Occurrence of tet(O/M/O) Mosaic Gene in Tetracycline-Resistant Campylobacter


Abstract
:1. Introduction
2. Materials and Methods
2.1. Microbial Growth And Antibiotic Resistance Testing
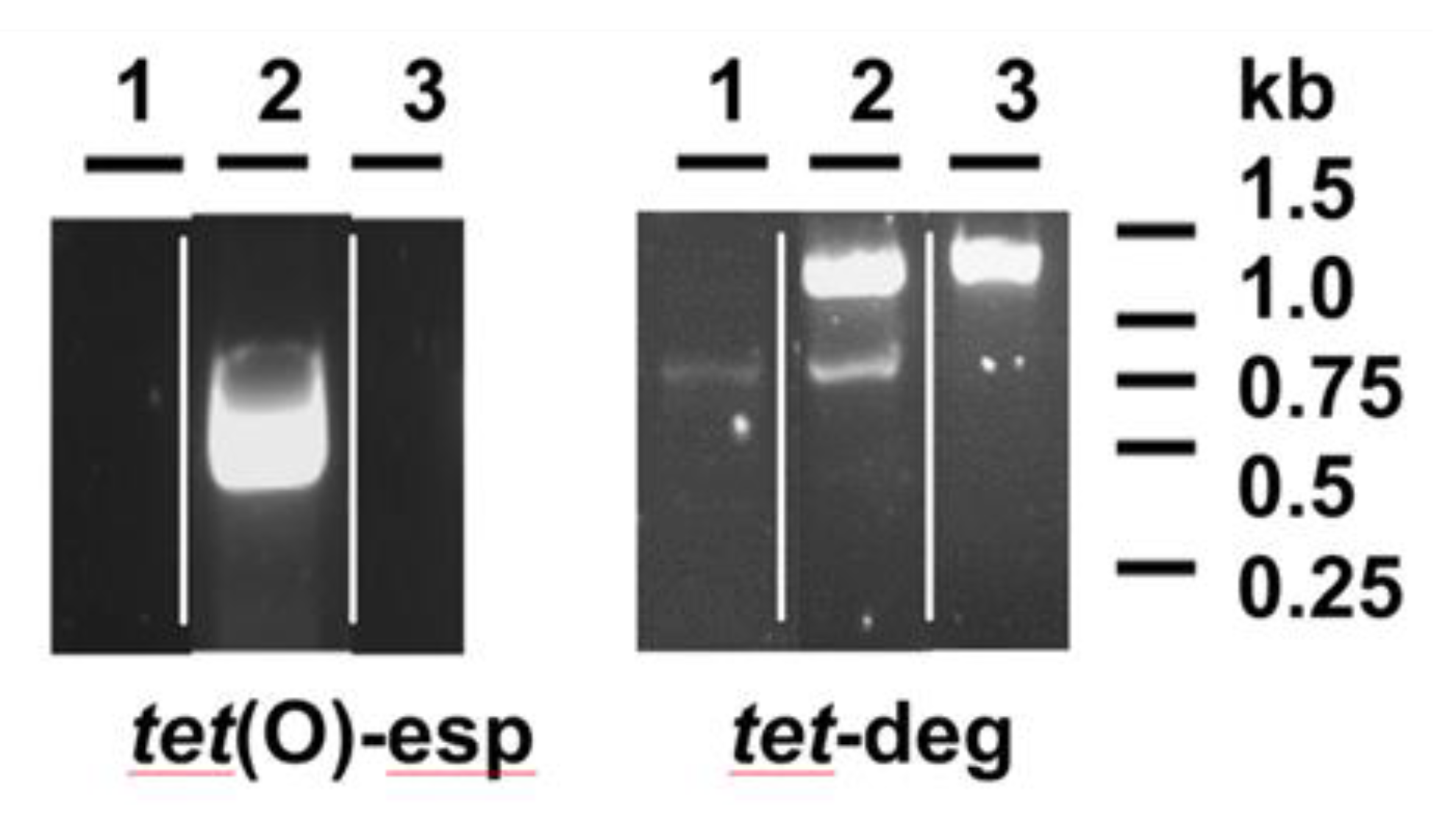
2.2. PCR Screening of Resistance Determinants
2.3. Bioinformatic Tools
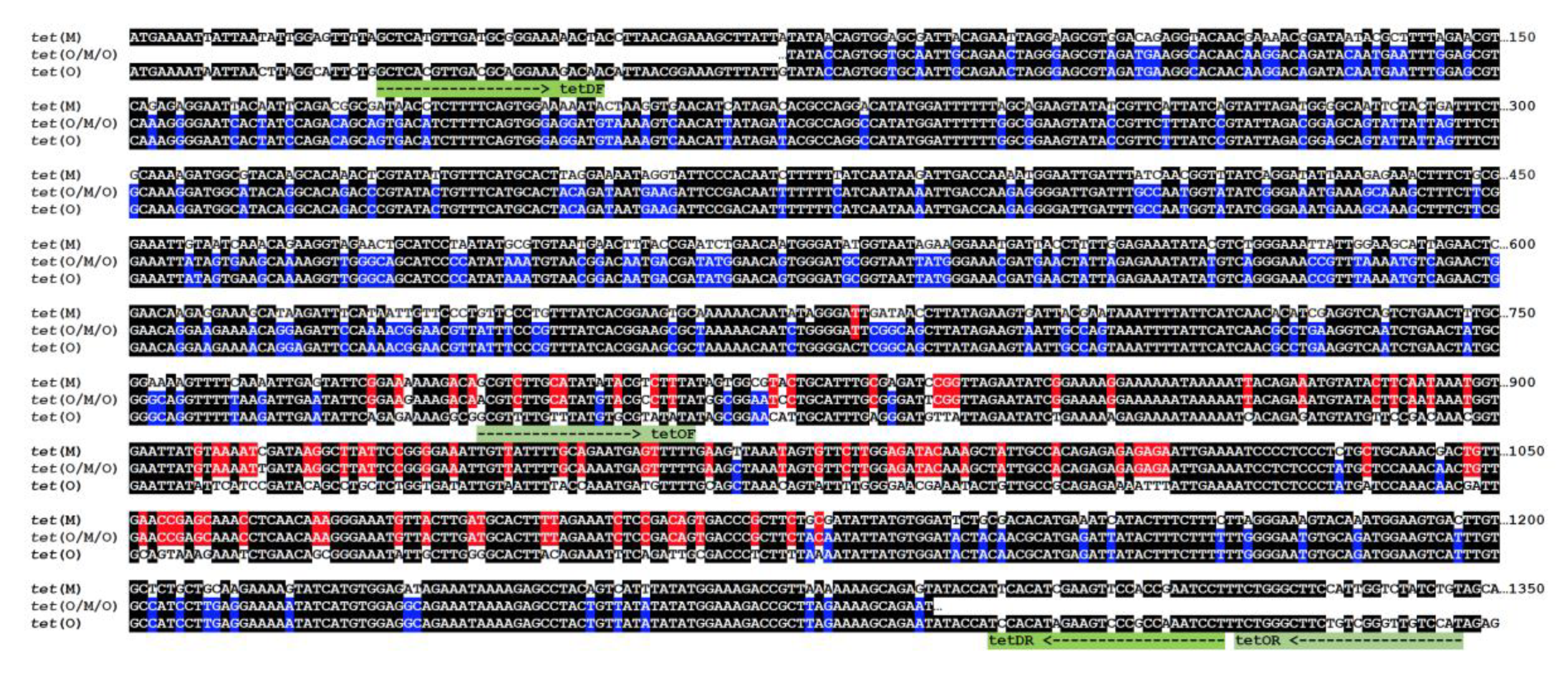
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- European Food Safety Authority, European Centre for Disease Prevention and Control. The European Union summary report on antimicrobial resistance in zoonotic and indicator bacteria from humans, animals and food in 2017. EFSA J. 2019, 17, e05598. [Google Scholar]
- Dai, L.; Sahin, O.; Grover, M.; Zhang, Q. New and alternative strategies for the prevention, control, and treatment of antibiotic-resistant Campylobacter. Transl. Res. 2020, 223, 76–88. [Google Scholar] [CrossRef]
- Arakawa, Y. Systematic research to overcome newly emerged multidrug-resistant bacteria. Microbiol. Immunol. 2020, 64, 231–251. [Google Scholar] [CrossRef]
- Tang, Y.; Fang, L.; Xu, C.; Zhang, Q. Antibiotic resistance trends and mechanisms in the foodborne pathogen, Campylobacter. Anim. Health Res. Rev. 2017, 18, 87–98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chopra, I.; Roberts, M. Tetracycline antibiotics: Mode of action, applications, molecular biology, and epidemiology of bacterial resistance. Microbiol. Mol. Biol. Rev. 2001, 65, 232–260. [Google Scholar] [CrossRef] [Green Version]
- Granados-Chinchilla, F.; Rodríguez, C. Tetracyclines in Food and Feedingstuffs: From Regulation to Analytical Methods, Bacterial Resistance, and Environmental and Health Implications. J. Anal. Methods Chem. 2017, 2017, 1315497. [Google Scholar] [CrossRef]
- Warburton, P.J.; Amodeo, N.; Roberts, A.P. Mosaic tetracycline resistance genes encoding ribosomal protection proteins. J. Antimicrob. Chemother. 2016, 71, 3333–3339. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zilhao, R.; Papadopoulou, B.; Courvalin, P. Occurrence of the Campylobacter resistance gene tetO in Enterococcus and Streptococcus spp. Antimicrob. Agents Chemother. 1988, 32, 1793–1796. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sougakoff, W.; Papadopoulou, B.; Nordmann, P.; Courvalin, P. Nucleotide sequence and distribution of gene tetO encoding tetracycline resistance in Campylobacter coli. FEMS Microbiol. Lett. 1987, 44, 153–159. [Google Scholar] [CrossRef]
- Leski, T.A.; Bangura, U.; Jimmy, D.H.; Ansumana, R.; Lizewski, S.E.; Stenger, D.A.; Taitt, C.R.; Vora, G.J. Multidrug-resistant tet(X)-containing hospital isolates in Sierra Leone. Int. J. Antimicrob. Agents 2013, 42, 83–86. [Google Scholar] [CrossRef] [PubMed]
- Marosevic, D.; Kaevska, M.; Jaglic, Z. Resistance to the tetracyclines and macrolide-lincosamide-streptogramin group of antibiotics and its genetic linkage—A review. Ann. Agric. Environ. Med. 2017, 24, 338–344. [Google Scholar] [CrossRef] [PubMed]
- Park, J.; Gasparrini, A.; Reck, M.R.; Symister, C.T.; Elliott, J.L.; Vogel, J.P.; Wencewicz, T.A.; Dantas, G.; Tolia, N.H.M. Plasticity, dynamics, and inhibition of emerging tetracycline resistance enzymes. Nat. Chem. Biol. 2017, 13, 730–736. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marilyn, C. Roberts. Available online: http://faculty.washington.edu/marilynr/ (accessed on 30 October 2020).
- Lynch, C.; Hawkins, K.; Lynch, H.; Egan, J.; Bolton, D.; Coffey, A.; Lucey, B. Investigation of molecular mechanisms underlying tetracycline resistance in thermophilic Campylobacter spp. suggests that previous reports of tet(A)-mediated resistance in these bacteria are premature. Gut Pathog. 2019, 11, 56. [Google Scholar] [CrossRef] [PubMed]
- Fabre, A.; Oleastro, M.; Nunes, A.; Santos, A.; Sifré, E.; Ducournau, A.; Bénéjat, L.; Buissonnière, A.; Floch, P.; Mégraud, F.; et al. Whole-Genome Sequence Analysis of Multidrug-Resistant Campylobacter Isolates: A Focus on Aminoglycoside Resistance Determinants. J. Clin. Microbiol. 2018, 56, e00390-18. [Google Scholar] [CrossRef] [Green Version]
- French, N.P.; Zhang, J.; Carter, G.P.; Midwinter, A.C.; Biggs, P.J.; Dyet, K.; Gilpin, B.J.; Ingle, D.J.; Mulqueen, K.; Rogers, L.E.; et al. Genomic Analysis of Fluoroquinolone- and Tetracycline-Resistant Campylobacter jejuni Sequence Type 6964 in Humans and Poultry, New Zealand, 2014–2016. Emerg. Infect. Dis. 2019, 25, 2226–2234. [Google Scholar] [CrossRef] [Green Version]
- Tenover, F.C.; Williams, S.; Gordon, K.P.; Nolan, C.; Plorde, J.J. Survey of plasmids and resistance factors in Campylobacter jejuni and Campylobacter coli. Antimicrob. Agents Chemother. 1985, 27, 37–41. [Google Scholar] [CrossRef] [Green Version]
- Crespo, M.D.; Olson, J.W.; Altermann, E.; Siletzky, R.M.; Kathariou, S. Chromosomal tet(O)-harboring regions in Campylobacter coli isolates from turkeys and swine. Appl. Environ. Microbiol. 2012, 78, 8488–8491. [Google Scholar] [CrossRef] [Green Version]
- Pratt, A.; Korolik, V. Tetracycline resistance of Australian Campylobacter jejuni and Campylobacter coli isolates. J. Antimicrob. Chemother. 2005, 55, 452–460. [Google Scholar] [CrossRef] [Green Version]
- Tang, Y.; Meinersmann, R.J.; Sahin, O.; Wu, Z.; Dai, L.; Carlson, J.; Plumblee Lawrence, J.; Genzlinger, L.; LeJeune, J.T.; Zhang, Q. Wide but Variable Distribution of a Hypervirulent Campylobacter jejuni Clone in Beef and Dairy Cattle in the United States. Appl. Environ. Microbiol. 2017, 83, e01425-17. [Google Scholar] [CrossRef] [Green Version]
- Hormeño, L.; Palomo, G.; Ugarte-Ruiz, M.; Porrero, M.C.; Borge, C.; Vadillo, S.; Píriz, S.; Domínguez, L.; Campos, M.J.; Quesada, A. Identification of the main quinolone resistance determinant in Campylobacter jejuni and Campylobacter coli by MAMA-DEG PCR. Diagn. Microbiol. Infect. Dis. 2016, 84, 236–239. [Google Scholar] [CrossRef]
- Hormeño, L.; Ugarte-Ruiz, M.; Palomo, G.; Florez-Cuadrado, D.; Vadillo, S.; Píriz, S.; Domínguez, L.; Campos, M.J.; Quesada, A. ant(6)-I Genes Encoding Aminoglycoside O-Nucleotidyltransferases Are Widely Spread Among Streptomycin Resistant Strains of Campylobacter jejuni and Campylobacter coli. Front. Microbiol. 2018, 9, 2515. [Google Scholar] [CrossRef] [Green Version]
- Mourkas, E.; Florez-Cuadrado, D.; Pascoe, B.; Calland, J.K.; Bayliss, S.C.; Mageiros, L.; Méric, G.; Hitchings, M.D.; Quesada, A.; Porrero, C.; et al. Gene pool transmission of multidrug resistance among Campylobacter from livestock, sewage and human disease. Environ. Microbiol. 2019, 21, 4597–4613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clinical and Laboratory Standard Institute (CLSI). Methods for Antimicrobial Dilution and Disk Susceptibility Testing of Infrequently Isolated or Fastidious Bacteria, 3rd ed.; CLSI guideline M45; Clinical and Laboratory Standard Institute: Wayne, PA, USA, 2016. [Google Scholar]
- Bacon, D.J.; Alm, R.A.; Burr, D.H.; Hu, L.; Kopecko, D.J.; Ewing, C.P.; Trust, T.J.; Guerry, P. Involvement of a plasmid in virulence of Campylobacter jejuni 81-176. Infect. Immun. 2000, 68, 4384–4390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barbosa, T.M.; Scott, K.P.; Flint, H.J. Evidence for recent intergeneric transfer of a new tetracycline resistance gene, tet(W), isolated from Butyrivibrio fibrisolvens, and the occurrence of tet(O) in ruminal bacteria. Environ. Microbiol. 1999, 1, 53–64. [Google Scholar] [CrossRef] [PubMed]
- National Center for Biotechnology Information, U.S. National Library of Medicine 8600 Rockville Pike, Bethesda MD, 20894 USA.
- Lopes, B.S.; Strachan, N.; Ramjee, M.; Thomson, A.; MacRae, M.; Shaw, S.; Forbes, K.J. Nationwide Stepwise Emergence and Evolution of Multidrug-Resistant Campylobacter jejuni Sequence Type 5136, United Kingdom. Emerg. Infect. Dis. 2019, 25, 1320–1329. [Google Scholar] [CrossRef] [Green Version]
- Dönhöfer, A.; Franckenberg, S.; Wickles, S.; Berninghausen, O.; Beckmann, R.; Wilson, D.N. Structural basis for TetM-mediated tetracycline resistance. Proc. Natl. Acad. Sci. USA 2012, 109, 16900–16905. [Google Scholar] [CrossRef] [Green Version]
- Elhadidy, M.; Ali, M.M.; El-Shibiny, A.; Miller, W.G.; Elkhatib, W.F.; Botteldoorn, N.; Dierick, K. Antimicrobial resistance patterns and molecular resistance markers of Campylobacter jejuni isolates from human diarrheal cases. PLoS ONE 2020, 15, e0227833. [Google Scholar] [CrossRef] [Green Version]
- Marotta, F.; Garofolo, G.; di Marcantonio, L.; di Serafino, G.; Neri, D.; Romantini, R.; Sacchini, L.; Alessiani, A.; Di Donato, G.; Nuvoloni, R.; et al. Antimicrobial resistance genotypes and phenotypes of Campylobacter jejuni isolated in Italy from humans, birds from wild and urban habitats, and poultry. PLoS ONE 2019, 14, e0223804. [Google Scholar] [CrossRef] [Green Version]
- European Committee on Antimicrobial Susceptibility Testing. Available online: https://eucast.org (accessed on 30 October 2020).
- Ocejo, M.; Oporto, B.; Hurtado, A. Occurrence of Campylobacter jejuni and Campylobacter coli in Cattle and Sheep in Northern Spain and Changes in Antimicrobial Resistance in Two Studies 10-years Apart. Pathogens 2019, 8, 98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- European Medicines Agency (EMA). European Surveillance of Veterinary Antimicrobial Consumption. Sales of Veterinary Antimicrobial Agents in 30 European Countries in 2015 (EMA/387934/2015); European Medicines Agency: Amsterdam, The Netherlands, 2018. [Google Scholar]
- Marasini, D.; Karki, A.B.; Buchheim, M.A.; Fakhr, M.K. Phylogenetic Relatedness Among Plasmids Harbored by Campylobacter jejuni and Campylobacter coli Isolated From Retail Meats. Front. Microbiol. 2018, 9, 2167. [Google Scholar] [CrossRef] [Green Version]
- Spigaglia, P.; Barbanti, F.; Mastrantonio, P. Tetracycline resistance gene tet(W) in the pathogenic bacterium Clostridium difficile. Antimicrob. Agents Chemother. 2008, 52, 770–773. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stanton, T.B.; Humphrey, S.B. Isolation of tetracycline-resistant Megasphaera elsdenii strains with novel mosaic gene combinations of tet(O) and tet(W) from swine. Appl. Environ. Microbiol. 2003, 69, 3874–3882. [Google Scholar] [CrossRef] [Green Version]
- Batchelor, R.A.; Pearson, B.M.; Friis, L.M.; Guerry, P.; Wells, J.M. Nucleotide sequences and comparison of two large conjugative plasmids from different Campylobacter species. Microbiology 2004, 150, 3507–3517. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Speer, B.S.; Shoemaker, N.B.; Salyers, A.A. Bacterial resistance to tetracycline: Mechanisms, transfer, and clinical significance. Clin. Microbiol. Rev. 1992, 5, 387–399. [Google Scholar] [CrossRef]
- Yu, A.M.; Choi, Y.H.; Tu, M.J. RNA Drugs and RNA Targets for Small Molecules: Principles, Progress, and Challenges. Pharmacol. Rev. 2020, 72, 862–898. [Google Scholar] [CrossRef]
- Li, W.; Atkinson, G.C.; Thakor, N.S.; Allas, U.; Lu, C.C.; Chan, K.Y.; Tenson, T.; Schulten, K.; Wilson, K.S.; Hauryliuk, V.; et al. Mechanism of tetracycline resistance by ribosomal protection protein Tet(O). Nat. Commun. 2013, 4, 1477. [Google Scholar] [CrossRef] [Green Version]
- Thaker, M.; Spanogiannopoulos, P.; Wright, G.D. The tetracycline resistome. Cell. Mol. Life Sci. 2010, 67, 419–431. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| MIC # (mg/L) | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Species | Tet(O) * | ≤4 | 8 | 16 | 32 | 64 | 128 | 256 | ≥512 | Sum |
| C. jejuni | + | 0 | 0 | 3 | 3 | 24 | 50 | 9 | 14 | 103 |
| - | 26 | 2 | 0 | 0 | 0 | 0 | 0 | 1 | 29 | |
| C. coli | + | 0 | 0 | 0 | 1 | 1 | 0 | 3 | 1 | 6 |
| - | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hormeño, L.; Campos, M.J.; Vadillo, S.; Quesada, A. Occurrence of tet(O/M/O) Mosaic Gene in Tetracycline-Resistant Campylobacter. Microorganisms 2020, 8, 1710. https://doi.org/10.3390/microorganisms8111710
Hormeño L, Campos MJ, Vadillo S, Quesada A. Occurrence of tet(O/M/O) Mosaic Gene in Tetracycline-Resistant Campylobacter. Microorganisms. 2020; 8(11):1710. https://doi.org/10.3390/microorganisms8111710
Chicago/Turabian StyleHormeño, Lorena, Maria J. Campos, Santiago Vadillo, and Alberto Quesada. 2020. "Occurrence of tet(O/M/O) Mosaic Gene in Tetracycline-Resistant Campylobacter" Microorganisms 8, no. 11: 1710. https://doi.org/10.3390/microorganisms8111710
APA StyleHormeño, L., Campos, M. J., Vadillo, S., & Quesada, A. (2020). Occurrence of tet(O/M/O) Mosaic Gene in Tetracycline-Resistant Campylobacter. Microorganisms, 8(11), 1710. https://doi.org/10.3390/microorganisms8111710