Functional Properties and Sustainability Improvement of Sourdough Bread by Lactic Acid Bacteria


Abstract
:1. Introduction
2. Materials and Methods
2.1. Matrices, Producer, and Indicator Strains
2.2. Carbohydrate Metabolism
2.3. Determination of Antifungal Activity by the Dual Culture Overlay Assay
2.4. Determination of Antibacillus Activity by Spot-on-the-Lawn Technique
2.5. Screening of LAB Strains for the Presence of Glutamic Acid Decarboxylase Gene
2.6. Lactic Acid Bacteria Strain Differentiation
2.7. Statistical Analysis
3. Results and Discussion
3.1. Strain Differentiation
3.2. Carbohydrate Metabolism
3.3. Antifungal Activity
3.4. Antibacilli Activity
3.5. Determination of the GAD Gene
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Asioli, D.; Aschemann-Witzel, J.; Caputo, V.; Vecchio, R.; Annunziata, A.; Naes, T.; Varela, P. Making sense of the “clean label” trends: A review of consumer food choice behavior and discussion of industry implications. Food Res. Int. 2017, 99, 58–71. [Google Scholar] [CrossRef]
- Ricci, A.; Bernini, V.; Maoloni, A.; Cirlini, M.; Galaverna, G.; Neviani, E.; Lazzi, C. Vegetable by-product lacto-fermentation as a new source of antimicrobial compounds. Microorganisms 2019, 7, 607. [Google Scholar] [CrossRef] [Green Version]
- Gänzle, M.G.; Zheng, J. Lifestyles of sourdough lactobacilli - Do they matter for microbial ecology and bread quality? Int. J. Food Microbiol. 2019, 302, 15–23. [Google Scholar] [CrossRef] [PubMed]
- Gobbetti, M. The sourdough microflora: Interactions of lactic acid bacteria and yeasts. Trends Food Sci. Technol. 1998, 9, 267–274. [Google Scholar] [CrossRef]
- Corsetti, A.; Settanni, L. Lactobacilli in sourdough fermentation. Food Res. Int. 2007, 40, 539–558. [Google Scholar] [CrossRef]
- Brandt, M.J.; Gänzle, M. Handbuch Sauerteig; Behr: Hamburg, Germany, 2006. [Google Scholar]
- Chen, C.C.; Lai, C.C.; Huang, H.L.; Huang, W.Y.; Toh, H.S.; Weng, T.C.; Chuang, Y.C.; Lu, Y.C.; Tang, H.J. Antimicrobial activity of Lactobacillus species against Carbapenem-resistant Enterobacteriaceae. Front. Microbiol. 2019, 10, 789. [Google Scholar] [CrossRef] [PubMed]
- Quattrini, M.; Liang, N.; Fortina, M.G.; Xiang, S.; Curtis, J.M.; Ganzle, M. Exploiting synergies of sourdough and antifungal organic acids to delay fungal spoilage of bread. Int. J. Food Microbiol. 2019, 302, 8–14. [Google Scholar] [CrossRef] [PubMed]
- Rosenkvist, H.; Hansen, A. Contamination profiles and characterisation of Bacillus species in wheat bread and raw materials for bread production. Int. J. Food Microbiol. 1995, 26, 353–363. [Google Scholar] [CrossRef]
- Mantzourani, I.; Plessas, S.; Saxami, G.; Alexopoulos, A.; Galanis, A.; Bezirtzoglou, E. Study of kefir grains application in sourdough bread regarding rope spoilage caused by Bacillus spp. Food Chem. 2014, 143, 17–21. [Google Scholar] [CrossRef]
- Şimşek, Ö.; Çon, A.H.; Tulumogˇlu, Ş. Isolating lactic starter cultures with antimicrobial activity for sourdough processes. Food Control. 2006, 17, 263–270. [Google Scholar] [CrossRef]
- Garofalo, C.; Zannini, E.; Aquilanti, L.; Silvestri, G.; Fierro, O.; Picariello, G.; Clementi, F. Selection of sourdough lactobacilli with antifungal activity for use as biopreservatives in bakery products. J. Agric. Food Chem. 2012, 60, 7719–7728. [Google Scholar] [CrossRef]
- Sadeghi, A.; Ebrahimi, M.; Mortazavi, S.A.; Abedfar, A. Application of the selected antifungal LAB isolate as a protective starter culture in pan whole-wheat sourdough bread. Food Control. 2019, 95, 298–307. [Google Scholar] [CrossRef]
- Sadeghi, A.; Ebrahimi, M.; Raeisi, M.; Nematollahi, Z. Biological control of foodborne pathogens and aflatoxins by selected probiotic LAB isolated from rice bran sourdough. Biol. Control. 2019, 130, 70–79. [Google Scholar] [CrossRef]
- Fekri, A.; Torbati, M.; Yari Khosrowshahi, A.; Bagherpour Shamloo, H.; Azadmard-Damirchi, S. Functional effects of phytate-degrading, probiotic lactic acid bacteria and yeast strains isolated from Iranian traditional sourdough on the technological and nutritional properties of whole wheat bread. Food Chem. 2020, 306, 125620. [Google Scholar] [CrossRef] [PubMed]
- Reale, A.; Di Renzo, T.; Succi, M.; Tremonte, P.; Coppola, R.; Sorrentino, E. Microbiological and fermentative properties of baker’s yeast starter used in breadmaking. J. Food Sci. 2013, 78, M1224–M1231. [Google Scholar] [CrossRef]
- Menteş, Ö.; Ercan, R.; Akçelik, M. Inhibitor activities of two Lactobacillus strains, isolated from sourdough, against rope-forming Bacillus strains. Food Control. 2007, 18, 359–363. [Google Scholar] [CrossRef]
- Valerio, F.; De Bellis, P.; Di Biase, M.; Lonigro, S.L.; Giussani, B.; Visconti, A.; Lavermicocca, P.; Sisto, A. Diversity of spore-forming bacteria and identification of Bacillus amyloliquefaciens as a species frequently associated with the ropy spoilage of bread. Int. J. Food Microbiol. 2012, 156, 278–285. [Google Scholar] [CrossRef]
- Garcia, M.V.; Bernardi, A.O.; Copetti, M.V. The fungal problem in bread production: Insights of causes, consequences, and control methods. Curr. Opin. Food Sci. 2019, 29, 1–6. [Google Scholar] [CrossRef]
- Legan, J.D. Mould spoilage of bread: The problem and some solutions. Int. Biodeterior. Biodegrad. 1993, 32, 33–53. [Google Scholar] [CrossRef]
- Bryden, W.L. Mycotoxins in the food chain: Human health implications. Asia Pac. J. Clin. Nutr. 2007, 16, 95–101. [Google Scholar]
- Siragusa, S.; De Angelis, M.; Di Cagno, R.; Rizzello, C.G.; Coda, R.; Gobbetti, M. Synthesis of gamma-aminobutyric acid by lactic acid bacteria isolated from a variety of Italian cheeses. Appl. Environ. Microbiol. 2007, 73, 7283–7290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, Q.; Tun, H.M.; Law, Y.S.; Khafipour, E.; Shah, N.P. Common distribution of gad operon in Lactobacillus brevis and its GadA contributes to efficient GABA synthesis toward cytosolic near-neutral pH. Front. Microbiol. 2017, 8, 206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diana, M.; Quílez, J.; Rafecas, M. Gamma-aminobutyric acid as a bioactive compound in foods: A review. J. Funct. Foods 2014, 10, 407–420. [Google Scholar] [CrossRef]
- Koubaa, M.; Delbecq, F.; Roohinejad, S.; Mallikarjunan, K. Gamma-Aminobutyric Acid. In Encyclopedia of Food Chemistry; Melton, L., Shahidi, F., Varelis, P., Eds.; Academic Press: Oxford, UK, 2019; pp. 528–534. [Google Scholar] [CrossRef]
- Yilmaz, C.; Gokmen, V. Neuroactive compounds in foods: Occurrence, mechanism and potential health effects. Food Res. Int. 2020, 128, 108744. [Google Scholar] [CrossRef] [PubMed]
- Venturi, M.; Galli, V.; Pini, N.; Guerrini, S.; Granchi, L. Use of selected lactobacilli to increase gamma-aminobutyric acid (GABA) content in sourdough bread enriched with amaranth flour. Foods 2019, 8. [Google Scholar] [CrossRef] [Green Version]
- Bartkiene, E.; Bartkevics, V.; Mozuriene, E.; Lele, V.; Zadeike, D.; Juodeikiene, G. The safety, technological, nutritional, and sensory challenges associated with lacto-fermentation of meat and meat products by using pure lactic acid bacteria strains and plant-lactic acid bacteria bioproducts. Front. Microbiol. 2019, 10, 1036. [Google Scholar] [CrossRef]
- Dimidi, E.; Cox, S.R.; Rossi, M.; Whelan, K. Fermented foods: Definitions and characteristics, impact on the gut microbiota and effects on gastrointestinal health and disease. Nutrients 2019, 11. [Google Scholar] [CrossRef] [Green Version]
- Holzapfel, W.H.; Wood, B.J.B. Lactic Acid Bacteria: Biodiversity and Taxonomy; Wiley: Hoboken, NJ, USA, 2014. [Google Scholar]
- Fraberger, V.; Unger, C.; Kummer, C.; Domig, K.J. Insights into microbial diversity of traditional Austrian sourdough. LWT Food Sci. Technol. 2020, 127. [Google Scholar] [CrossRef]
- Call, L.-M.; D’Amico, S.; Grausgruber, H.; Schönlechner, R. Fruktane in alten und neuen österreichischen Weizensorten. Getreide Mehl Und Brot 2018, 2018, 2–6. [Google Scholar]
- Magnusson, J.; Schnurer, J. Lactobacillus coryniformis subsp. coryniformis strain Si3 produces a broad-spectrum proteinaceous antifungal compound. Appl. Environ. Microbiol. 2001, 67, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Manini, F.; Casiraghi, M.C.; Poutanen, K.; Brasca, M.; Erba, D.; Plumed-Ferrer, C. Characterization of lactic acid bacteria isolated from wheat bran sourdough. LWT Food Sci. Technol. 2016, 66, 275–283. [Google Scholar] [CrossRef]
- Stiles, J.; Penkar, S.; Plockova, M.; Chumchalova, J.; Bullerman, L.B. Antifungal activity of sodium acetate and Lactobacillus rhamnosus. J. Food Prot. 2002, 65, 1188–1191. [Google Scholar] [CrossRef]
- Tremonte, P.; Pannella, G.; Succi, M.; Tipaldi, L.; Sturchio, M.; Coppola, R.; Luongo, D.; Sorrentino, E. Antimicrobial activity of Lactobacillus plantarum strains isolated from different environments: A preliminary study. Int. Food Res. J. 2017, 24, 852–859. [Google Scholar]
- Demirbaş, F.; İspirli, H.; Kurnaz, A.A.; Yilmaz, M.T.; Dertli, E. Antimicrobial and functional properties of lactic acid bacteria isolated from sourdoughs. LWT Food Sci. Technol. 2017, 79, 361–366. [Google Scholar] [CrossRef]
- Svec, P.; Vancanneyt, M.; Seman, M.; Snauwaert, C.; Lefebvre, K.; Sedlacek, I.; Swings, J. Evaluation of (GTG)5-PCR for identification of Enterococcus spp. FEMS Microbiol. Lett. 2005, 247, 59–63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gevers, D.; Huys, G.; Swings, J. Applicability of rep-PCR fingerprinting for identification of Lactobacillus species. FEMS Microbiol. Lett. 2001, 205, 31–36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watson, D.; O’Connell Motherway, M.; Schoterman, M.H.; van Neerven, R.J.; Nauta, A.; van Sinderen, D. Selective carbohydrate utilization by lactobacilli and bifidobacteria. J. Appl. Microbiol. 2013, 114, 1132–1146. [Google Scholar] [CrossRef]
- Ehrmann, M.A.; Vogel, R.F. Molecular taxonomy and genetics of sourdough lactic acid bacteria. Trends Food Sci. Technol. 2005, 16, 31–42. [Google Scholar] [CrossRef]
- Fraberger, V.; Call, L.M.; Domig, K.J.; D’Amico, S. Applicability of yeast fermentation to reduce fructans and other FODMAPs. Nutrients 2018, 10, 1247. [Google Scholar] [CrossRef] [Green Version]
- Bryla, M.; Roszko, M.; Szymczyk, K.; Jedrzejczak, R.; Slowik, E.; Obiedzinski, M.W. Effect of baking on reduction of free and hidden fumonisins in gluten-free bread. J. Agric. Food Chem. 2014, 62, 10341–10347. [Google Scholar] [CrossRef]
- Coda, R.; Cassone, A.; Rizzello, C.G.; Nionelli, L.; Cardinali, G.; Gobbetti, M. Antifungal activity of Wickerhamomyces anomalus and Lactobacillus plantarum during sourdough fermentation: Identification of novel compounds and long-term effect during storage of wheat bread. Appl. Environ. Microbiol. 2011, 77, 3484–3492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gerez, C.L.; Torino, M.I.; Rollán, G.; Font de Valdez, G. Prevention of bread mould spoilage by using lactic acid bacteria with antifungal properties. Food Control. 2009, 20, 144–148. [Google Scholar] [CrossRef]
- Axel, C.; Brosnan, B.; Zannini, E.; Peyer, L.C.; Furey, A.; Coffey, A.; Arendt, E.K. Antifungal activities of three different Lactobacillus species and their production of antifungal carboxylic acids in wheat sourdough. Appl. Microbiol. Biotechnol. 2016, 100, 1701–1711. [Google Scholar] [CrossRef]
- Belz, M.C.; Mairinger, R.; Zannini, E.; Ryan, L.A.; Cashman, K.D.; Arendt, E.K. The effect of sourdough and calcium propionate on the microbial shelf-life of salt reduced bread. Appl Microbiol. Biotechnol. 2012, 96, 493–501. [Google Scholar] [CrossRef] [PubMed]
- Schwenninger, S.M.; von Ah, U.; Niederer, B.; Teuber, M.; Meile, L. Detection of antifungal properties in Lactobacillus paracasei subsp. paracasei SM20, SM29, and SM63 and molecular typing of the strains. J. Food Prot. 2005, 68, 111–119. [Google Scholar] [CrossRef]
- Magnusson, J.; Ström, K.; Roos, S.; Sjögren, J.r.; Schnürer, J. Broad and complex antifungal activity among environmental isolates of lactic acid bacteria. FEMS Microbiol. Lett. 2003, 219, 129–135. [Google Scholar] [CrossRef] [Green Version]
- Lavermicocca, P.; Valerio, F.; Evidente, A.; Lazzaroni, S.; Corsetti, A.; Gobbetti, M. Purification and characterization of novel antifungal compounds from the sourdough Lactobacillus plantarum strain 21B. Appl. Environ. Microbiol. 2000, 66, 4084–4090. [Google Scholar] [CrossRef] [Green Version]
- Muhialdin, B.J.; Algboory, H.L.; Kadum, H.; Mohammed, N.K.; Saari, N.; Hassan, Z.; Meor Hussin, A.S. Antifungal activity determination for the peptides generated by Lactobacillus plantarum TE10 against Aspergillus flavus in maize seeds. Food Control. 2020, 109. [Google Scholar] [CrossRef]
- Russo, P.; Arena, M.P.; Fiocco, D.; Capozzi, V.; Drider, D.; Spano, G. Lactobacillus plantarum with broad antifungal activity: A promising approach to increase safety and shelf-life of cereal-based products. Int. J. Food Microbiol. 2017, 247, 48–54. [Google Scholar] [CrossRef]
- Bartkiene, E.; Zavistanaviciute, P.; Lele, V.; Ruzauskas, M.; Bartkevics, V.; Bernatoniene, J.; Gallo, P.; Tenore, G.C.; Santini, A. Lactobacillus plantarum LUHS135 and paracasei LUHS244 as functional starter cultures for the food fermentation industry: Characterisation, mycotoxin-reducing properties, optimisation of biomass growth and sustainable encapsulation by using dairy by-products. LWT Food Sci. Technol. 2018, 93, 649–658. [Google Scholar] [CrossRef]
- Hassan, Y.I.; Bullerman, L.B. Antifungal activity of Lactobacillus paracasei subsp. tolerans against Fusarium proliferatum and Fusarium graminearum in a liquid culture setting. J. Food Prot. 2008, 71, 2213–2216. [Google Scholar] [CrossRef]
- Gajbhiye, M.; Kapadnis, B. Bio-efficiency of antifungal lactic acid bacterial isolates for pomegranate fruit rot management. Proc. Natl. Acad. Sci. India Sect. B Biol. Sci. 2017, 88, 1477–1488. [Google Scholar] [CrossRef]
- Schnürer, J.; Magnusson, J. Antifungal lactic acid bacteria as biopreservatives. Trends Food Sci. Technol. 2005, 16, 70–78. [Google Scholar] [CrossRef]
- Pepe, O.; Blaiotta, G.; Moschetti, G.; Greco, T.; Villani, F. Rope-producing strains of Bacillus spp. from wheat bread and strategy for their control by lactic acid bacteria. Appl. Environ. Microbiol. 2003, 69, 2321–2329. [Google Scholar] [CrossRef] [Green Version]
- Arena, M.P.; Silvain, A.; Normanno, G.; Grieco, F.; Drider, D.; Spano, G.; Fiocco, D. Use of Lactobacillus plantarum Strains as a Bio-Control Strategy against Food-Borne Pathogenic Microorganisms. Front. Microbiol. 2016, 7, 464. [Google Scholar] [CrossRef] [Green Version]
- Mantzourani, I.; Plessas, S.; Odatzidou, M.; Alexopoulos, A.; Galanis, A.; Bezirtzoglou, E.; Bekatorou, A. Effect of a novel Lactobacillus paracasei starter on sourdough bread quality. Food Chem. 2019, 271, 259–265. [Google Scholar] [CrossRef]
- Digaitiene, A.; Hansen, A.S.; Juodeikiene, G.; Eidukonyte, D.; Josephsen, J. Lactic acid bacteria isolated from rye sourdoughs produce bacteriocin-like inhibitory substances active against Bacillus subtilis and fungi. J. Appl. Microbiol. 2012, 112, 732–742. [Google Scholar] [CrossRef]
- Villegas, J.M.; Brown, L.; de Giori, G.S.; Hebert, E.M. Optimization of batch culture conditions for GABA production by Lactobacillus brevis CRL 1942, isolated from quinoa sourdough. LWT Food Sci. Technol. 2016, 67, 22–26. [Google Scholar] [CrossRef]


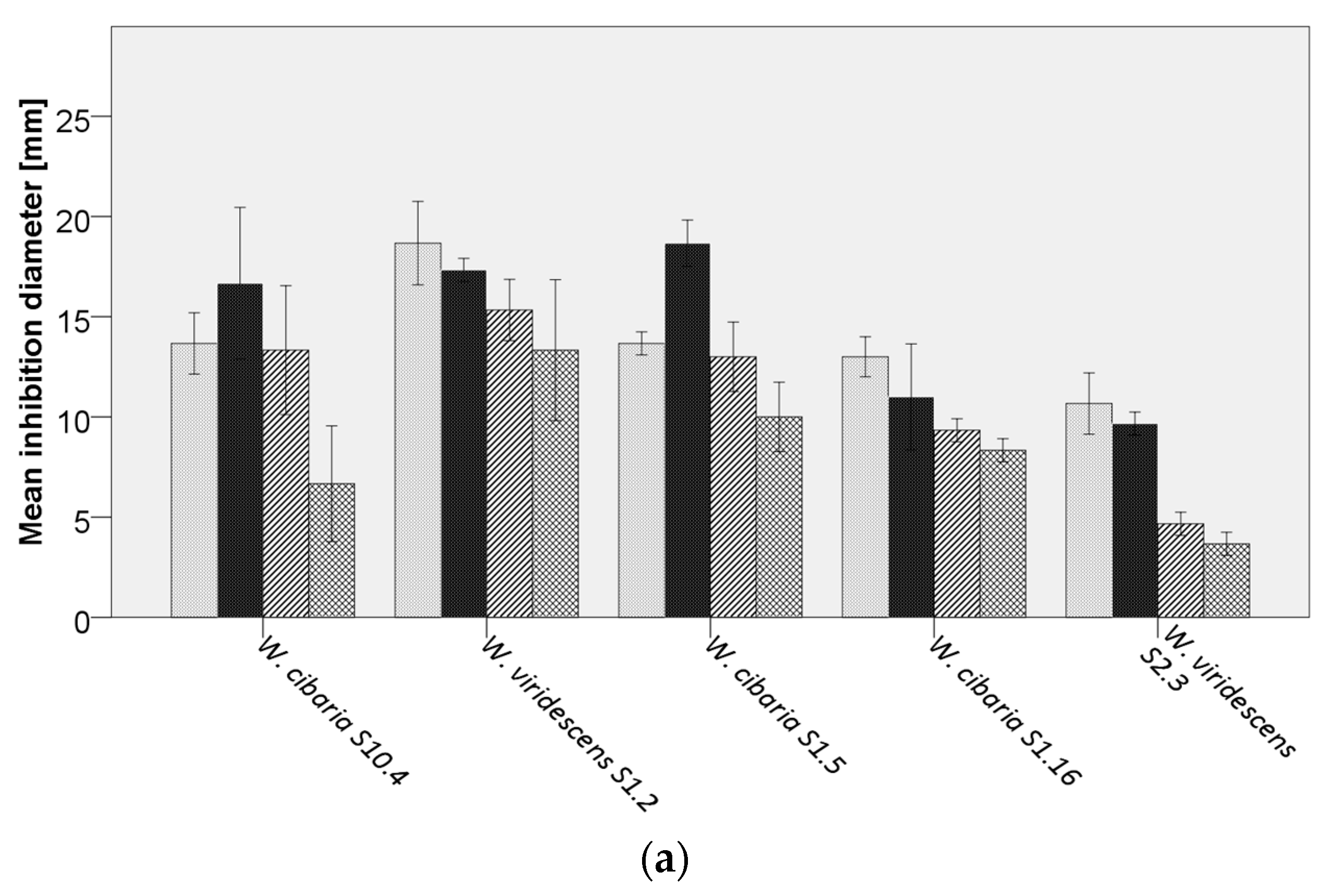
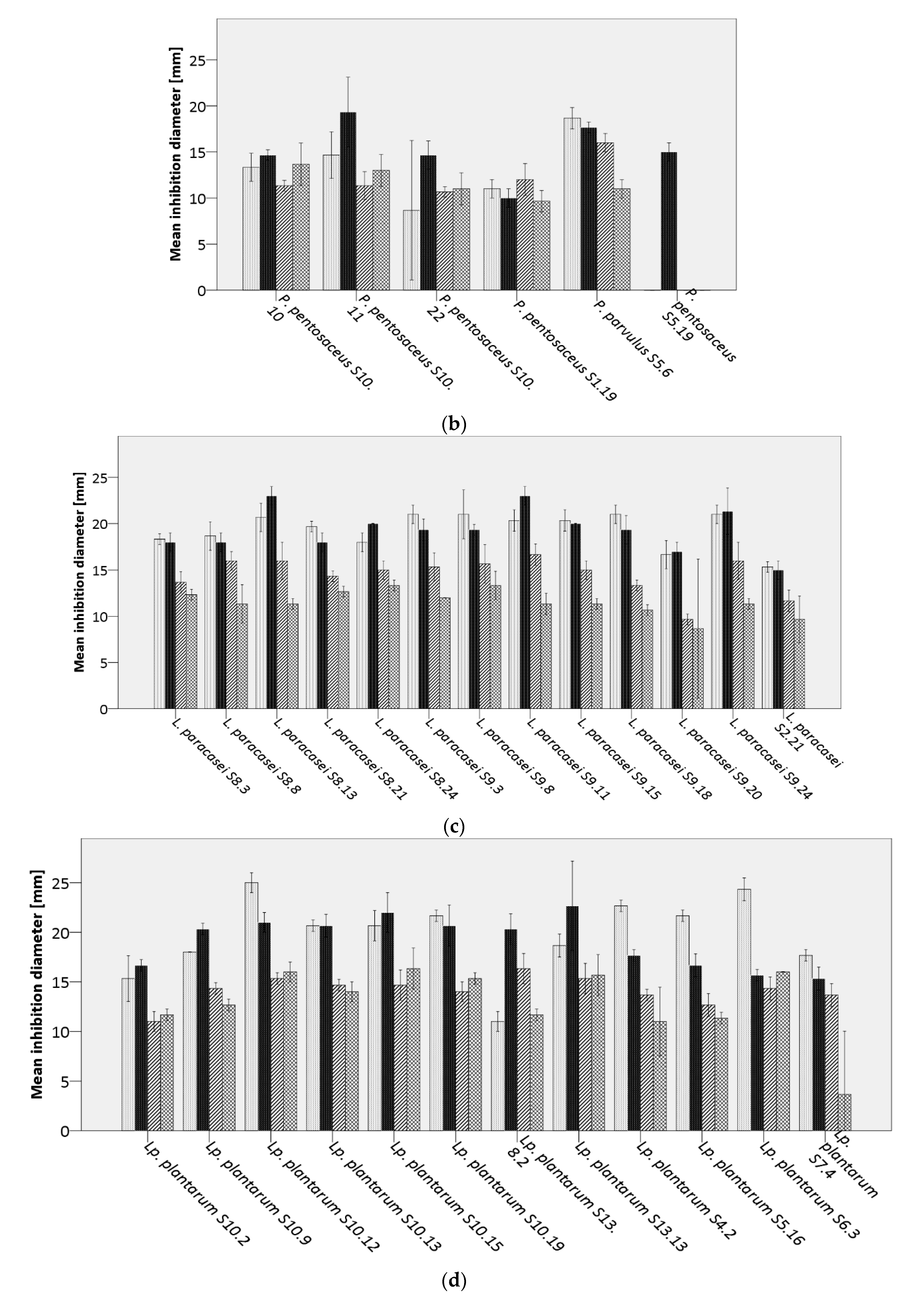
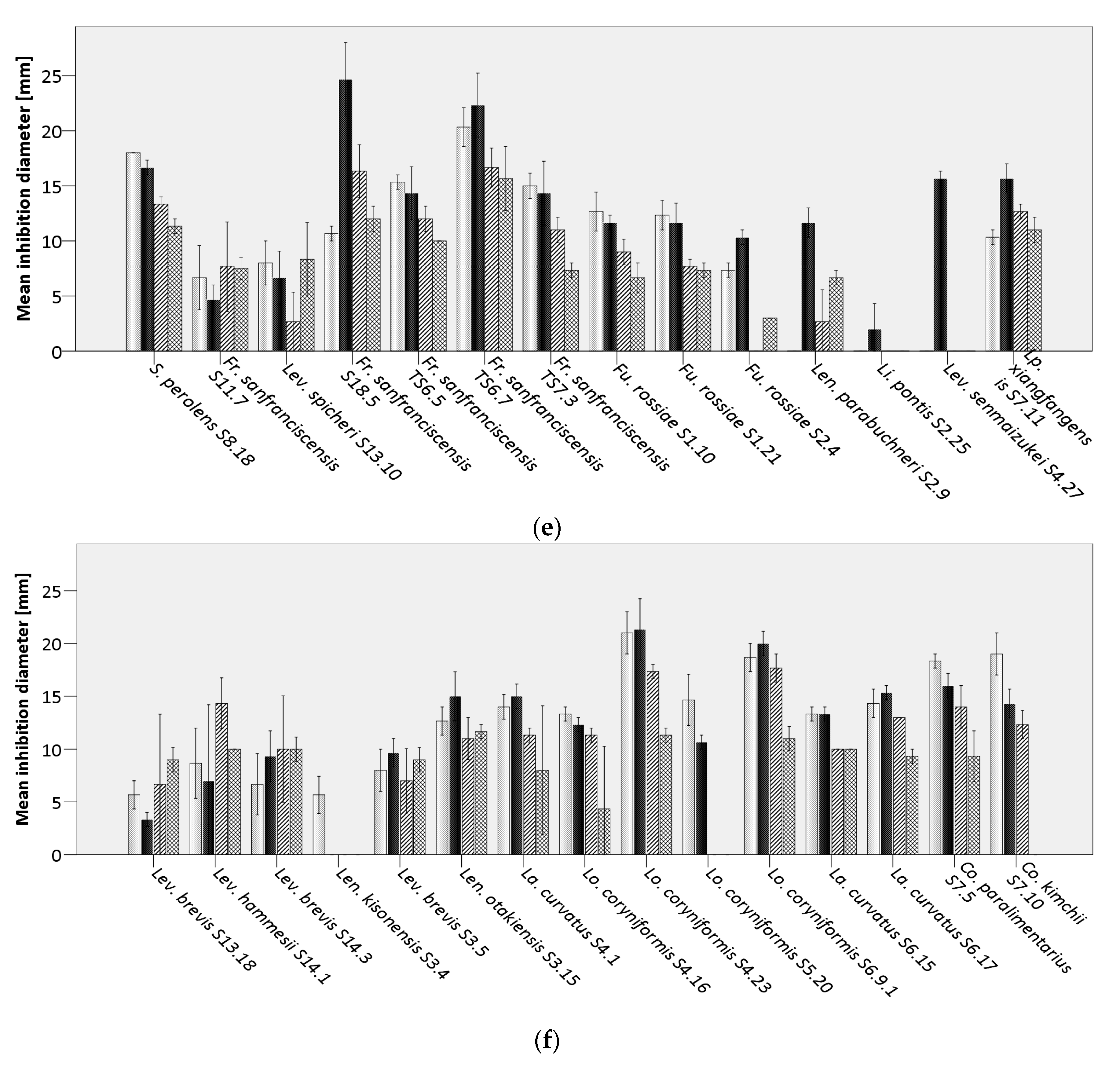
 Bacillus cereus DSM31,
Bacillus cereus DSM31,  B. licheniformis DSM13,
B. licheniformis DSM13,  B. subtilis LMG7135, and
B. subtilis LMG7135, and  B. subtilis S15.20, by (a) Weissella spp., (b) Pediococcus spp., (c) Lacticaseibacillus paracasei, (d) Lactiplantibacillus plantarum, (e,f) further lactobacilli.
Bacillus cereus DSM31, B. licheniformis DSM13, B. subtilis LMG7135, and B. subtilis S15.20, by (a) Weissella spp., (b) Pediococcus spp., (c) Lacticaseibacillus paracasei, (d) Lactiplantibacillus plantarum, (e,f) further lactobacilli.
B. subtilis S15.20, by (a) Weissella spp., (b) Pediococcus spp., (c) Lacticaseibacillus paracasei, (d) Lactiplantibacillus plantarum, (e,f) further lactobacilli.
Bacillus cereus DSM31, B. licheniformis DSM13, B. subtilis LMG7135, and B. subtilis S15.20, by (a) Weissella spp., (b) Pediococcus spp., (c) Lacticaseibacillus paracasei, (d) Lactiplantibacillus plantarum, (e,f) further lactobacilli.



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Strain | Source of Isolation |
|---|---|---|
| Bacillus cereus | DSM 1 31 | unknown |
| Bacillus licheniformis | DSM 1 13 | unknown |
| Bacillus subtilis | DSM 1 7135 | unknown |
| Bacillus subtilis | isolate S15.20 | sourdough |
| Aspergillus flavus | MUCL 2 11945 | wheat flour |
| Aspergillus fumigatus | n.d. 3 | unknown |
| Aspergillus brasiliensis | DSM 1 1988 | Vaccinium sp., fruit |
| Penicillium roqueforti | DSM 1 1079 | gorgonzola cheese |
| Gibberella zeae [anamorph Fusarium graminearum] | MUCL 2 43764 | wheat before malting |
| Antifungal Performance on MRS|APT Medium 1 | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Isolate | A. brasiliensis DSM1988 | A. fumigatus | Pe. roqueforti DSM1079 | F. graminearum MUCL43764 | A. flavus MUCL11945 | Isolate | A. brasiliensis DSM1988 | A. fumigatus | Pe. roqueforti DSM1079 | F. graminearum MUCL43764 | A. flavus MUCL11945 |
| Co. paralimentarius S7.5 | +|- | +++|- | -|- | +++|- | -|- | Len. parabuchneri S2.16 | ++|- | +++|- | +|- | +++|+ | ++|- |
| Fr. sanfranciscensis S18.5 | +++|- | +++|+ | +|- | +++|+++ | ++|- | Len. parabuchneri S2.9 | +++|- | +++|- | +++|- | +++|++ | ++|- |
| Fr. sanfranciscensis TS6.5 | -|- | -|- | -|- | +++|- | -|- | Lev. brevis S13.18 | ++|- | +++|- | -|- | +++|- | ++|- |
| Fr. sanfranciscensis TS6.7 | -|- | -|- | -|- | +++|- | -|- | Lev. brevis S14.3 | ++|- | +++|- | -|- | +++|++ | +|- |
| Fr. sanfranciscensis TS7.3 | -|- | +|- | -|- | +++|+++ | ++|- | Lev. brevis S3.5 | +++|- | +++|- | ++|- | +++|+++ | -|- |
| Fr. sanfranciscensis S11.7 | -|- | -|+ | -|- | +++|+++ | ++|- | Lev. brevis S4.5 | ++|- | +++|- | +++|- | +++|+++ | -|- |
| Fu. rossiae S1.10 | ++|- | +++|- | +|- | +++|++ | ++|- | Lev. brevis S6.13 | +++|- | ++|- | -|- | +++|+ | -|- |
| Fu. rossiae S1.21 | +|- | ++|- | -|- | +++|+++ | +|- | Lev. hammesii S14.1 | ++|- | +++|- | +|++ | +++|++ | ++|- |
| Fu. rossiae S2.4 | ++|- | +++|- | ++|- | +++|+++ | +|- | Lev. kimchii S7.10 | +|- | +|- | -|- | +++|+++ | +|- |
| La. curvatus S4.1 | +|- | +|- | -|- | n.a.2 | n.a.2 | Lev. senmaizukei S4.24 | ++|- | ++|- | -|- | +++|+++ | ++|- |
| La. curvatus S4.14 | -|- | -|- | -|- | +++|- | -|- | Lev. senmaizukei S4.27 | +++|- | +++|- | -|- | +++|+++ | +|- |
| La. curvatus S5.22 | -|- | -|- | -|- | +++|- | -|- | Lev. senmaizukei S5.18 | +|- | +++|- | ++|- | +++|+++ | +|- |
| La. curvatus S6.15 | -|- | -|- | -|- | +++|- | -|- | Lev. senmaizukei S6.21 | ++|- | ++|- | -|- | +++|+++ | +|- |
| L. paracasei S2.21 | +|- | -|- | -|- | +++|- | -|- | Lev. spicheri S4.26 | ++|- | +++|- | -|- | +++|+++ | ++|- |
| L. paracasei S8.13 | +|- | +++|- | +|- | +++|- | ++|- | Lev. spicheri S6.10 | +|- | ++|- | -|- | +++|+ | +|- |
| L. paracasei S8.18 | -|- | -|- | +|- | +++|+ | -|- | Lev. spicheri S13.10 | ++|- | +++|- | ++|- | +++|+++ | +|- |
| L. paracasei S8.21 | ++|- | +++|- | -|- | +++|++ | +|- | Li. pontis S15.14 | -|- | -|- | -|- | -|- | -|- |
| L. paracasei S8.24 | ++|- | ++|- | -|- | +++|- | +|- | Li. pontis S15.3 | -|- | -|- | -|- | -|- | -|- |
| L. paracasei S8.3 | ++|- | +++|- | -|- | +++|- | ++|- | Li. pontis S2.25 | ++|- | +++|- | -|- | +++|+++ | ++|- |
| L. paracasei S8.8 | ++|- | ++|- | -|- | +++|- | +|- | Lo. coryniformis S4.16 | ++|- | +++|- | -|- | +++|+ | ++|- |
| L. paracasei S9.11 | -|- | +|- | +++|- | +++|- | +|- | Lo. coryniformis S4.23 | +|- | ++|- | -|- | +++|++ | -|- |
| L. paracasei S9.15 | +|- | +++|- | -|- | +++|- | +|- | Lo. coryniformis S4.4.2 | ++|- | ++|- | +++|- | n.a.2|+++ | n.a.2 |
| L. paracasei S9.18 | +|- | +++|- | -|- | +++|+ | +|- | Lo. coryniformis S5.20 | +|- | ++|- | -|- | +++|++ | +|- |
| L. paracasei S9.20 | +|- | ++|- | -|- | +++|+ | ++|- | Lo. coryniformis S6.9.1 | -|- | ++|- | -|- | +++|++ | +|- |
| L. paracasei S9.24 | -|- | +++|- | -|- | +++|- | +|- | Lp. plantarum S13.8.2 | +++|- | +++|- | +|- | +++|- | -|- |
| L. paracasei S9.3 | -|- | ++|- | -|- | +++|- | ++|- | Lp. plantarum S4.2 | +++|- | +++|- | +++|- | +++|+++ | ++|- |
| L. paracasei S9.8 | -|- | +++|- | -|- | +++|+ | +|- | Lp. plantarum S5.16 | +|- | +++|- | -|- | +++|++ | ++|- |
| Lat. sakei S4.19 | -|- | -|- | -|- | +++|- | -|- | Lp. plantarum S6.3 | ++|- | +++|- | +|- | +++|+++ | +|- |
| Len. diolivorans S3.2 | +|- | +|- | +|- | +++|++ | +|- | Lp. plantarum S7.4 | -|- | +++|- | -|- | +++|- | -|- |
| Len. kisonensis S3.10 | +|- | ++|- | +|- | +++|++ | +|- | Lp. plantarum S10.12 | ++|- | +++|- | +|+ | +++|++ | ++|- |
| Len. kisonensis S3.4 | +++|- | +++|- | +|- | +++|++ | ++|- | Lp. plantarum S10.13 | +|- | +++|- | ++|- | +++|++ | ++|- |
| Len. otakiensis S3.15 | -|- | -|- | -|- | +++|- | -|- | Lp. plantarum S10.15 | +|- | +++|- | +|+ | +++|+++ | ++|- |
| Lp. plantarum S10.19 | +|- | +++|- | -|- | +++|++ | ++|- | P. pentosaceus S10.10 | ++|- | +++|- | +|- | +++|++ | -|- |
| Lp. plantarum S10.2 | ++|- | +++|- | -|++ | +++|++ | +|- | P. pentosaceus S10.11 | ++|- | +++|- | +|- | +++|+ | +|- |
| Lp. plantarum S10.9 | ++|- | +++|- | +|- | +++|++ | +|- | P. pentosaceus S10.22 | ++|- | ++|- | -|- | +++|++ | ++|- |
| Lp. plantarum S13.13 | +++|- | +++|- | +|- | +++|+++ | ++|- | P. pentosaceus S5.19 | ++|- | ++|- | -|- | +++|+++ | +|- |
| Lp. xiangfangensis S7.11 | ++|- | +++|- | -|- | +++|++ | +|- | W. cibaria S10.4 | ++|- | +++|- | -|++ | +++|++ | +|- |
| Pa. vaccinostercus S6.20 | -|- | -|- | -|- | -|- | -|- | W. viridescens S2.3 | ++|- | +++|- | -|- | +++|+++ | ++|- |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fraberger, V.; Ammer, C.; Domig, K.J. Functional Properties and Sustainability Improvement of Sourdough Bread by Lactic Acid Bacteria. Microorganisms 2020, 8, 1895. https://doi.org/10.3390/microorganisms8121895
Fraberger V, Ammer C, Domig KJ. Functional Properties and Sustainability Improvement of Sourdough Bread by Lactic Acid Bacteria. Microorganisms. 2020; 8(12):1895. https://doi.org/10.3390/microorganisms8121895
Chicago/Turabian StyleFraberger, Vera, Claudia Ammer, and Konrad J. Domig. 2020. "Functional Properties and Sustainability Improvement of Sourdough Bread by Lactic Acid Bacteria" Microorganisms 8, no. 12: 1895. https://doi.org/10.3390/microorganisms8121895
APA StyleFraberger, V., Ammer, C., & Domig, K. J. (2020). Functional Properties and Sustainability Improvement of Sourdough Bread by Lactic Acid Bacteria. Microorganisms, 8(12), 1895. https://doi.org/10.3390/microorganisms8121895