 3521. [Google Scholar] [CrossRef] [Green Version]
- Compost Definition. Available online: https://www.compostingcouncil.org/page/CompostDefinition (accessed on 6 November 2019).
- Neher, D.A.; Weicht, T.R.; Dunseith, P. Compost for management of weed seeds, pathogen, and early blight on brassicas in organic farmer fields. Agroecol. Sust. Food. 2015, 39, 3–18. [Google Scholar] [CrossRef]
- Hadar, Y.; Papadopoulou, K.K. Suppressive composts: Microbial ecology links between abiotic environments and healthy plants. Annu. Rev. Phytopathol. 2012, 50, 133–153. [Google Scholar] [CrossRef]
- Neher, D.A.; Weicht, T.R.; Bates, S.T.; Leff, J.W.; Fierer, N. Changes in bacterial and fungal communities across compost recipes, preparation methods, and composting times. PLoS ONE 2013, 8, e79512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sidhu, J.; Gibbs, R.A.; Ho, G.E.; Unkovich, I. The role of indigenous microorganisms in suppression of Salmonella regrowth in composted biosolids. Water Res. 2001, 35, 913–920. [Google Scholar] [CrossRef]
- Gilbert, T. Feeding Community Food Scraps to Laying Hens in an Active Composting Systems. Available online: http://www.nvda.net/files/Feeding%20Food%20Scraps%20Manual%20.pdf (accessed on 6 November 2019).
- Eckstrom, K.; Barlow, J.W. Resistome metagenomics from plate to farm: The resistome and microbial composition during food waste feeding and composting on a Vermont poultry farm. PLoS ONE 2019, 14, e0219807. [Google Scholar] [CrossRef] [Green Version]
- Manure Composting as a Pathogen Reduction Strategy. Available online: http://www.omafra.gov.on.ca/english/engineer/facts/05-021.htm#what (accessed on 9 August 2019).
- Neher, D.A.; Hointink, H.A. Compost use for plant disease suppression. On-Farm Composting Handbook, 2nd ed. Rynk, R., Ed.; in press.
- Lugtenberg, B.; Kamilova, F. Plant-growth-promoting rhizobacteria. Annu. Rev. Microbiol. 2009, 63, 541–556. [Google Scholar] [CrossRef] [Green Version]
- Vacheron, J.; Desbrosses, G.; Bouffaud, M.L.; Touraine, B.; Moenne-Loccoz, Y.; Muller, D.; Legendre, L.; Wisniewski-Dye, F.; Prigent-Combaret, C. Plant growth-promoting rhizobacteria and root system functioning. Front. Plant Sci. 2013, 4, 356. [Google Scholar] [CrossRef] [Green Version]
- Sinsabaugh, R.L.; Belnap, J.; Findlay, S.G.; Shah, J.J.F.; Hill, B.H.; Kuehn, K.A.; Kuske, C.R.; Litvak, M.E.; Martinez, N.G.; Moorhead, D.L.; et al. Extracellular enzyme kinetics scale with resource availability. Biogeochemistry 2014, 121, 287–304. [Google Scholar] [CrossRef]
- Neher, D.A.; Cutler, A.J.; Weicht, T.R.; Sharma, M.; Millner, P.D. Composts of poultry litter or dairy manure differentially affect survival of enteric bacteria in fields with spinach. J. Appl. Microbiol. 2019, 126, 1910–1922. [Google Scholar] [CrossRef] [PubMed]
- Neher, D.A.; Fang, L.; Weicht, T.R. Ecoenzymes as indicators of compost to suppress Rhizoctonia solani. Compost. Sci. Util. 2017, 25, 251–261. [Google Scholar] [CrossRef] [Green Version]
- Lauber, C.L.; Hamady, M.; Knight, R.; Fierer, N. Pyrosequencing-based assessment of soil pH as a predictor of soil bacterial community structure at the continental scale. Appl. Environ. Microb. 2009, 75, 5111–5120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edgar, R.C. UPARSE: Highly accurate OTU sequences from microbial amplicon reads. Nat. Methods 2013, 10, 996–998. [Google Scholar] [CrossRef] [PubMed]
- Emerson, J.B.; Keady, P.B.; Brewer, T.E.; Clements, N.; Morgan, E.E.; Awerbuch, J.; Miller, S.L.; Fierer, N. Impacts of flood damage on airborne bacteria and fungi in homes after the 2013 Colorado Front Range flood. Environ. Sci. Technol. 2015, 49, 2675–2684. [Google Scholar] [CrossRef]
- Wang, Q.; Garrity, G.M.; Tiedje, J.M.; Cole, J.R. Naive Bayesian classifier for rapid assignment of rRNA sequences into the new bacterial taxonomy. Appl. Environ. Microb. 2007, 73, 5261–5267. [Google Scholar] [CrossRef] [Green Version]
- McDonald, D.; Price, M.N.; Goodrich, J.; Nawrocki, E.P.; DeSantis, T.Z.; Probst, A.; Andersen, G.L.; Knight, R.; Hugenholtz, P. An improved Greengenes taxonomy with explicit ranks for ecological and evolutionary analyses of bacteria and archaea. ISME J. 2012, 6, 610–618. [Google Scholar] [CrossRef] [PubMed]
- Koljalg, U.; Nilsson, R.H.; Abarenkov, K.; Tedersoo, L.; Taylor, A.F.; Bahram, M.; Bates, S.T.; Bruns, T.D.; Bengtsson-Palme, J.; Callaghan, T.M.; et al. Towards a unified paradigm for sequence-based identification of fungi. Mol. Ecol. 2013, 22, 5271–5277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saiya-Cork, K.R.; Sinsabaugh, R.L.; Zak, D.R. The effects of long term nitrogen deposition on extracellular enzyme activity in an Acer saccharum forest soil. Soil Biol. Biochem. 2002, 34, 1309–1315. [Google Scholar] [CrossRef]
- ter Braak, C.J.F.; Šmilauer, P. Canoco Reference Manual and User’s Guide: Software for Ordination (version 5.0); Biometris: Wageningen, The Netherlands, 2012. [Google Scholar]
- Moorhead, D.L.; Sinsabaugh, R.L.; Hill, B.H.; Weintraub, M.N. Vector analysis of ecoenzyme activities reveal constraints on coupled C, N and P dynamics. Soil Biol. Biochem. 2016, 93, 1–7. [Google Scholar] [CrossRef]
- Sasaki, H.; Nonaka, J.; Otawa, K.; Kitazume, O.; Asano, R.; Sasaki, T.; Nakai, Y. Analysis of the structure of the bacterial community in the livestock manure-based composting process. Asian Austral. J. Anim. 2009, 22, 113–118. [Google Scholar] [CrossRef]
- Oakley, B.B.; Lillehoj, H.S.; Kogut, M.H.; Kim, W.K.; Maurer, J.J.; Pedroso, A.; Lee, M.D.; Collett, S.R.; Johnson, T.J.; Cox, N.A. The chicken gastrointestinal microbiome. FEMS Microbiol. Lett. 2014, 360, 100–112. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.; Wang, Y.; Li, Y.; Zhang, Y.; Liu, T.; Wang, Y.; Sharpton, T.J.; Zhu, W. Progressive colonization of bacteria and degradation of rice straw in the rumen by Illumina sequencing. Front. Microbiol. 2017, 8, 2165. [Google Scholar] [CrossRef] [PubMed]
- Thomas, F.; Hehemann, J.H.; Rebuffet, E.; Czjzek, M.; Michel, G. Environmental and gut Bacteroidetes: The food connection. Front. Microbiol. 2011, 2, 93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, H.; Narihiro, T.; Ma, X.; Zhang, X.X.; Ren, H.; Ye, L. Diverse aromatic-degrading bacteria present in a highly enriched autotrophic nitrifying sludge. Sci. Total Environ. 2019, 666, 245–251. [Google Scholar] [CrossRef] [PubMed]
- Hester, E.; Harpenslager, S.; van Diggelen, J.; Lamers, L.; Jetten, M.; Lüke, C.; Lücker, S.; Welte, C. Linking nitrogen load to the structure and function of wetland soil and rhizosphere microbial communities. mSystems. 2018, 3, e00214–e00217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fierer, N.; Bradford, M.A.; Jackson, R.B. Toward an ecological classification of soil bacteria. Ecology 2007, 88, 1354–1364. [Google Scholar] [CrossRef] [PubMed]
- Han, X.M.; Hu, H.W.; Chen, Q.L.; Yang, L.Y.; Li, H.L.; Zhu, Y.G.; Li, X.Z.; Ma, Y.B. Antibiotic resistance genes and associated bacterial communities in agricultural soils amended with different sources of animal manures. Soil Biol. Biochem. 2018, 126, 91–102. [Google Scholar] [CrossRef]
- Yang, Z.; Peng, H.; Lu, X.; Liu, Q.; Huang, R.; Hu, B.; Kachanoski, G.; Zuidhof, M.J.; Le, X.C. Arsenic metabolites, including n-acetyl-4-hydroxy-m-arsanilic acid, in chicken litter from a roxarsone-feeding study involving 1600 chickens. Environ. Sci. Technol. 2016, 50, 6737–6743. [Google Scholar] [CrossRef] [Green Version]
- Vollú, R.E.; Cotta, S.R.; Jurelevicius, D.; Leite, D.C.d.A.; Parente, C.E.T.; Malm, O.; Martins, D.C.; Resende, Á.V.; Marriel, I.E.; Seldin, L. Response of the bacterial communities associated with maize rhizosphere to poultry litter as an organomineral fertilizer. Front. Environ. Sci. 2018, 6, 118. [Google Scholar] [CrossRef] [Green Version]
- Udikovic-Kolic, N.; Wichmann, F.; Broderick, N.A.; Handelsman, J. Bloom of resident antibiotic-resistant bacteria in soil following manure fertilization. PNAS 2014, 111, 15202–15207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, H.; Carvalhais, L.C.; Crawford, M.; Dang, Y.P.; Dennis, P.G.; Schenk, P.M. Strategic tillage increased the relative abundance of Acidobacteria but did not impact on overall soil microbial properties of a 19-year no-till Solonetz. Biol. Fert. Soils 2016, 52, 1021–1035. [Google Scholar] [CrossRef]
- Erickson, M.C.; Habteselassie, M.Y.; Liao, J.; Webb, C.C.; Mantripragada, V.; Davey, L.E.; Doyle, M.P. Examination of factors for use as potential predictors of human enteric pathogen survival in soil. J. Appl. Microbiol. 2014, 116, 335–349. [Google Scholar] [CrossRef] [PubMed]
- van Bruggen, A.H.C.; Gamliel, A.; Finckh, M.R. Plant disease management in organic farming systems. Pest Manag. Sci. 2016, 72, 30–44. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter * | Lilac Soil | Wheelock Soil | Dairy Manure Compost (DCM) | Poultry Litter Compost (PLC) |
|---|---|---|---|---|
| Total solids (%) | ND † | ND | 62.7 | 81.5 |
| Total carbon (%) | ND | ND | 27.3 | 22.7 |
| Total nitrogen (%) | ND | ND | 1.87 | 3.09 |
| Total potassium (%) | 2.4 × 10−3 | 1.2 × 10−2 | 1.07 | 3.8 |
| Total phosphorus (%) | 2.8 × 10−3 | 1.4 × 10−3 | 0.22 | 2.1 |
| Total calcium (%) | 1.3 × 10−1 | 1.2 × 10−1 | 1.00 | 3.0 |
| Total magnesium (%) | 1.1 × 10−2 | 9.7 × 10−3 | 0.29 | 0.57 |
| Total sodium (%) | 5.3 × 10−4 | 6.0 × 10−4 | 0.2 | 1.4 |
| Volatile solids (%) | ND | ND | 54.0 | 50.2 |
| pH | 6.2 | 6.2 | 7.9 | 7.7 |
| Bulk density (lbs yd−3) | ND | ND | 430 | 910 |
| Bulk density (kg m−3) | ND | ND | 255.1 | 539.9 |
| Conductivity (dS m−1) | ND | ND | 7.1 | 29.1 |
| C:N ratio | ND | ND | 14.6 | 7.4 |
| NH4-N (mg kg−1) | 3.2 | 3.3 | 29.2 | 1510 |
| NO3-N (mg kg−1) | 15.8 | 19.5 | 1.11 | 816 |
| Total boron (mg kg−1) | 0.2 | 0.2 | 19.4 | 65.6 |
| Total copper (mg kg−1) | 0.2 | 0.2 | 237 | 458 |
| Total iron (mg kg−1) | 4.3 | 4.2 | 539 | 1390 |
| Total manganese (mg kg−1) | 4.8 | 5.5 | 51.3 | 634 |
| Total zinc (mg kg−1) | 2.7 | 1.9 | 57.8 | 540 |
| Year | Season Duration (dpc) * | Cum Rainfall (cm) | Mean (SD) Air Temp (°C) † | Dairy Manure Compost (t ha−1) | Poultry Litter Compost (t ha−1) |
|---|---|---|---|---|---|
| 2015 | 57 | 25.7 | 21.2 (2.86) | 15.1 | 13.4 |
| 2016 | 64 | 15.8 | 21.3 (3.45) | 5.4 | 6.8 |
| 2017 | 53 | 21.8 | 17.1 (5.11) | 9.6 | 12.2 |
| Year | Compost Addition | Ecoenzymes (dpc *) | DNA (dpc) † |
|---|---|---|---|
| 2015 | June 2 | 8, 16, 23, 30, 50, 65 | 16S and ITS: 8, 15, 23, 30, 49, 104 ‡ |
| 2016 | May 31 | 2, 8, 15, 30, 36, 43, 57, 64 | 16S: 2, 8, 15, 30, 56, 63, 99 ‡ ITS: 2, 30, 99 ‡ |
| 2017 | May 6 | 3, 6, 11, 27, 53 | 16S and ITS: 3, 6, 11, 27, 53 |
| OTU | Phylum | Class | Order | Family | Genus |
|---|---|---|---|---|---|
| 216 | Fibrobacteres | Fibrobacteria | 258ds10 | ||
| 255 | Bacteroidetes | Sphingobacteriia | Sphingobacteriales | Sphingobacteriaceae | |
| 262 | Bacteroidetes | Sphingobacteriia | Sphingobacteriales | Sphingobacteriaceae | Pedobacter |
| 415 | Proteobacteria | γ-proteobacteria | Alteromonadales | Alteromonadaceae | Cellvibrio |
| 649 | Bacteroidetes | Flavobacteriia | Flavobacteriales | Cryomorphaceae | Fluviicola |
| 5358 | Bacteroidetes | Sphingobacteriia | Sphingobacteriales | Sphingobacteriaceae | Pedobacter |
| 25209 | Bacteroidetes | Flavobacteriia | Flavobacteriales | Flavobacteriaceae | Flavobacterium |
| 34099 | Bacteroidetes | Flavobacteriia | Flavobacteriales | Flavobacteriaceae | Flavobacterium |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Neher, D.A.; Limoges, M.A.; Weicht, T.R.; Sharma, M.; Millner, P.D.; Donnelly, C. Bacterial Community Dynamics Distinguish Poultry Compost from Dairy Compost and Non-Amended Soils Planted with Spinach. Microorganisms 2020, 8, 1601. https://doi.org/10.3390/microorganisms8101601
Neher DA, Limoges MA, Weicht TR, Sharma M, Millner PD, Donnelly C. Bacterial Community Dynamics Distinguish Poultry Compost from Dairy Compost and Non-Amended Soils Planted with Spinach. Microorganisms. 2020; 8(10):1601. https://doi.org/10.3390/microorganisms8101601
Chicago/Turabian StyleNeher, Deborah A., Marie A. Limoges, Thomas R. Weicht, Manan Sharma, Patricia D. Millner, and Catherine Donnelly. 2020. "Bacterial Community Dynamics Distinguish Poultry Compost from Dairy Compost and Non-Amended Soils Planted with Spinach" Microorganisms 8, no. 10: 1601. https://doi.org/10.3390/microorganisms8101601
APA StyleNeher, D. A., Limoges, M. A., Weicht, T. R., Sharma, M., Millner, P. D., & Donnelly, C. (2020). Bacterial Community Dynamics Distinguish Poultry Compost from Dairy Compost and Non-Amended Soils Planted with Spinach. Microorganisms, 8(10), 1601. https://doi.org/10.3390/microorganisms8101601