1. Introduction
Soil fertility and nutrient cycling are influenced by the soil microbial population. An understanding of the interrelationships between functional microbial communities and cycling of C, N, P and S as an indicator of soil health is essential for soil fertility management [
1]. The major shortcomings associated with the use of chemical fertilizers include high cost, limited supply and environmental polluting effects (especially from N and P fertilizers). Thus, there is a need for alternative and sustainable sources of nutrients for crops. The adoption of an integrated nutrient management (INM) approach offers an alternative strategy against the adverse effects of using conventional N and P fertilizers alone. An INM system involves minimal chemical fertilizer application in combination with both natural and man-made sources of crop nutrients to improve nutrient-use efficiency and crop productivity in an environmental-friendly manner [
2]. In addition, advances in the understanding of the relationships between crops and microorganisms colonizing rhizospheric soils have increased screening efforts for microbial strains showing plant growth-promoting (PGP) ability. This ability is through direct and/or indirect enhancement of plant nutrient uptake by use as microbial inoculants or in microbial-based fertilizers termed as bio-fertilizers.
Plant growth-promoting rhizobacteria (PGPR) are free-living soil bacteria which when applied to seed or crops, propagate aggressively in the rhizosphere or plant roots and impact crop growth and production [
3]. Thus, beneficial crop-microbe interactions in the rhizosphere could improve crop vigor and soil fertility [
4]. Most PGP effects of PGPR are by means of metabolite production that perfunctorily enhance growth. Thus, PGPR influence crop growth and development by modifying the physiological conditions and morphological characteristics of inoculated roots to improve nutrient uptake [
5,
6,
7,
8]. As an example, plants treated with
Azospirillum exhibited increased N, P, K use efficiency and microelements uptake [
9]. Bio-fertilizers exert better crop growth effects via diverse or a combination of mechanisms which include improved soil nutrient availability, biological N
2 fixation, phytohormone production and disease antagonism [
10].
The genus
Burkholderia comprises diverse species and many strains have been found in human and animal samples in addition to environmental sources [
11]. The plant growth-promoting rhizobacteria status of many
Burkholderia species has been well established because of the many PGP mechanisms shown by the strains. Some have been reported to possess N-fixing ability as free-living or endophytic bacteria [
12] or by forming a symbiotic relationship with leguminous plants [
13,
14]. Some strains have been reported to be phosphate-solubilizing bacteria (PSB) [
15,
16]. Others with the ability to inhibit many plant-disease-causing fungi are being used as biocontrol agents [
16,
17]. Among other known features of the strains of this genus include phytohormone and/or siderophore production [
18]. However, the genus
Burkholderia has recently been divided into two genera with the novel genus
Paraburkholderia [
19,
20,
21] bearing primarily environmental
Burkholderia species which are cited as the good or beneficial ones [
19,
22,
23].
Burkholderia sensu stricto genus consists of environmental, clinical and phytopathogenic species that are harmful [
19,
24].
Paraburkholderia nodosa strains reported as legume-symbionts have been isolated from root nodules of
Mimosa (
M. bimucronata and
M. scabrella) [
25] and other plants from the “Piptadenia group” [
26]. More recently
P. nodosa strains were also recovered from members of the
Papilionoidea subfamily such as in
Phaseolus vulgaris [
27]. However, information on free-living
P. nodosa strains and their potential PGP effects on non-legume plants such as maize is lacking.
The environmental
Burkholderia cepacia is classified as one of the
Burkholderia cepacia complex (Bcc) strains, in which the latter are opportunistic pathogens in patients with cystic fibrosis and immuno-compromised individuals [
28]. The Bcc strains have since been severely restricted in agricultural usage [
28]. Nevertheless, many
Burkholderia cepacia strains have been reported to possess considerable biotechnological potentials such as PGPR [
29] and biological control of plant pathogens [
30,
31]. In terms of phosphate-solubilization, many studies have found
Burkholderia cepacia strains to be responsible for P release from polyphosphate in low pH environments [
32,
33,
34,
35]. Similarly,
Burkholderia cepacia, a PGPR strain isolated from Korean agricultural soils solubilized insoluble phosphates, owing to its large production of organic acids especially gluconic acid [
36].
The genus
Serratia belongs to the family
Enterobacteriaceae of the class
Gammaproteobacteria. Although some members of this genus, in particularly
S. marcescens, have clinical importance as human pathogens [
37,
38], many
Serratia species were found to be plant-associated beneficial bacteria [
39]. For instance,
S. plymuthica enhanced the growth of plants and inhibited soil-borne plant pathogens [
40];
S. liquefaciens,
S. plymuthica and
S. rubidaea associated with the rhizosphere of oilseed rape was shown to have antifungal properties [
41] and an association of
S. rubidaea with marine alga [
42]. Furthermore,
S. nematodiphila, along with
S. entomophila,
S. glossinae,
S. proteamaculans and
S. ureilytica have been reported as non-pathogenic to humans [
39].
The first report on novel
Serratia nematodiphila was of the strain symbiont with the entomopathogenic nematode
Heterorhabditidoides chongmingensis (Rhabditida: Rhabditidae) and it was isolated from the latter’s intestine [
43]. The
Serratia nematodiphila strain was non-cellulolytic [
44]. There have been many reports on the beneficial effects of
Serratia nematodiphila such as a multiple-PGPR-trait strain improving black pepper plant growth [
4], gibberellin-producing-PGPR strain improving growth of pepper plants under low-temperature stress [
45] and an endophytic bacterial strain LRE07 reducing growth suppression of
Solanum nigrum L. grown in a cadmium-polluted soil [
46]. However, there is a dearth of information on the cellulolytic strain
Serratia nematodiphila as PGPR and its effects on maize plants.
There are several research reports on the beneficial impacts of PGPR on maize growth. Bacterial genera such as
Azospirillum,
Azotobacter,
Acetobacter,
Pseudomonas,
Paraburkholderia,
Herbaspirillum and
Rhizobium have been reported to show good PGPR effects on maize. Noumavo et al. [
47] found that
Azospirillum lipoferum-treated plants produced maximal plant heights and underground biomass by 37.32% and 56%, respectively, whereas a combination of
Pseudomonas fluorescens and
P.
putida produced higher aerial dry matter by 59.11% compared with the control. Co-inoculation of
Bacillus megaterium,
Azotobacter chroococcum and
Bacillus mucilaginous has also been reported to significantly improve maize biomass and height with comparable effects 50% higher than that with chemical fertilizer alone [
48].
Therefore, the objectives of this study were to determine the effects of beneficial cellulolytic Serratia nematodiphila C46d, N-fixing Paraburkholderia nodosa NB1 and phosphate-solubilizing Burkholderia cepacia PB3 on: (1) interactions among functional rhizobacteria, (2) selected growth variables of maize, (3) plant growth-promotion based on nutrient uptake and use efficiency of maize and (4) selected properties of the Bekenu series soil. It was hypothesized that the right amount of chemical fertilizer and compost combination with selected PGPR inoculants in single or consortium treatment used in this present study could exert positive influences on maize growth and nutrient use efficiency on Bekenu series (Typic Paleudults).
2. Materials and Methods
2.1. Bacterial Inoculants
Bacterial cultures of
Serratia nematodiphila C46d,
Paraburkholderia nodosa NB1 and
Burkholderia cepacia PB3 isolated from a rehabilitated forest soils at Universiti Putra Malaysia Bintulu Sarawak Campus, Malaysia were used in this study. All the isolates were previously identified based on 16S rRNA gene sequence similarities and were also characterized based on morphological and biochemical properties [
49] using Bergey’s Manual of Determinative Bacteriology and Bergey’s Manual of Systematic Bacteriology [
50,
51].
The functional and cross-functional activity indices of the three isolates were calculated by ratio of total diameter (colony + halo or clearing zone around colonies) and colony diameter [
52] on selective media.
Phosphate-solubilization activity was evaluated on tricalcium phosphate medium (modified from Nautiyal [
53], Buis [
54], Gupta et al. [
55] and Kokal [
56]): Prepared using a 50 mL salts solution (containing 100.0 g L
−1 MgCl
2·6H
2O, 5.0 g L
−1 MgSO
4·7H
2O, 4.0 g L
−1 KCl and 2.0 g L
−1 (NH
4)
2SO
4), 10.0 g sucrose, 2.5 g Ca
3(PO
4)
2, 6 mL bromophenol blue (in 0.4% ethanol solution), 15.0 g agar, 1.0 L distilled water, pH 7.0. Cellulolytic activity was evaluated on cellulose-Congo red medium (modified from Hendricks et al. [
57]): Prepared using 0.50 g K
2HPO
4, 0.25 g MgSO
4, 1.88 g cellulose microgranular powder, 0.20 g Congo red (after pH modification), 15.0 g agar, 2.0 g gelatine, 100 mL soil extract (1:2 soil solution ratio), 900 mL tap water, pH 7.0. Nitrogen-fixation activity was evaluated on N-free malate (Nfb) medium (modified from Döbereiner and Day [
58]): Prepared using 0.4 g KH
2PO
4, 0.1 g K
2HPO
4, 0.2 g MgSO
4.7H
2O, 0.1 g NaCl, 0.02 g CaCl
2, 0.01 g FeCl
3, 0.002 g Mo.O
4Na.2H
2O, 5.0 g sodium malate, 5 mL bromothymol blue (in 0.5% alcohol solution), 15.0 g agar, 1.0 L distilled water, pH 7.0.
The three isolates possess phosphate-solubilizing ability with the phosphate-solubilizing isolate
Burkholderia cepacia PB3 exhibiting a phosphate-solubilization activity index value of 4.93, cellulolytic activity of 4.58 and N-fixing index value of 3.12. Cellulolytic isolate
Serratia nematodiphila C46d demonstrated a functional activity of 10.41 on cellulose agar media, phosphate-solubilization activity of 2.10 and 0.00 (zero) N-fixing ability. Nitrogen-fixing isolate
Paraburkholderia nodosa NB1 without cellulose-hydrolyzing capability had the lowest phosphate-solubilization activity index of 1.41 and showed the strongest intensity blue halo on Nfb medium with a N-fixing index value of 10.82 among all three isolates. These three isolates also produced phytohormone indole-3-acetic acid (IAA) which was determined according to Asghar et al. [
59], with
Serratia nematodiphila C46d,
Burkholderia cepacia PB3 and
Paraburkholderia nodosa NB1 generating IAA quantities of 11.39 µg mL
−1, 7.81 µg mL
−1 and 0.43 µg mL
−1, respectively.
2.2. Bacterial Compatibility or Interactions and Establishment of Bacterial Exponential Growth Equations
The three selected bacterial isolates used as inocula in this pot trial were observed for their positive or negative interactions with one another via cross streak assay method using nutrient agar medium [
60]. Absence of any inhibition zone at crossover areas is an indication of compatibility or non-antagonistic interaction among the tested isolates.
The growth of the aforementioned three bacterial isolates was studied so as to establish a modeling equation unique to each isolate for estimation of their population exponential growth at any given time. A few colonies from a purified plate culture were inoculated into 50 mL of nutrient broth in a 250 mL Erlenmeyer flask before agitating at 200 rpm at 30 °C. At 1 hr interval, starting from 0 hr or before beginning the assay by agitation, a portion of the well-mixed culture was dispensed for absorbance reading using a UV–Vis spectrophotometer at 600 nm and serial dilutions before plating on nutrient agar for colony-forming-unit (cfu) counting. The procedure was repeated hourly up to stationary phase as indicated by the observed trend of optical density (OD) values. Samples were diluted to be within the linear range of the dynamic optical system. The number of colonies formed on the plates was counted after 16 hrs of incubation or as soon as colonies were visible for counting. The cfu counts were plotted against OD readings (which were both selected at up to log growth phase) in triplicate datasets using a curve-fitting software (DataFit Version 9.0 by Oakdale Engineering) and each selected isolate‘s exponential growth equation was established.
2.3. Modulation and Preparation of Solid Carriers as Inocula
Two types of carrier formulations or materials were shortlisted to be optimized and tested, after which one was selected for the bacterial inoculation in the pot trial. The materials were peat moss and compost-charcoal powder mixture at a ratio 3:1. The peat moss was Tulip Profi No 1 from Hardenberg, Netherlands. Both formulations were oven dried at 70 °C for 72 hr, ground and sieved to pass a 2-mm sieve. The water-holding capacity of both formulations was determined so as to ascertain the maximum volume of bacterial culture to be inoculated or mixed into the carriers. Five grams of each carrier type was packed in small autoclavable plastic bags and amended with approximately 10% CaCO3 to maintain a neutral pH. Twenty percent gum arabic was added at 1 mL g−1 as carrier stabilizer and all components were then mixed well manually. They were then autoclaved at 121 °C for 20 min.
The three selected bacterial isolates NB1, PB3 and C46d were used as inoculants. Their cultures were prepared by inoculating each isolate’s pure plate culture colonies into 100 mL nutrient broth in 250 mL Erlenmeyer flasks. The flasks were then agitated at 200 rpm at 30 °C with several intermittent samplings in aseptic conditions for optical density checks, up to log phase of growth before harvesting. The bacteria were harvested by centrifugation at 6000 rpm at 4 °C before being aseptically transferred into both types of carriers to up to 50% water retention capacity level of each carrier type. The mean population sizes of NB1, PB3 and C46d were determined to be 9.78 × 106 cfu mL−1 g−1, 3.64 × 108 cfu mL−1 g−1 and 4.96 × 108 cfu mL−1 g−1, respectively following inoculation.
The inoculant-carriers were manually mixed before aseptically resealing the inocula packets. These were then placed for curing at room temperature and away from sunlight or heat for 3 days, followed by quality check via cfu counting. After the curing period the rest of the carrier-inoculants were transferred into a refrigerator for storage at 4 °C for further use with maximum storage time of 3 months. The inoculated peat moss was chosen as the carrier-inoculant for the pot trial based on its higher cfu counts by one logarithm compared to the compost-charcoal formulation after the curing period and prior to use. One day before the commencement of inoculation for the pot trial, the peat moss inoculums were incubated overnight at room temperature in the dark to allow bacterial cells to equilibrate or adjust to the higher temperature at the pot trial site. The compost used for the carrier formulations and the pot trial was locally supplied by Green Grow, Malaysia. The total N, available P and available K contents of the compost were 0.23%, 87.1 mg kg−1 and 714 mg kg−1, respectively.
2.4. Soil Characterization
The soil used for the pot trial was Bekenu Series (
Typic Paleudults). It was collected at 0 to 25 cm depth using an auger from an uncultivated area at Universiti Putra Malaysia Bintulu Sarawak Campus, Sarawak, Malaysia. The soil samples were air-dried, ground to pass a 2-mm sieve and mixed thoroughly. Soil bulk density and texture were determined using the methods according to Tan [
61]. The following soil properties were analyzed before and after the pot trial. Soil pH in distilled water and 1M KCl solution (both with a soil and solution ratio of 1:2.5) were determined using a pH meter [
62]; organic matter and total C via combustion method [
63]; total N using Kjeldahl method [
64]; and exchangeable NH
4+ and available NO
3- using Keeney and Nelson [
65] method. The soil available P and exchangeable cations (K, Ca and Mg) were extracted using double acid method [
66]. Then the concentration of soil available P was determined using the molybdenum blue method [
67] and Atomic Absorption Spectrophotometry (AAS) (Analyst 800, Perkin Elmer, Norwalk, USA) was used for the exchangeable cations. Soil CEC was determined by leaching method [
68] followed by steam distillation [
64]. The bulk density of the soil was 1.10 g cm
−3 and a sandy clay loam texture. Selected chemical properties of the soil are shown in
Table A1 [
69].
2.5. Pot Trial
A pot trial was conducted in a greenhouse at Universiti Putra Malaysia Bintulu Sarawak Campus, Malaysia using randomized complete block design (RCBD). The experimental site experiences an average temperature of 31.90 ± 3.20 °C, relative humidity of 69.40 ± 13.70% and light intensity of 34,500.00 ± 6362.10 lux. The plastic pots used in this study were sized Ø
bottom 19 cm × 23 cm x Ø
top 28 cm and each was filled with 7.15 kg soil (adjusted based on the bulk density of the soil and the size of the pots). Seven treatments including bacterial inoculations and uninoculated treatments were used. The treatments were: organic fertilizer and chemical fertilizer without any bacteria (OCF), organic fertilizer and chemical fertilizer with cellulolytic bacteria (TC), organic fertilizer and chemical fertilizer with N-fixing bacteria (TN), organic fertilizer and chemical fertilizer with P-solubilizing bacteria (TP), organic fertilizer and chemical fertilizer with the three bacterial types (TCNP), chemical fertilizer alone (CF) and no treatment at all (Control). The details and the codes of the treatments are shown in
Table 1.
Maize (var. F1 Thai hybrid) was used as the test crop. Fertilizer requirements for the maize based on the Malaysia Agriculture Research Development Institute (MARDI)’s recommendations were 60 kg ha
−1 N, 60 kg ha
−1 P
2O
5 and 40 kg ha
−1 K
2O. The chemical fertilizers used were urea (46% N), Egyptian Rock Phosphate (ERP) (28% P
2O
5) and muriate of potash (MOP) or KCl (60% K
2O). For treatment 7 there was 100% fertilization based on MARDI’s recommended rates which when scaled down to per pot basis was equivalent to 1.44 g urea, 5.87 g ERP and 3.58 g MOP. These fertilizers were applied in two equal splits at 10 and 28 days after seeding (DAS). The combined effects of bacterial inoculums, compost and chemical fertilizer at 50% total fertilization requirement of maize was compared to no fertilization (Control), organic-chemical fertilization (OCF) at 50% total requirement and 100% total requirement using chemical fertilizer alone (
Table 1).
Treatments 2 to 6 had the total fertilization level reduced to only 50% supplied mainly by a combination of compost and chemical fertilizer with/or without bacterial inoculants according to the treatment design. They were applied at 10 DAS only. The amounts of organic fertilizer (compost) and chemical fertilizers used in combination with treatments in this study were based on P content of the compost because it has been reported that basing organic fertilizer on N supply typically results in P addition in excess of crops needs [
70]. Five grams of peat moss-based bacterial inoculums were inoculated into the pots according to the treatment design.
Similar amounts of the autoclaved carrier-based inocula were also added to all the treatments without bacterial inoculation so as to maintain similarity of soil texture and other properties for all the pots. There were seven replications for each treatment. The maize seeds were washed for a few rounds to remove any fungicide coatings, followed by surface disinfection using 0.1% sodium hypochlorite solution for 5 min before rinsing at least three times with distilled water to thoroughly remove sterilants. The seeds were then soaked in water for 24 hrs to aid seed germination and better establishment of seedlings upon seeding.
The soil in each pot was moistened up to 70% field capacity using tap water prior to sowing the germinated seeds. Four seeds were sown directly into planting holes at 4 cm soil depth per pot and then the holes were loosely covered with soil from the surface to enable rapid emergence of the seeds. At seven DAS the seedlings were thinned to two per pot so as to reduce competition among the plants. Soil moisture was maintained at field capacity using tap water. The plants were monitored up to tasselling stage at 56 DAS. Tasselling stage is the final development stage of maize before it reaches productive stage [
71].
2.6. Biological and Chemical Analyses
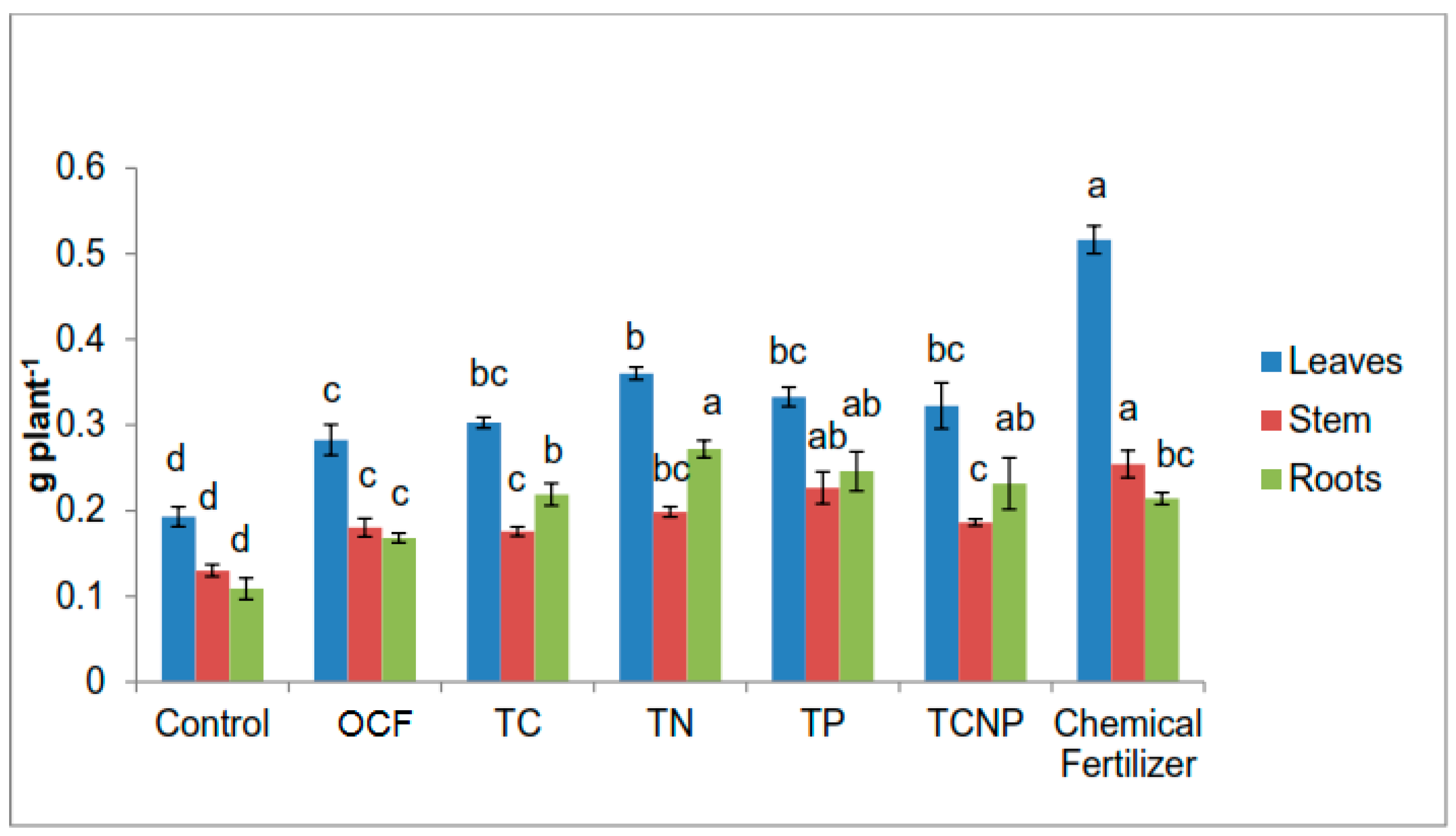
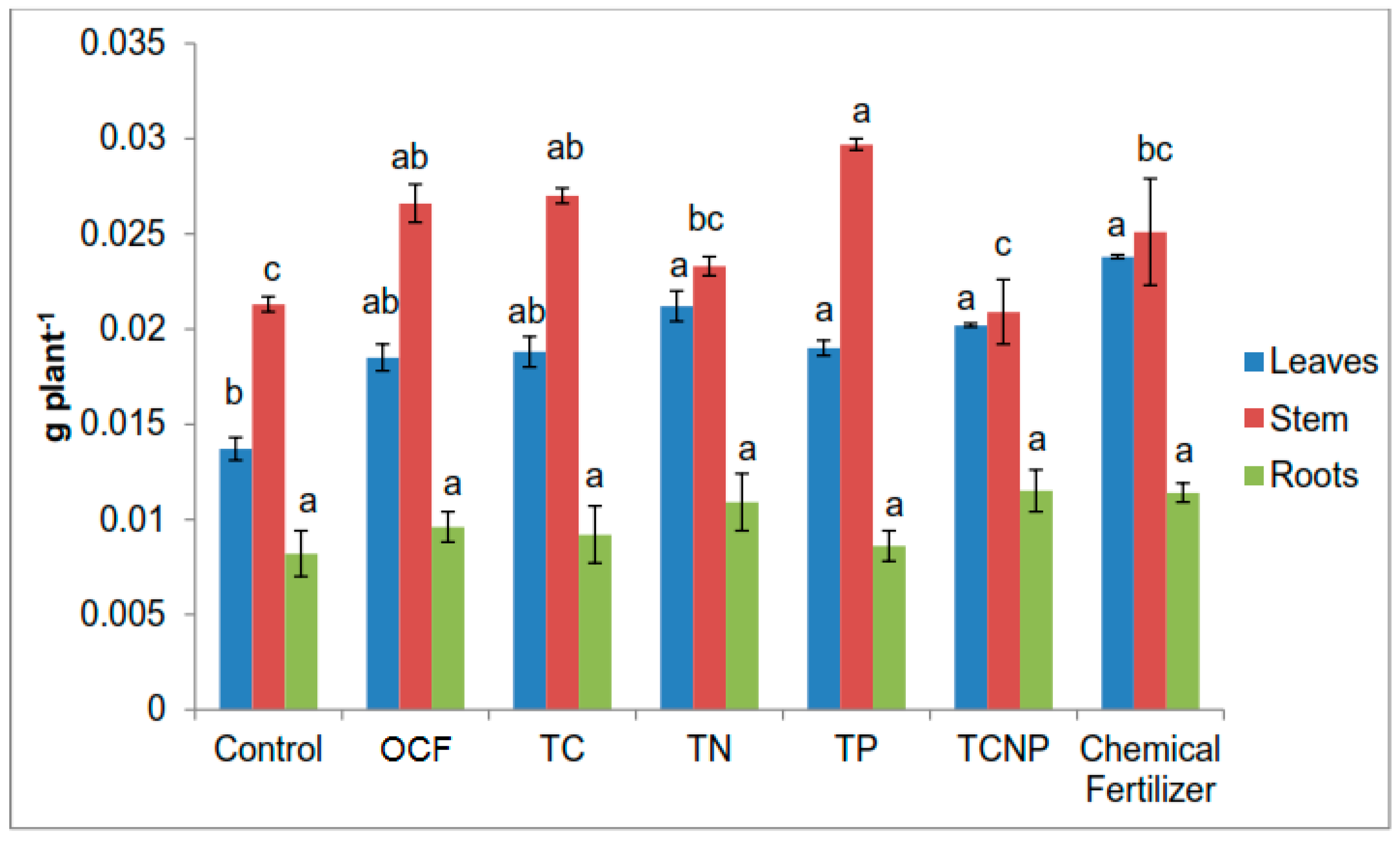
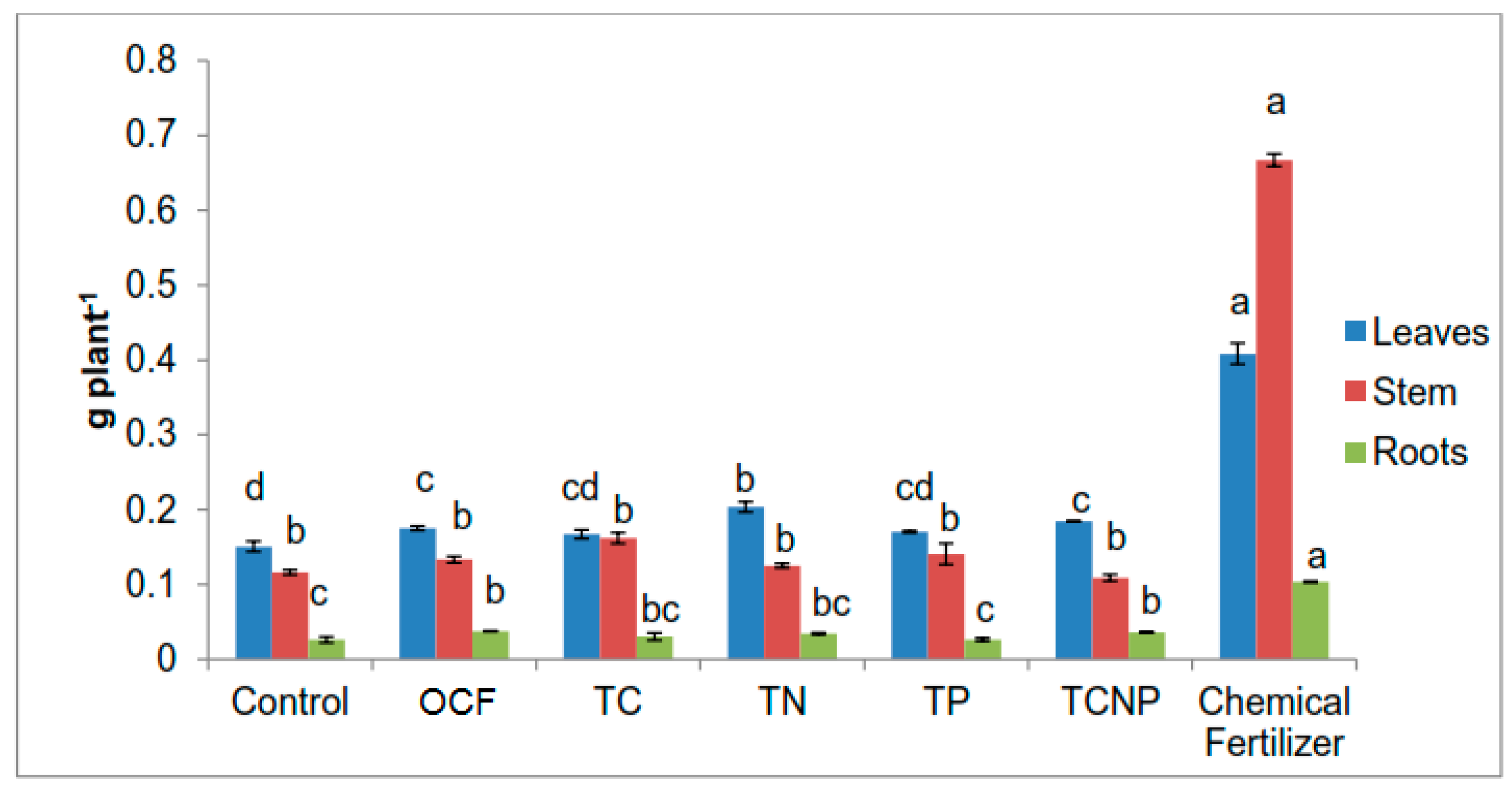
Growth variables of maize were determined based on height using a measuring tape and stem diameter at 10 cm from soil surface using a digital vernier calliper at 56 DAS. Chlorophyll content was also determined using a Minolta’s SPAD-502 chlorophyll meter. The crops were harvested at 56 DAS. The maize plants were partitioned into leaves, stem and roots. The plant parts were then washed with tap water, rinsed using distilled water and blotted gently to remove contaminants such as soil residues and dust followed by oven-drying at 60 °C to constant weight. The dry weight of the plant parts was measured using a digital balance. The oven-dried plant samples were ground using a grinder before being analyzed for total N content using Kjeldahl method [
64], and both P and K contents by dry ashing extraction [
63], after which P was analyzed using the molybdenum blue method [
67] and K by using an AAS. Nutrient uptake was calculated by multiplying nutrient concentration with the dry weight of the respective plant parts. Nutrient use efficiency was calculated according to the Pomares-Garcia and Pratt [
72] equation:
A = uptake with fertilizer; B = uptake without fertilizer; C = Total amount of fertilizer applied
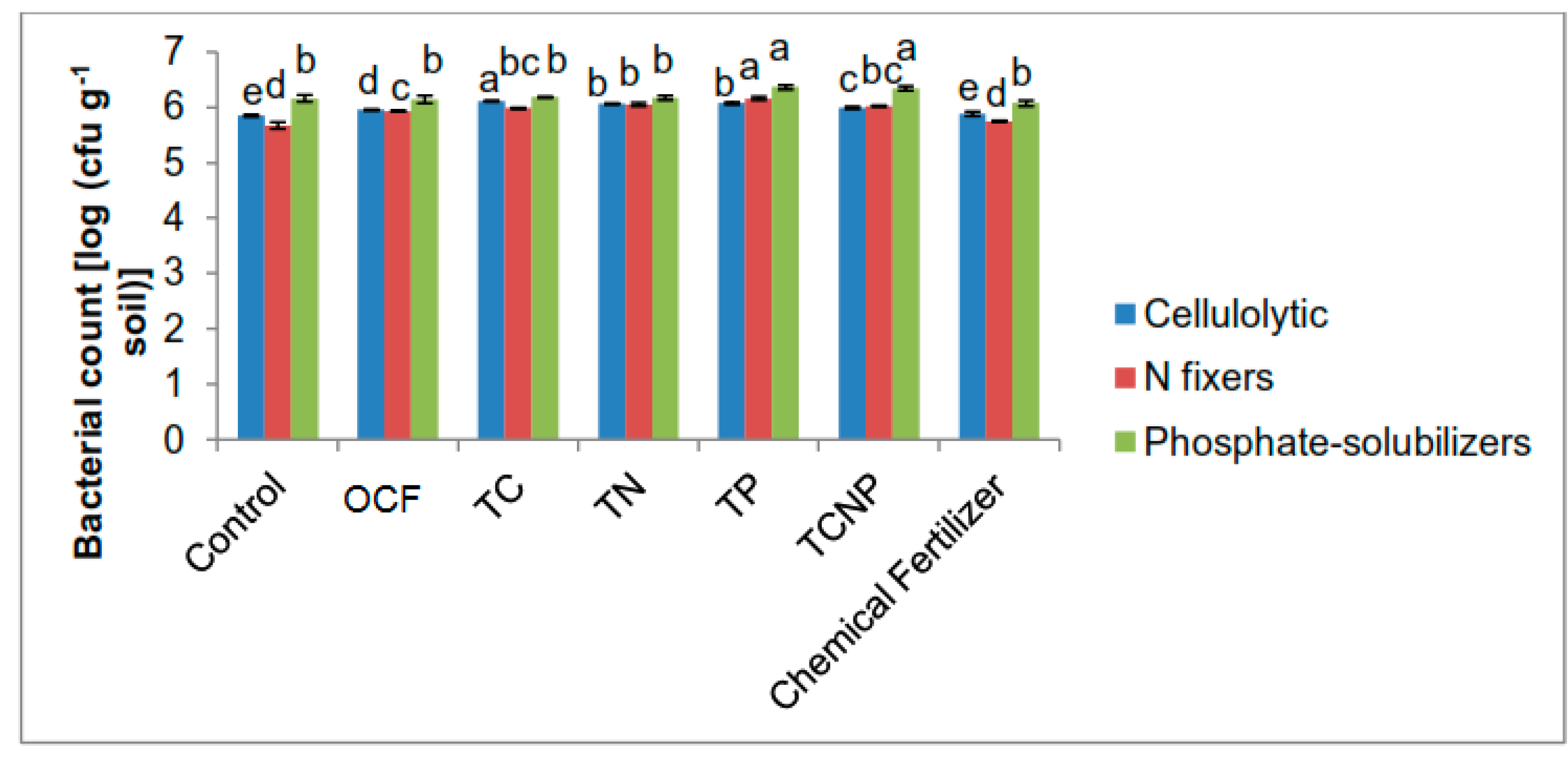
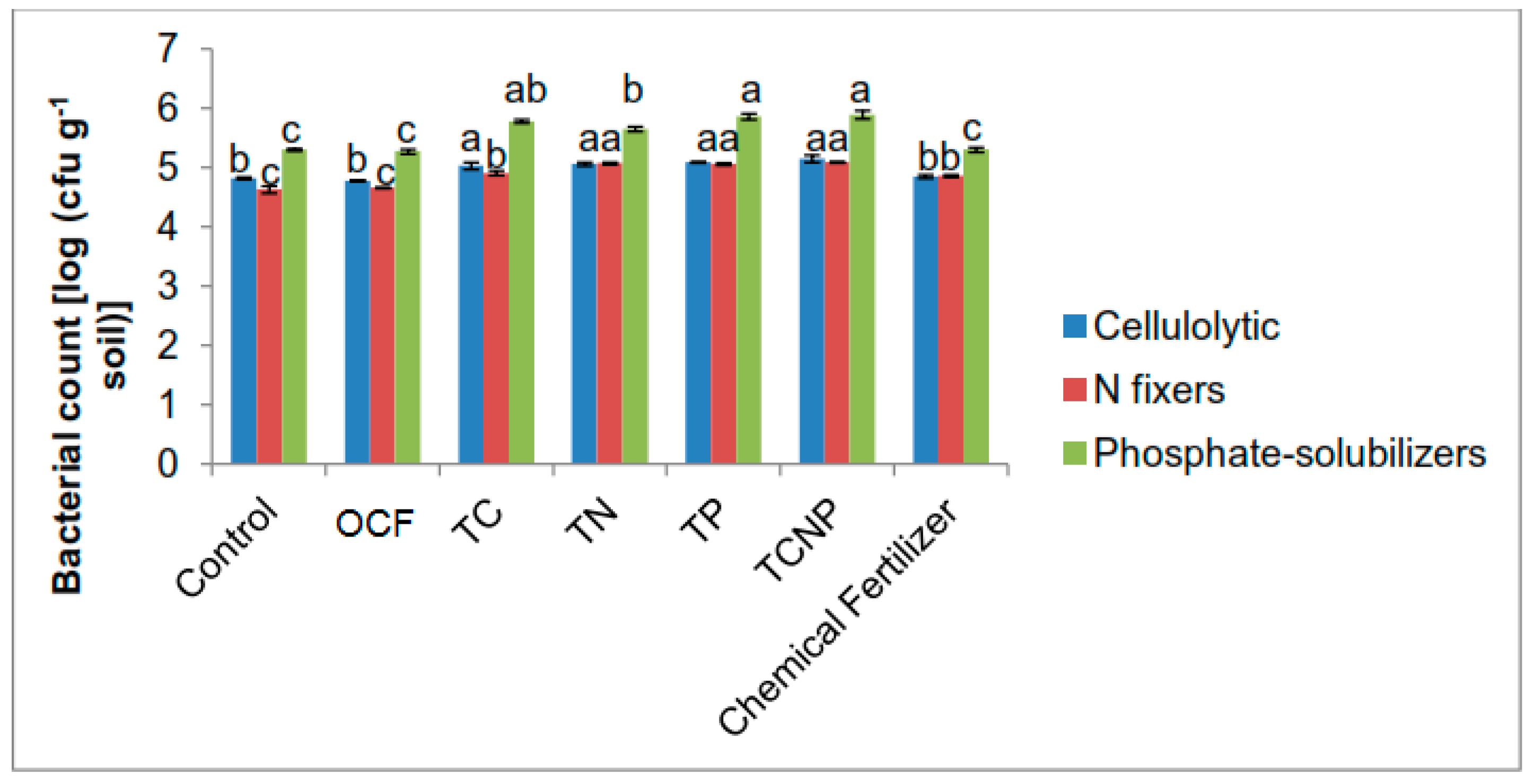
Soil samples were also collected at 56 DAS for bacterial enumeration and the aforementioned soil analyses. Enumeration of cellulolytic, N-fixing and phosphate-solubilizing bacterial populations was also done at 30 DAS and it was determined from fresh soil samples using suspension dilution techniques on the aforementioned respective selective media under
Section 2.1.
2.7. Data Analysis
Treatment effects were ascertained using analysis of variance (ANOVA) and Tukey’s Test was used for comparison of treatment means. The Statistical Analysis System (SAS) version 9.2 was used for all statistical analysis at a significance level of P ≤ 0.05.




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}





