The T3SS of Shigella: Expression, Structure, Function, and Role in Vacuole Escape

 , ,
, , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
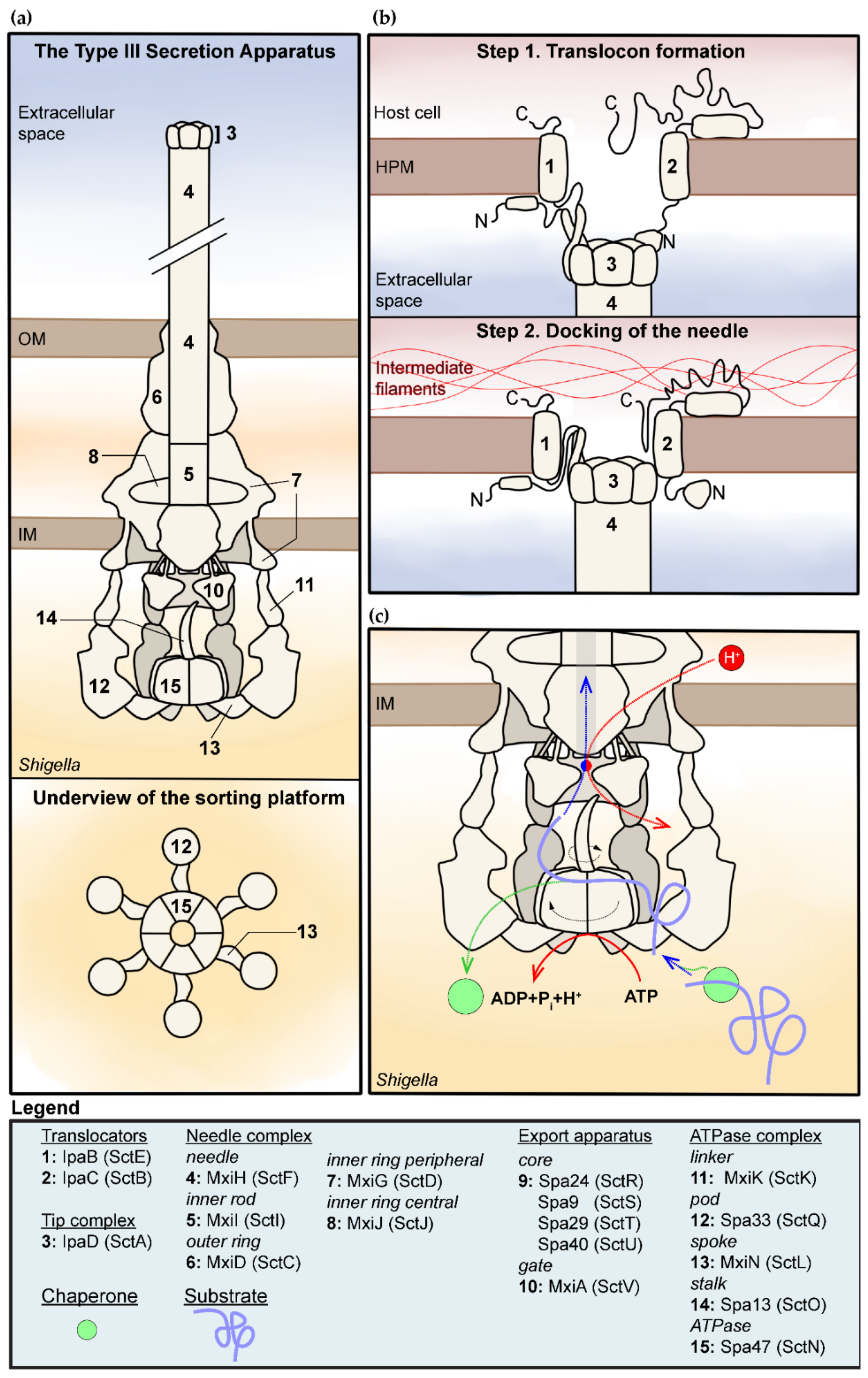
2. The Structure of the T3SA
2.1. Overview of the T3SA
2.2. The Detailed Structure of the T3SA
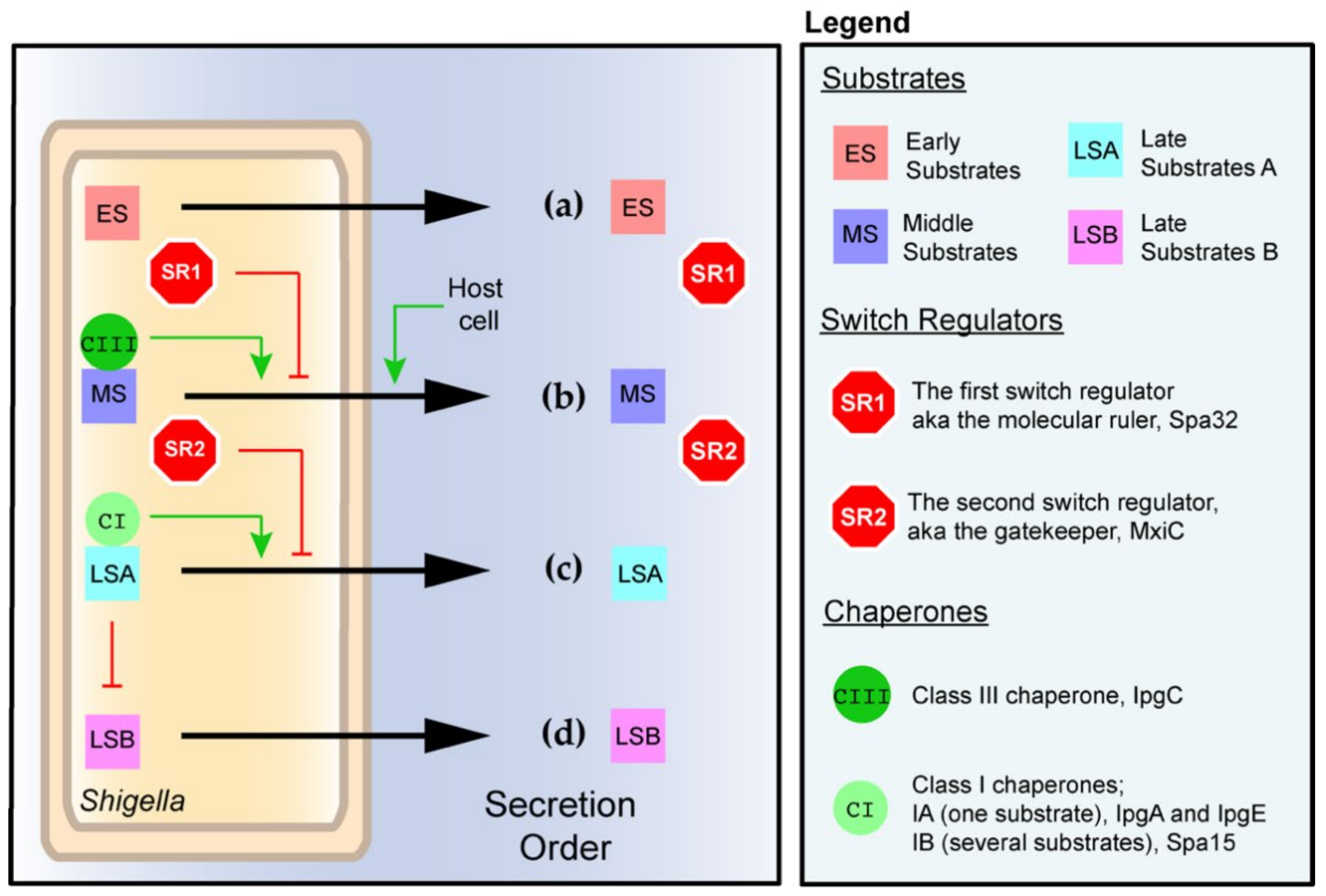
2.3. The Assembly of the T3SA and the Orderly Secretion of Its Substrates
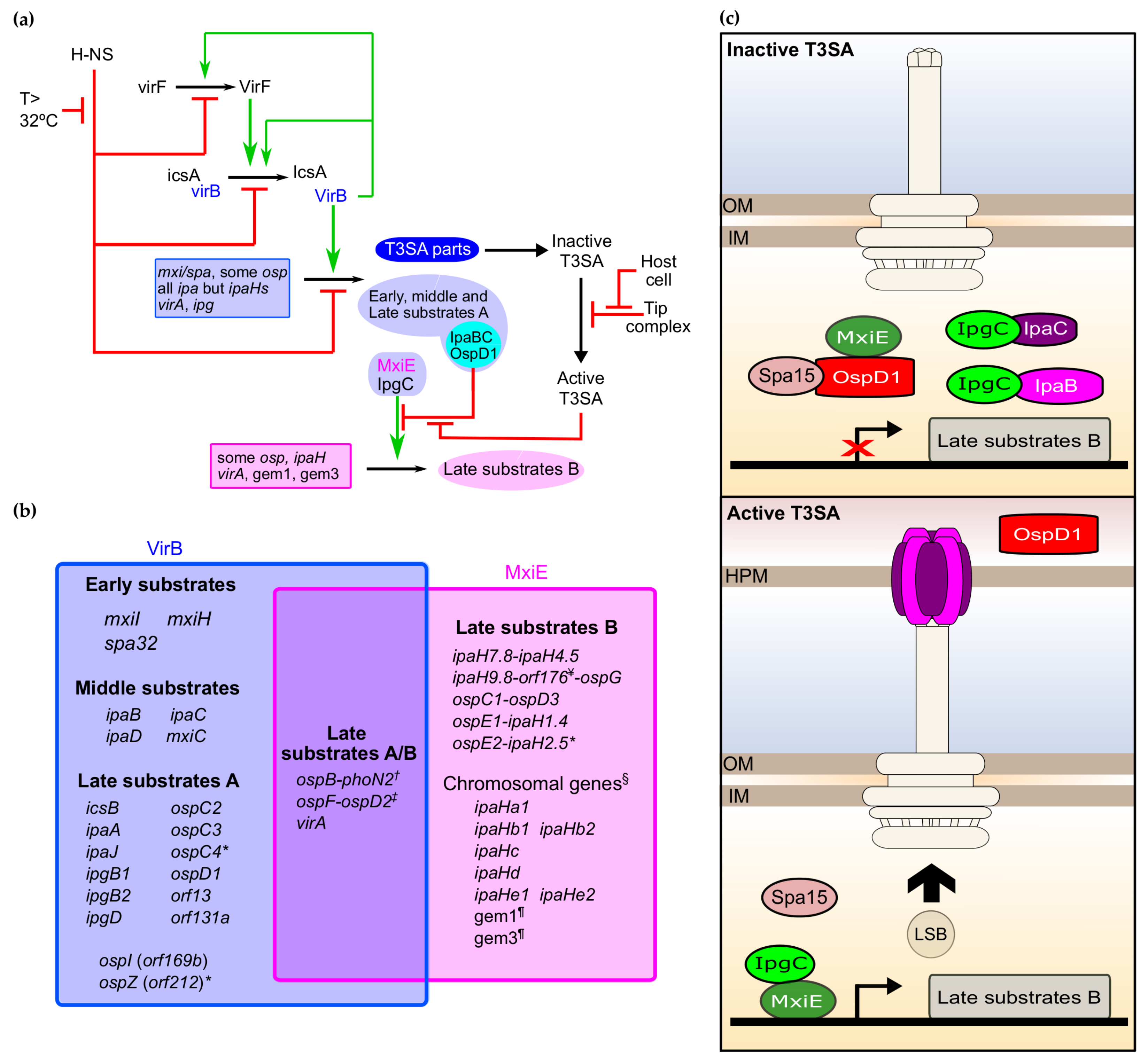
3. The Regulation of the Expression of the T3SS in Shigella
3.1. Overview of the Regulatory Cascade of the T3SS
3.2. Master Regulators of the T3SS in Shigella: H-NS, VirF, and VirB
3.3. The MxiE Regulon
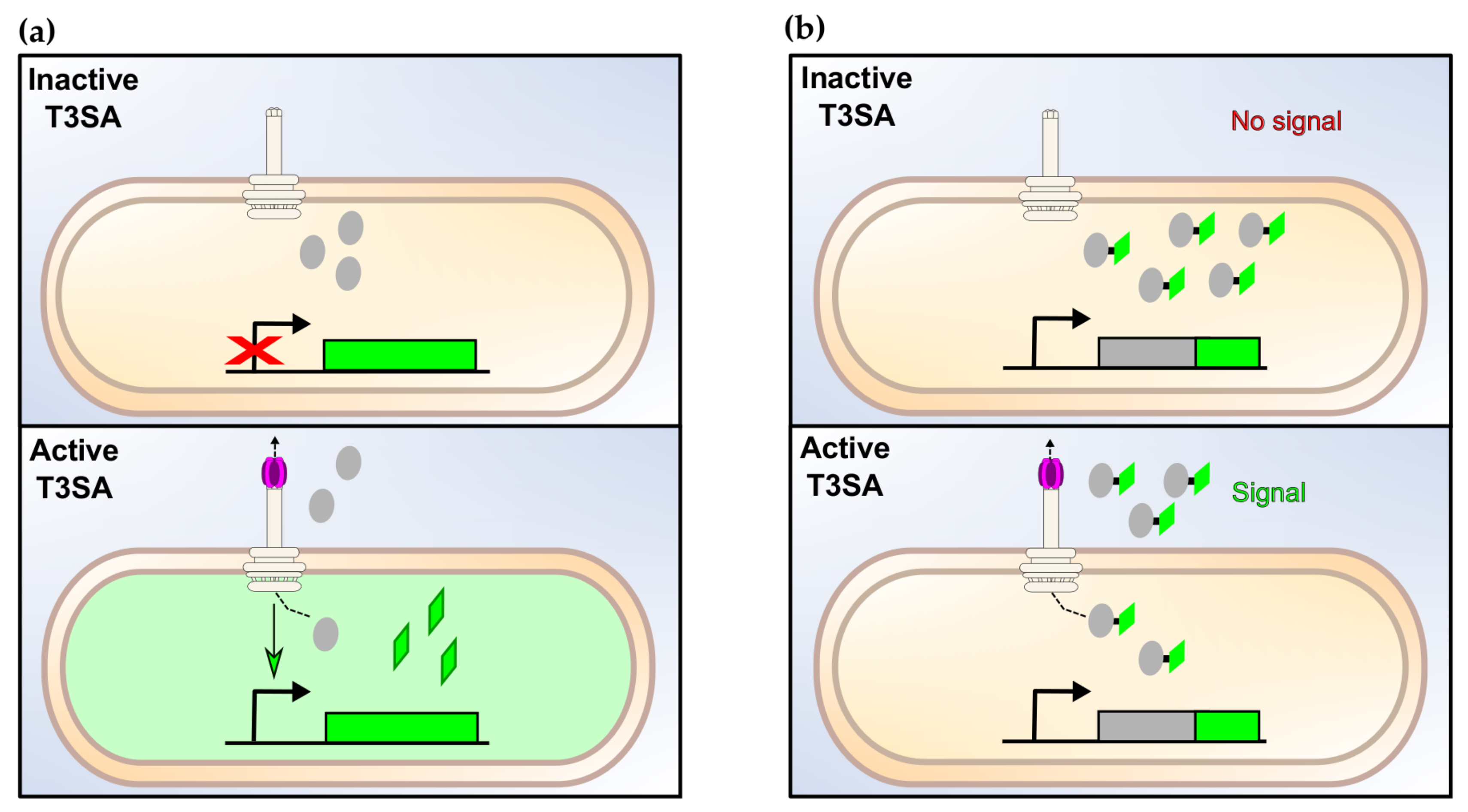
4. Genetically-Encoded Reporters to Monitor the Activity of the T3SA
4.1. Transcription-Based Assays
4.2. Direct Secretion Assays
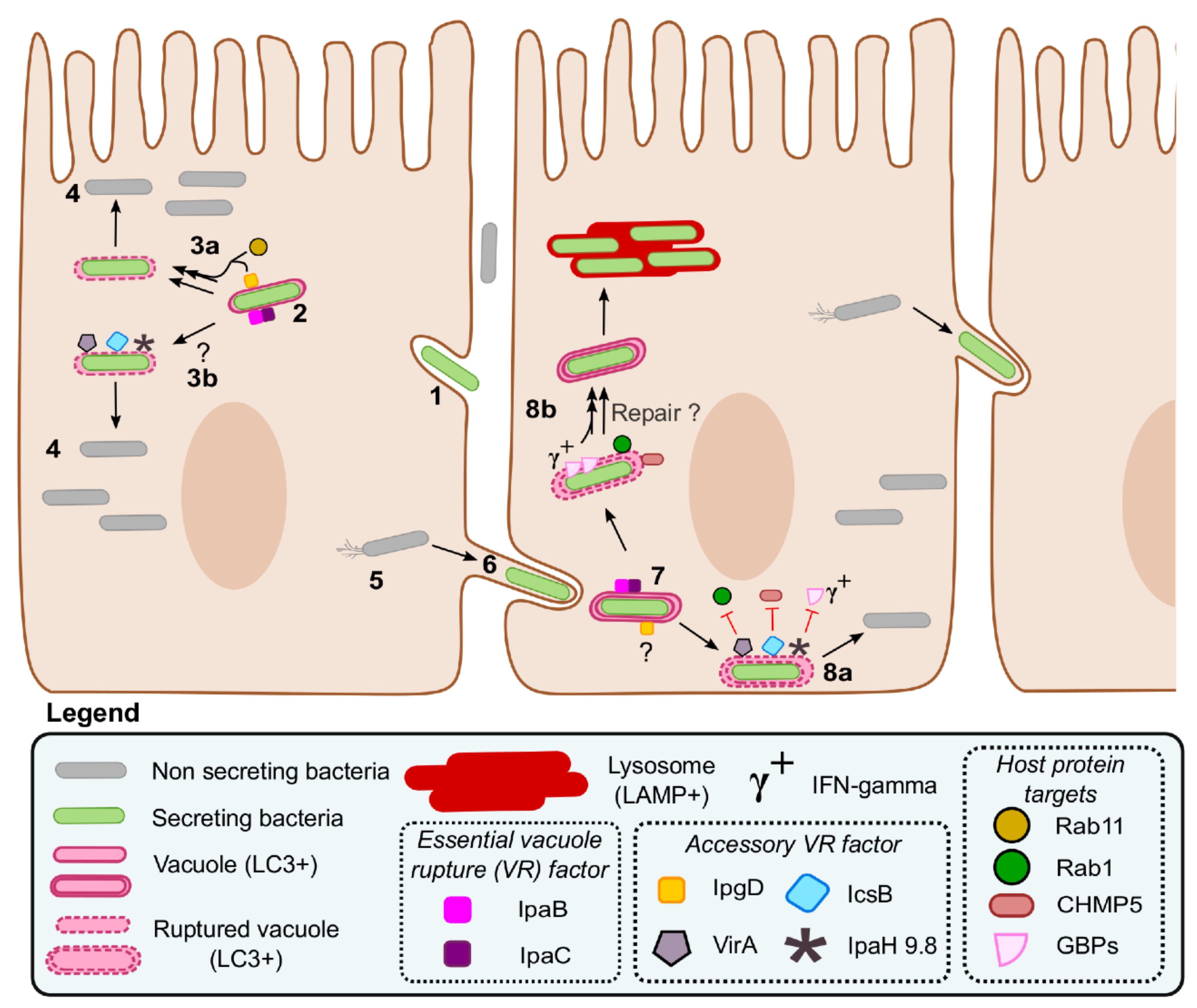
5. Mechanisms of Vacuole Rupture and Escape in Shigella
5.1. The Intracellular Niche of Shigella
5.2. Effectors Implicated in the Escape from the Dissemination Vacuole
6. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kotloff, K.L.; Riddle, M.S.; Platts-Mills, J.A.; Pavlinac, P.; Zaidi, A.K.M. Shigellosis. Lancet 2018, 391, 801–812. [Google Scholar] [CrossRef]
- Lampel, K.A.; Formal, S.B.; Maurelli, A.T. A brief history of Shigella. EcoSal Plus 2018, 8, 1–25. [Google Scholar] [CrossRef] [PubMed]
- Campbell-Valois, F.-X.; Pontier, S.M. Implications of spatiotemporal regulation of Shigella flexneri type three secretion activity on effector functions: Think globally, act locally. Front. Cell. Infect. Microbiol. 2016, 6, 28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Izoré, T.; Job, V.; Dessen, A. Biogenesis, regulation, and targeting of the type III secretion system. Structure 2011, 19, 603–612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deng, W.; Marshall, N.C.; Rowland, J.L.; McCoy, J.M.; Worrall, L.J.; Santos, A.S.; Strynadka, N.C.J.; Finlay, B.B. Assembly, structure, function and regulation of type III secretion systems. Nat. Rev. Microbiol. 2017, 15, 323–337. [Google Scholar] [CrossRef] [PubMed]
- Portaliou, A.G.; Tsolis, K.C.; Loos, M.S.; Zorzini, V.; Economou, A. Type III secretion: Building and operating a remarkable nanomachine. Trends Biochem. Sci. 2016, 41, 175–189. [Google Scholar] [CrossRef]
- Wagner, S.; Grin, I.; Malmsheimer, S.; Singh, N.; Torres-Vargas, C.E.; Westerhausen, S. Bacterial type III secretion systems: A complex device for the delivery of bacterial effector proteins into eukaryotic host cells. FEMS Microbiol. Lett. 2018, 365. [Google Scholar] [CrossRef] [Green Version]
- McVicker, G.; Tang, C.M. Deletion of toxin-antitoxin systems in the evolution of Shigella sonnei as a host-adapted pathogen. Nat. Microbiol. 2016, 2, 16204–16211. [Google Scholar] [CrossRef]
- Cervantes-Rivera, R.; Tronnet, S.; Puhar, A. Complete genome sequence and annotation of the laboratory reference strain Shigella flexneri serotype 5a M90T and genome-wide transcriptional start site determination. BMC Genom. 2020, 21, 285. [Google Scholar] [CrossRef] [Green Version]
- Buchrieser, C.; Glaser, P.; Rusniok, C.; Nedjari, H.; d’Hauteville, H.; Kunst, F.; Sansonetti, P.; Parsot, C. The virulence plasmid pWR100 and the repertoire of proteins secreted by the type III secretion apparatus of Shigella flexneri. Mol. Microbiol. 2000, 38, 760–771. [Google Scholar] [CrossRef]
- Venkatesan, M.M.; Goldberg, M.B.; Rose, D.J.; Grotbeck, E.J.; Burland, V.; Blattner, F.R. Complete DNA sequence and analysis of the large virulence plasmid of Shigella flexneri. Infect. Immun. 2001, 69, 3271–3285. [Google Scholar] [CrossRef] [Green Version]
- Edgren, T.; Forsberg, A.; Rosqvist, R.; Wolf-Watz, H. Type III secretion in Yersinia: Injectisome or not? PLoS Pathog. 2012, 8, e1002669. [Google Scholar] [CrossRef] [Green Version]
- Kubori, T.; Matsushima, Y.; Nakamura, D.; Uralil, J.; Lara-Tejero, M.; Sukhan, A.; Galan, J.E.; Aizawa, S.I. Supramolecular structure of the Salmonella typhimurium type III protein secretion system. Science 1998, 280, 602–605. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blocker, A.; Jouihri, N.; Larquet, E.; Gounon, P.; Ebel, F.; Parsot, C.; Sansonetti, P.; Allaoui, A. Structure and composition of the Shigella flexneri “needle complex,” a part of its type III secreton. Mol. Microbiol. 2001, 39, 652–663. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.; Worrall, L.J.; Strynadka, N.C. Towards capture of dynamic assembly and action of the T3SS at near atomic resolution. Curr. Opin. Struct. Biol. 2020, 61, 71–78. [Google Scholar] [CrossRef]
- Hu, B.; Morado, D.R.; Margolin, W.; Rohde, J.R.; Arizmendi, O.; Picking, W.L.; Picking, W.D.; Liu, J. Visualization of the type III secretion sorting platform of Shigella flexneri. Proc. Natl. Acad. Sci. USA 2015, 112, 1047–1052. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, B.; Lara-Tejero, M.; Kong, Q.; Galán, J.E.; Liu, J. In situ molecular architecture of the salmonella type III secretion machine. Cell 2017, 168, 1065–1074. [Google Scholar] [CrossRef] [Green Version]
- Diepold, A.; Armitage, J.P. Type III secretion systems: The bacterial flagellum and the injectisome. Philos. Trans. R. Soc. Lond. Bbiol. Sci. 2015, 370, 20150020. [Google Scholar] [CrossRef] [Green Version]
- Habenstein, B.; El Mammeri, N.; Tolchard, J.; Lamon, G.; Tawani, A.; Berbon, M.; Loquet, A. Structures of type III secretion system needle filaments. Curr. Top. Microbiol. Immunol. 2020, 427, 109–131. [Google Scholar] [PubMed]
- Demers, J.-P.; Sgourakis, N.G.; Gupta, R.; Loquet, A.; Giller, K.; Riedel, D.; Laube, B.; Kolbe, M.; Baker, D.; Becker, S.; et al. The common structural architecture of Shigella flexneri and Salmonella typhimurium type three secretion needles. PLoS Pathog. 2013, 9, e1003245. [Google Scholar] [CrossRef] [Green Version]
- Demers, J.-P.; Habenstein, B.; Loquet, A.; Kumar Vasa, S.; Giller, K.; Becker, S.; Baker, D.; Lange, A.; Sgourakis, N.G. High-resolution structure of the Shigella type-III secretion needle by solid-state NMR and cryo-electron microscopy. Nat. Commun. 2014, 5, 4976–5012. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verasdonck, J.; Shen, D.-K.; Treadgold, A.; Arthur, C.; Böckmann, A.; Meier, B.H.; Blocker, A.J. Reassessment of MxiH subunit orientation and fold within native Shigella T3SS needles using surface labelling and solid-state NMR. J. Struct. Biol. 2015, 192, 441–448. [Google Scholar] [CrossRef] [PubMed]
- Epler, C.R.; Dickenson, N.E.; Bullitt, E.; Picking, W.L. Ultrastructural analysis of IpaD at the tip of the nascent MxiH type III secretion apparatus of Shigella flexneri. J. Mol. Biol. 2012, 420, 29–39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheung, M.; Shen, D.-K.; Makino, F.; Kato, T.; Roehrich, A.D.; Martinez-Argudo, I.; Walker, M.L.; Murillo, I.; Liu, X.; Pain, M.; et al. Three-dimensional electron microscopy reconstruction and cysteine-mediated crosslinking provide a model of the type III secretion system needle tip complex. Mol. Microbiol. 2015, 95, 31–50. [Google Scholar] [CrossRef]
- Olive, A.J.; Kenjale, R.; Espina, M.; Moore, D.S.; Picking, W.L.; Picking, W.D. Bile salts stimulate recruitment of IpaB to the Shigella flexneri surface, where it colocalizes with IpaD at the tip of the type III secretion needle. Infect. Immun. 2007, 75, 2626–2629. [Google Scholar] [CrossRef] [Green Version]
- Stensrud, K.F.; Adam, P.R.; La Mar, C.D.; Olive, A.J.; Lushington, G.H.; Sudharsan, R.; Shelton, N.L.; Givens, R.S.; Picking, W.L.; Picking, W.D. Deoxycholate interacts with IpaD of Shigella flexneri in inducing the recruitment of IpaB to the type III secretion apparatus needle tip. J. Biol. Chem. 2008, 283, 18646–18654. [Google Scholar] [CrossRef] [Green Version]
- Veenendaal, A.K.J.; Hodgkinson, J.L.; Schwarzer, L.; Stabat, D.; Zenk, S.F.; Blocker, A.J. The type III secretion system needle tip complex mediates host cell sensing and translocon insertion. Mol. Microbiol. 2007, 63, 1719–1730. [Google Scholar] [CrossRef]
- Menard, R.; Sansonetti, P.; Parsot, C. The secretion of the Shigella flexneri Ipa invasins is activated by epithelial cells and controlled by IpaB and IpaD. EMBO J. 1994, 13, 5293–5302. [Google Scholar] [CrossRef]
- Dickenson, N.E.; Arizmendi, O.; Patil, M.K.; Toth, R.T.; Middaugh, C.R.; Picking, W.D.; Picking, W.L. N-terminus of IpaB provides a potential anchor to the Shigella type III secretion system tip complex protein IpaD. Biochemistry 2013, 52, 8790–8799. [Google Scholar] [CrossRef] [Green Version]
- Barta, M.L.; Tachiyama, S.; Muthuramalingam, M.; Arizmendi, O.; Villanueva, C.E.; Ramyar, K.X.; Geisbrecht, B.V.; Lovell, S.; Battaile, K.P.; Picking, W.L.; et al. Using disruptive insertional mutagenesis to identify the in situ structure-function landscape of the Shigella translocator protein IpaB. Protein Sci. 2018, 27, 1392–1406. [Google Scholar] [CrossRef] [Green Version]
- Tachiyama, S.; Chang, Y.; Muthuramalingam, M.; Hu, B.; Barta, M.L.; Picking, W.L.; Liu, J.; Picking, W.D. The cytoplasmic domain of MxiG interacts with MxiK and directs assembly of the sorting platform in the Shigella type III secretion system. J. Biol. Chem. 2019, 294, 19184–19196. [Google Scholar] [CrossRef] [PubMed]
- Butan, C.; Lara-Tejero, M.; Li, W.; Liu, J.; Galán, J.E. High-resolution view of the type III secretion export apparatus in situ reveals membrane remodeling and a secretion pathway. Proc. Natl. Acad. Sci. USA 2019, 116, 24786–24795. [Google Scholar] [CrossRef] [PubMed]
- Russo, B.C.; Stamm, L.M.; Raaben, M.; Kim, C.M.; Kahoud, E.; Robinson, L.R.; Bose, S.; Queiroz, A.L.; Herrera, B.B.; Baxt, L.A.; et al. Intermediate filaments enable pathogen docking to trigger type 3 effector translocation. Nat. Microbiol. 2016, 1, 16025. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Russo, B.C.; Duncan, J.K.; Wiscovitch, A.L.; Hachey, A.C.; Goldberg, M.B. Activation of Shigella flexneri type 3 secretion requires a host-induced conformational change to the translocon pore. PLoS Pathog. 2019, 15, e1007928. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Allison, S.E.; Tuinema, B.R.; Everson, E.S.; Sugiman-Marangos, S.; Zhang, K.; Junop, M.S.; Coombes, B.K. Identification of the docking site between a type III secretion system ATPase and a chaperone for effector cargo. J. Biol. Chem. 2014, 289, 23734–23744. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Majewski, D.D.; Worrall, L.J.; Hong, C.; Atkinson, C.E.; Vuckovic, M.; Watanabe, N.; Yu, Z.; Strynadka, N.C.J. Cryo-EM structure of the homohexameric T3SS ATPase-central stalk complex reveals rotary ATPase-like asymmetry. Nat. Commun. 2019, 10, 626. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Minamino, T.; Morimoto, Y.V.; Hara, N.; Aldridge, P.D.; Namba, K. The bacterial flagellar type III export gate complex is a dual fuel engine that can use both H+ and Na+ for flagellar protein export. PLoS Pathog. 2016, 12, e1005495. [Google Scholar] [CrossRef] [PubMed]
- Terashima, H.; Imada, K. Novel insight into an energy transduction mechanism of the bacterial flagellar type III protein export. Biophys. Physicobiol. 2018, 15, 173–178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lunelli, M.; Kamprad, A.; Bürger, J.; Mielke, T.; Spahn, C.M.T.; Kolbe, M. Cryo-EM structure of the Shigella type III needle complex. PLoS Pathog. 2020, 16, e1008263. [Google Scholar] [CrossRef] [PubMed]
- Torres-Vargas, C.E.; Kronenberger, T.; Roos, N.; Dietsche, T.; Poso, A.; Wagner, S. The inner rod of virulence-associated type III secretion systems constitutes a needle adapter of one helical turn that is deeply integrated into the system’s export apparatus. Mol. Microbiol. 2019, 112, 918–931. [Google Scholar] [CrossRef] [PubMed]
- Johnson, S.; Kuhlen, L.; Deme, J.C.; Abrusci, P.; Lea, S.M. The structure of an injectisome export gate demonstrates conservation of architecture in the core export gate between flagellar and virulence type III secretion systems. mBio 2019, 10, 111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, J.; Worrall, L.J.; Hong, C.; Vuckovic, M.; Atkinson, C.E.; Caveney, N.; Yu, Z.; Strynadka, N.C.J. Cryo-EM analysis of the T3S injectisome reveals the structure of the needle and open secretin. Nat. Commun. 2018, 9, 3840–3911. [Google Scholar] [CrossRef] [PubMed]
- Kuhlen, L.; Johnson, S.; Zeitler, A.; Bäurle, S.; Deme, J.C.; Caesar, J.J.E.; Debo, R.; Fisher, J.; Wagner, S.; Lea, S.M. The substrate specificity switch FlhB assembles onto the export gate to regulate type three secretion. Nat. Commun. 2020, 11, 1296–1310. [Google Scholar] [CrossRef] [PubMed]
- Abrusci, P.; Vergara-Irigaray, M.; Johnson, S.; Beeby, M.D.; Hendrixson, D.R.; Roversi, P.; Friede, M.E.; Deane, J.E.; Jensen, G.J.; Tang, C.M.; et al. Architecture of the major component of the type III secretion system export apparatus. Nat. Struct. Mol. Biol. 2013, 20, 99–104. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.; Beeby, M.; Murphy, G.E.; Leadbetter, J.R.; Hendrixson, D.R.; Briegel, A.; Li, Z.; Shi, J.; Tocheva, E.I.; Müller, A.; et al. Structural diversity of bacterial flagellar motors. EMBO J. 2011, 30, 2972–2981. [Google Scholar] [CrossRef] [Green Version]
- Qin, Z.; Tu, J.; Lin, T.; Norris, S.J.; Li, C.; Motaleb, M.A.; Liu, J. Cryo-electron tomography of periplasmic flagella in Borrelia burgdorferi reveals a distinct cytoplasmic ATPase complex. PLoS Biol. 2018, 16, e3000050. [Google Scholar] [CrossRef] [Green Version]
- Gao, X.; Mu, Z.; Yu, X.; Qin, B.; Wojdyla, J.; Wang, M.; Cui, S. Structural Insight into conformational changes induced by ATP binding in a type III secretion-associated ATPase from Shigella flexneri. Front. Microbiol. 2018, 9, 1468. [Google Scholar] [CrossRef]
- McDowell, M.A.; Marcoux, J.; McVicker, G.; Johnson, S.; Fong, Y.H.; Stevens, R.; Bowman, L.A.; Degiacomi, M.T.; Yan, J.; Wise, A.; et al. Characterisation of Shigella Spa33 and Thermotoga FliM/N reveals a new model for C-ring assembly in T3SS. Mol. Microbiol. 2016, 99, 749–766. [Google Scholar] [CrossRef] [Green Version]
- Kadari, M.; Lakhloufi, D.; Delforge, V.; Imbault, V.; Communi, D.; Smeesters, P.; Botteaux, A. Multiple proteins arising from a single gene: The role of the Spa33 variants in Shigella T3SS regulation. MicrobiologyOpen 2019, 8, e932. [Google Scholar] [CrossRef] [Green Version]
- Kimbrough, T.G.; Miller, S.I. Contribution of Salmonella typhimurium type III secretion components to needle complex formation. Proc. Natl. Acad. Sci. USA 2000, 97, 11008–11013. [Google Scholar] [CrossRef] [Green Version]
- Wagner, S.; Königsmaier, L.; Lara-Tejero, M.; Lefebre, M.; Marlovits, T.C.; Galán, J.E. Organization and coordinated assembly of the type III secretion export apparatus. Proc. Natl. Acad. Sci. USA 2010, 107, 17745–17750. [Google Scholar] [CrossRef] [Green Version]
- Dietsche, T.; Tesfazgi Mebrhatu, M.; Brunner, M.J.; Abrusci, P.; Yan, J.; Franz-Wachtel, M.; Schärfe, C.; Zilkenat, S.; Grin, I.; Galán, J.E.; et al. Structural and functional characterization of the bacterial type III secretion export apparatus. PLoS Pathog. 2016, 12, e1006071. [Google Scholar] [CrossRef] [PubMed]
- De Oliviera Silva, Y.R.; Contreras-Martel, C.; Macheboeuf, P.; Dessen, A. Bacterial secretins: Mechanisms of assembly and membrane targeting. Protein Sci. 2020, 29, 893–904. [Google Scholar] [CrossRef] [PubMed]
- Schuch, R.; Maurelli, A.T. MxiM and MxiJ, base elements of the Mxi-Spa type III secretion system of Shigella, interact with and stabilize the MxiD secretin in the cell envelope. J. Bacteriol. 2001, 183, 6991–6998. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sidik, S.; Kottwitz, H.; Benjamin, J.; Ryu, J.; Jarrar, A.; Garduno, R.; Rohde, J.R. A Shigella flexneri virulence plasmid encoded factor controls production of outer membrane vesicles. G3 (Bethesda) 2014, 4, 2493–2503. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diepold, A.; Amstutz, M.; Abel, S.; Sorg, I.; Jenal, U.; Cornelis, G.R. Deciphering the assembly of the Yersinia type III secretion injectisome. EMBO J. 2010, 29, 1928–1940. [Google Scholar] [CrossRef] [Green Version]
- Parsot, C.; Menard, R.; Gounon, P.; Sansonetti, P.J. Enhanced secretion through the Shigella flexneri Mxi-Spa translocon leads to assembly of extracellular proteins into macromolecular structures. Mol. Microbiol. 1995, 16, 291–300. [Google Scholar] [CrossRef]
- Pinaud, L.; Ferrari, M.L.; Friedman, R.; Jehmlich, N.; von Bergen, M.; Phalipon, A.; Sansonetti, P.J.; Campbell-Valois, F.-X. Identification of novel substrates of Shigella T3SA through analysis of its virulence plasmid-encoded secretome. PLoS ONE 2017, 12, e0186920. [Google Scholar] [CrossRef] [Green Version]
- Journet, L.; Agrain, C.; Broz, P.; Cornelis, G.R. The needle length of bacterial injectisomes is determined by a molecular ruler. Science 2003, 302, 1757–1760. [Google Scholar] [CrossRef]
- Magdalena, J.; Hachani, A.; Chamekh, M.; Jouihri, N.; Gounon, P.; Blocker, A.; Allaoui, A.I.M. Spa32 regulates a switch in substrate specificity of the type III secreton of Shigella flexneri from needle components to Ipa proteins. J. Bacteriol. 2002, 184, 3433–3441. [Google Scholar] [CrossRef] [Green Version]
- Botteaux, A.; Sani, M.; Kayath, C.A.; Boekema, E.J.; Allaoui, A. Spa32 interaction with the inner-membrane Spa40 component of the type III secretion system of Shigella flexneri is required for the control of the needle length by a molecular tape measure mechanism. Mol. Microbiol. 2008, 70, 1515–1528. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, D.-K.; Moriya, N.; Martinez-Argudo, I.; Blocker, A.J. Needle length control and the secretion substrate specificity switch are only loosely coupled in the type III secretion apparatus of Shigella. Microbiology 2012, 158, 1884–1896. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Botteaux, A.; Kayath, C.A.; Page, A.L.; Jouihri, N.; Sani, M.; Boekema, E.; Biskri, L.; Parsot, C.; Allaoui, A. The 33 carboxyl-terminal residues of Spa40 orchestrate the multi-step assembly process of the type III secretion needle complex in Shigella flexneri. Microbiology 2010, 156, 2807–2817. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ho, O.; Rogne, P.; Edgren, T.; Wolf-Watz, H.; Login, F.H.; Wolf-Watz, M. Characterization of the ruler protein interaction interface on the substrate specificity switch protein in the Yersinia type III secretion system. J. Biol. Chem. 2017, 292, 3299–3311. [Google Scholar] [CrossRef] [Green Version]
- Deane, J.E.; Graham, S.C.; Mitchell, E.P.; Flot, D.; Johnson, S.; Lea, S.M. Crystal structure of Spa40, the specificity switch for the Shigella flexneri type III secretion system. Mol. Microbiol. 2008, 69, 267–276. [Google Scholar] [CrossRef] [Green Version]
- Murillo, I.; Martinez-Argudo, I.; Blocker, A.J. Genetic dissection of the signaling cascade that controls activation of the Shigella type III secretion system from the needle tip. Sci. Rep. 2016, 6, 27649. [Google Scholar] [CrossRef] [Green Version]
- Picking, W.L.; Picking, W.D. The many faces of IpaB. Front. Cell. Infect. Microbiol. 2016, 6, 12. [Google Scholar] [CrossRef]
- Park, D.; Lara-Tejero, M.; Waxham, M.N.; Li, W.; Hu, B.; Galán, J.E.; Liu, J. Visualization of the type III secretion mediated Salmonella-host cell interface using cryo-electron tomography. Elife 2018, 7, 935. [Google Scholar] [CrossRef]
- Blocker, A.; Gounon, P.; Larquet, E.; Niebuhr, K.; Cabiaux, V.; Parsot, C.; Sansonetti, P. The tripartite type III secreton of Shigella flexneri inserts IpaB and IpaC into host membranes. J. Cell Biol. 1999, 147, 683–693. [Google Scholar] [CrossRef] [Green Version]
- Mounier, J.; Popoff, M.R.; Enninga, J.; Frame, M.C.; Sansonetti, P.J.; van Nhieu, G.T. The IpaC carboxyterminal effector domain mediates Src-dependent actin polymerization during Shigella invasion of epithelial cells. PLoS Pathog. 2009, 5, e1000271. [Google Scholar] [CrossRef] [Green Version]
- Russo, B.C.; Duncan, J.K.; Goldberg, M.B. Topological analysis of the type 3 secretion system Translocon pore protein IpaC following its native delivery to the plasma membrane during infection. mBio 2019, 10, e00877-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roehrich, A.D.; Guillossou, E.; Blocker, A.J.; Martinez-Argudo, I. Shigella IpaD has a dual role: Signal transduction from the type III secretion system needle tip and intracellular secretion regulation. Mol. Microbiol. 2013, 87, 690–706. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kenjale, R.; Wilson, J.; Zenk, S.F.; Saurya, S.; Picking, W.L.; Picking, W.D.; Blocker, A. The needle component of the type III secreton of Shigella regulates the activity of the secretion apparatus. J. Biol. Chem. 2005, 280, 42929–42937. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Demler, H.J.; Case, H.B.; Morales, Y.; Bernard, A.R.; Johnson, S.J.; Dickenson, N.E. Interfacial amino acids support Spa47 oligomerization and shigella type three secretion system activation. Proteins 2019, 87, 931–942. [Google Scholar] [CrossRef] [PubMed]
- Burgess, J.L.; Case, H.B.; Burgess, R.A.; Dickenson, N.E. Dominant negative effects by inactive Spa47 mutants inhibit T3SS function and Shigella virulence. PLoS ONE 2020, 15, e0228227. [Google Scholar] [CrossRef]
- Burgess, J.L.; Burgess, R.A.; Morales, Y.; Bouvang, J.M.; Johnson, S.J.; Dickenson, N.E. Structural and biochemical characterization of Spa47 provides mechanistic insight into type III secretion system ATPase activation and Shigella virulence regulation. J. Biol. Chem. 2016, 291, 25837–25852. [Google Scholar] [CrossRef] [Green Version]
- Case, H.B.; Dickenson, N.E. MxiN differentially regulates monomeric and oligomeric species of the Shigella type three secretion system ATPase Spa47. Biochemistry 2018, 57, 2266–2277. [Google Scholar] [CrossRef]
- Akeda, Y.; Galán, J.E. Chaperone release and unfolding of substrates in type III secretion. Nature 2005, 437, 911–915. [Google Scholar] [CrossRef]
- Cherradi, Y.; Hachani, A.; Allaoui, A. Spa13 of Shigella flexneri has a dual role: Chaperone escort and export gate-activator switch of the type III secretion system. Microbiology 2014, 160, 130–141. [Google Scholar] [CrossRef] [Green Version]
- Ohgita, T.; Hayashi, N.; Hama, S.; Tsuchiya, H.; Gotoh, N.; Kogure, K. A novel effector secretion mechanism based on proton-motive force-dependent type III secretion apparatus rotation. FASEB J. 2013, 27, 2862–2872. [Google Scholar] [CrossRef]
- Ohgita, T.; Saito, H. Biophysical mechanism of protein export by bacterial type III secretion system. Chem. Pharm. Bull. 2019, 67, 341–344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xing, Q.; Shi, K.; Portaliou, A.; Rossi, P.; Economou, A.; Kalodimos, C.G. Structures of chaperone-substrate complexes docked onto the export gate in a type III secretion system. Nat. Commun. 2018, 9, 1773–1779. [Google Scholar] [CrossRef] [PubMed]
- Lee, P.-C.; Rietsch, A. Fueling type III secretion. Trends Microbiol. 2015, 23, 296–300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Botteaux, A.; Sory, M.P.; Biskri, L.; Parsot, C.; Allaoui, A. MxiC is secreted by and controls the substrate specificity of the Shigella flexneri type III secretion apparatus. Mol. Microbiol. 2009, 71, 449–460. [Google Scholar] [CrossRef] [PubMed]
- Roehrich, A.D.; Bordignon, E.; Mode, S.; Shen, D.-K.; Liu, X.; Pain, M.; Murillo, I.; Martinez-Argudo, I.; Sessions, R.B.; Blocker, A.J. Steps for Shigella gatekeeper protein MxiC function in hierarchical type III secretion regulation. J. Biol. Chem. 2017, 292, 1705–1723. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cherradi, Y.; Schiavolin, L.; Moussa, S.; Meghraoui, A.; Meksem, A.; Biskri, L.; Azarkan, M.; Allaoui, A.; Botteaux, A. Interplay between predicted inner-rod and gatekeeper in controlling substrate specificity of the type III secretion system. Mol. Microbiol. 2013, 87, 1183–1199. [Google Scholar] [CrossRef] [Green Version]
- El Hajjami, N.; Moussa, S.; Houssa, J.; Monteyne, D.; Perez-Morga, D.; Botteaux, A. The inner-rod component of Shigella flexneri type 3 secretion system, MxiI, is involved in the transmission of the secretion activation signal by its interaction with MxiC. MicrobiologyOpen 2018, 7, e00520. [Google Scholar] [CrossRef]
- Shen, D.-K.; Blocker, A.J. MxiA, MxiC and IpaD regulate substrate selection and secretion mode in the T3SS of Shigella flexneri. PLoS ONE 2016, 11, e0155141. [Google Scholar] [CrossRef] [Green Version]
- Radics, J.; Königsmaier, L.; Marlovits, T.C. Structure of a pathogenic type 3 secretion system in action. Nat. Struct. Mol. Biol. 2014, 21, 82–87. [Google Scholar] [CrossRef]
- Campbell-Valois, F.-X.; Schnupf, P.; Nigro, G.; Sachse, M.; Sansonetti, P.J.; Parsot, C. A fluorescent reporter reveals on/off regulation of the shigella type III secretion apparatus during entry and cell-to-cell spread. Cell Host Microbe 2014, 15, 177–189. [Google Scholar] [CrossRef] [Green Version]
- Martinez-Argudo, I.; Blocker, A.J. The Shigella T3SS needle transmits a signal for MxiC release, which controls secretion of effectors. Mol. Microbiol. 2010, 78, 1365–1378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lara-Tejero, M.; Kato, J.; Wagner, S.; Liu, X.; Galán, J.E. A sorting platform determines the order of protein secretion in bacterial type III systems. Science 2011, 331, 1188–1191. [Google Scholar] [CrossRef] [Green Version]
- Ernst, N.H.; Reeves, A.Z.; Ramseyer, J.E.; Lesser, C.F. High-Throughput screening of type III secretion determinants reveals a major chaperone-independent pathway. mBio 2018, 9, 415–513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dorman, M.J.; Dorman, C.J. Regulatory hierarchies controlling virulence gene expression in Shigella flexneriand Vibrio cholerae. Front. Microbiol. 2018, 9, 2686. [Google Scholar] [CrossRef] [PubMed]
- Parsot, C. Shigella type III secretion effectors: How, where, when, for what purposes? Curr. Opin. Microbiol. 2009, 12, 110–116. [Google Scholar] [CrossRef]
- Silué, N.; Marcantonio, E.; Campbell-Valois, F.-X. RNA-Seq analysis of the T3SA regulon in Shigella flexneri reveals two new chromosomal genes upregulated in the on-state. Methods 2020, 176, 71–81. [Google Scholar] [CrossRef] [PubMed]
- Bongrand, C.; Sansonetti, P.J.; Parsot, C. Characterization of the promoter, MxiE box and 5’ UTR of genes controlled by the activity of the type III secretion apparatus in Shigella flexneri. PLoS ONE 2012, 7, e32862. [Google Scholar] [CrossRef] [Green Version]
- Deighan, P.; Beloin, C.; Dorman, C.J. Three-way interactions among the Sfh, StpA and H-NS nucleoid-structuring proteins of Shigella flexneri 2a strain 2457T. Mol. Microbiol. 2003, 48, 1401–1416. [Google Scholar] [CrossRef]
- Picker, M.A.; Wing, H.J. H-NS, its family members and their regulation of virulence genes in Shigella species. Genes 2016, 7, 112. [Google Scholar] [CrossRef]
- Beloin, C.; Deighan, P.; Doyle, M.; Dorman, C.J. Shigella flexneri 2a strain 2457T expresses three members of the H-NS-like protein family: Characterization of the Sfh protein. Mol. Genet. Genom. 2003, 270, 66–77. [Google Scholar] [CrossRef]
- Riccardi, E.; Van Mastbergen, E.C.; Navarre, W.W.; Vreede, J. Predicting the mechanism and rate of H-NS binding to AT-rich DNA. PLoS Comput. Biol. 2019, 15, e1006845. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Chen, H.; Kenney, L.J.; Yan, J. A divalent switch drives H-NS/DNA-binding conformations between stiffening and bridging modes. Genes Dev. 2010, 24, 339–344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ali, S.S.; Xia, B.; Liu, J.; Navarre, W.W. Silencing of foreign DNA in bacteria. Curr. Opin. Microbiol. 2012, 15, 175–181. [Google Scholar] [CrossRef] [PubMed]
- Kotlajich, M.V.; Hron, D.R.; Boudreau, B.A.; Sun, Z.; Lyubchenko, Y.L.; Landick, R. Bridged filaments of histone-like nucleoid structuring protein pause RNA polymerase and aid termination in bacteria. Elife 2015, 2015, 1–24. [Google Scholar] [CrossRef]
- Tobe, T.; Yoshikawa, M.; Mizuno, T.; Sasakawa, C. Transcriptional control of the invasion regulatory gene virB of Shigella flexneri: Activation by virF and repression by H-NS. J. Bacteriol. 1993, 175, 6142–6149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beloin, C.; Dorman, C.J. An extended role for the nucleoid structuring protein H-NS in the virulence gene regulatory cascade of Shigella flexneri. Mol. Microbiol. 2003, 47, 825–838. [Google Scholar] [CrossRef] [PubMed]
- Falconi, M.; Colonna, B.; Prosseda, G.; Micheli, G.; Gualerzi, C.O. Thermoregulation of Shigella and Escherichia coli EIEC pathogenicity. A temperature-dependent structural transition of DNA modulates accessibility of virF promoter to transcriptional repressor H-NS. EMBO J. 1998, 17, 7033–7043. [Google Scholar] [CrossRef] [Green Version]
- Di Martino, M.L.; Micheli, G.; Falconi, M.; Colonna, B.; Prosseda, G. The multifaceted activity of the VirF regulatory protein in the Shigella Lifestyle. Front. Mol. Biosci. 2016, 3, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marman, H.E.; Mey, A.R.; Payne, S.M. Elongation factor P and modifying enzyme PoxA are necessary for virulence of Shigella flexneri. Infect. Immun. 2014, 82, 3612–3621. [Google Scholar] [CrossRef] [Green Version]
- Dorman, C.J. Virulence gene regulation in Shigella. EcoSal Plus 2004, 1, 1–18. [Google Scholar] [CrossRef]
- Durand, J.M.; Björk, G.R.; Kuwae, A.; Yoshikawa, M.; Sasakawa, C. The modified nucleoside 2-methylthio-N6-isopentenyladenosine in tRNA of Shigella flexneri is required for expression of virulence genes. J. Bacteriol. 1997, 179, 5777–5782. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Durand, J.M.; Dagberg, B.; Uhlin, B.E.; Björk, G.R. Transfer RNA modification, temperature and DNA superhelicity have a common target in the regulatory network of the virulence of Shigella flexneri: The expression of the virF gene. Mol. Microbiol. 2000, 35, 924–935. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Martino, M.L.; Romilly, C.; Wagner, E.G.H.; Colonna, B.; Prosseda, G. One gene and two proteins: A leaderless mRNA supports the translation of a shorter form of the Shigella VirF regulator. mBio 2016, 7, 235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tran, C.N.; Giangrossi, M.; Prosseda, G.; Brandi, A.; Di Martino, M.L.; Colonna, B.; Falconi, M. A multifactor regulatory circuit involving H-NS, VirF and an antisense RNA modulates transcription of the virulence gene icsA of Shigella flexneri. Nucleic Acids Res. 2011, 39, 8122–8134. [Google Scholar] [CrossRef] [PubMed]
- Giangrossi, M.; Giuliodori, A.M.; Tran, C.N.; Amici, A.; Marchini, C.; Falconi, M. VirF relieves the transcriptional attenuation of the virulence gene icsA of Shigella flexneri Affecting the icsA mRNA-RnaG complex formation. Front. Microbiol. 2017, 8, 650. [Google Scholar] [CrossRef] [PubMed]
- Taniya, T.; Mitobe, J.; Nakayama, S.I.; Mingshan, Q.; Okuda, K.; Watanabe, H. Determination of the InvE binding site required for expression of IpaB of the Shigella sonnei virulence plasmid: Involvement of a ParB boxA-like sequence. J. Bacteriol. 2003, 185, 5158–5165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kane, K.A.; Dorman, C.J. VirB-mediated positive feedback control of the virulence gene regulatory cascade of Shigella flexneri. J. Bacteriol. 2012, 194, 5264–5273. [Google Scholar] [CrossRef] [Green Version]
- Karney, M.M.A.A.; McKenna, J.A.; Weatherspoon-Griffin, N.; Karabachev, A.D.; Millar, M.E.; Potocek, E.A.; Wing, H.J. Investigating the DNA-binding site for VirB, a key transcriptional regulator of Shigella virulence genes, using an in vivo binding tool. Genes 2019, 10, 149. [Google Scholar] [CrossRef] [Green Version]
- Basta, D.W.; Pew, K.L.; Immak, J.A.; Park, H.S.; Picker, M.A.; Wigley, A.F.; Hensley, C.T.; Pearson, J.S.; Hartland, E.L.; Wing, H.J. Characterization of the ospZ promoter in Shigella flexneri and its regulation by VirB and H-NS. J. Bacteriol. 2013, 195, 2562–2572. [Google Scholar] [CrossRef] [Green Version]
- Weatherspoon-Griffin, N.; Picker, M.A.; Pew, K.L.; Park, H.S.; Ginete, D.R.; Karney, M.M.; Usufzy, P.; Castellanos, M.I.; Duhart, J.C.; Harrison, D.J.; et al. Insights into transcriptional silencing and anti-silencing in Shigella flexneri: A detailed molecular analysis of the icsP virulence locus. Mol. Microbiol. 2018, 108, 505–518. [Google Scholar] [CrossRef] [Green Version]
- Wing, H.J.; Yan, A.W.; Goldman, S.R.; Goldberg, M.B. Regulation of IcsP, the outer membrane protease of the Shigella actin tail assembly protein IcsA, by virulence plasmid regulators VirF and VirB. J. Bacteriol. 2004, 186, 699–705. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turner, E.C.; Dorman, C.J. H-NS antagonism in Shigella flexneri by VirB, a virulence gene transcription regulator that is closely related to plasmid partition factors. J. Bacteriol. 2007, 189, 3403–3413. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McKenna, J.A.; Wing, H.J. The anti-activator of type III secretion, OspD1, is transcriptionally regulated by VirB and H-NS from remote sequences in Shigella flexneri. J. Bacteriol. 2020, 202, e00072-20. [Google Scholar] [CrossRef] [PubMed]
- Le Gall, T.; Mavris, M.; Martino, M.C.; Bernardini, M.L.; Denamur, E.; Parsot, C. Analysis of virulence plasmid gene expression defines three classes of effectors in the type III secretion system of Shigella flexneri. Microbiology 2005, 151, 951–962. [Google Scholar] [CrossRef] [Green Version]
- Gore, A.L.; Payne, S.M. CsrA and Cra influence Shigella flexneri pathogenesis. Infect. Immun. 2010, 78, 4674–4682. [Google Scholar] [CrossRef] [Green Version]
- Vakulskas, C.A.; Potts, A.H.; Babitzke, P.; Ahmer, B.M.M.; Romeo, T. Regulation of bacterial virulence by Csr (Rsm) systems. Microbiol. Mol. Biol. Rev. 2015, 79, 193–224. [Google Scholar] [CrossRef] [Green Version]
- Katsowich, N.; Elbaz, N.; Pal, R.R.; Mills, E.; Kobi, S.; Kahan, T.; Rosenshine, I. Host cell attachment elicits posttranscriptional regulation in infecting enteropathogenic bacteria. Science 2017, 355, 735–739. [Google Scholar] [CrossRef]
- Standish, A.J.; Teh, M.Y.; Tran, E.N.H.; Doyle, M.T.; Baker, P.J.; Morona, R. Unprecedented abundance of protein tyrosine phosphorylation modulates Shigella flexneri virulence. J. Mol. Biol. 2016, 428, 4197–4208. [Google Scholar] [CrossRef]
- Mettert, E.L.; Kiley, P.J. Reassessing the structure and function relationship of the O2 sensing transcription factor FNR. Antioxid. Redox Signal. 2018, 29, 1830–1840. [Google Scholar] [CrossRef]
- Marteyn, B.; West, N.P.; Browning, D.F.; Cole, J.A.; Shaw, J.G.; Palm, F.; Mounier, J.; Prevost, M.-C.; Sansonetti, P.; Tang, C.M. Modulation of Shigella virulence in response to available oxygen in vivo. Nature 2010, 465, 355–358. [Google Scholar] [CrossRef] [Green Version]
- Vergara-Irigaray, M.; Fookes, M.C.; Thomson, N.R.; Tang, C.M. RNA-seq analysis of the influence of anaerobiosis and FNR on Shigella flexneri. BMC Genom. 2014, 15, 438–522. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Penno, C.; Sansonetti, P.; Parsot, C. Frameshifting by transcriptional slippage is involved in production of MxiE, the transcription activator regulated by the activity of the type III secretion apparatus in Shigella flexneri. Mol. Microbiol. 2005, 56, 204–214. [Google Scholar] [CrossRef] [PubMed]
- Demers, B.; Sansonetti, P.J.; Parsot, C. Induction of type III secretion in Shigella flexneri is associated with differential control of transcription of genes encoding secreted proteins. EMBO J. 1998, 17, 2894–2903. [Google Scholar] [CrossRef]
- Kane, C.D.; Schuch, R.; Day, W.A.; Maurelli, A.T. MxiE regulates intracellular expression of factors secreted by the Shigella flexneri 2a type III secretion system. J. Bacteriol. 2002, 184, 4409–4419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mavris, M.; Page, A.-L.; Tournebize, R.; Demers, B.; Sansonetti, P.; Parsot, C. Regulation of transcription by the activity of the Shigella flexneri type III secretion apparatus. Mol. Microbiol. 2002, 43, 1543–1553. [Google Scholar] [CrossRef]
- Parsot, C.; Ageron, E.; Penno, C.; Mavris, M.; Jamoussi, K.; d’Hauteville, H.; Sansonetti, P.; Demers, B. A secreted anti-activator, OspD1, and its chaperone, Spa15, are involved in the control of transcription by the type III secretion apparatus activity in Shigella flexneri. Mol. Microbiol. 2005, 56, 1627–1635. [Google Scholar] [CrossRef] [PubMed]
- Pilonieta, M.C.; Munson, G.P. The chaperone IpgC copurifies with the virulence regulator MxiE. J. Bacteriol. 2008, 190, 2249–2251. [Google Scholar] [CrossRef] [Green Version]
- Mavris, M.; Sansonetti, P.J.; Parsot, C. Identification of the cis-acting site involved in activation of promoters regulated by activity of the type III secretion apparatus in shigella flexneri. J. Bacteriol. 2002, 184, 6751–6759. [Google Scholar] [CrossRef] [Green Version]
- Campbell-Valois, F.-X.; Sansonetti, P.J. Tracking bacterial pathogens with genetically-encoded reporters. FEBS Lett. 2014, 588, 2428–2436. [Google Scholar] [CrossRef] [PubMed]
- Campbell-Valois, F.-X.; Sachse, M.; Sansonetti, P.J.; Parsot, C. Escape of actively secreting Shigella flexnerifrom ATG8/LC3-Positive vacuoles formed during cell-to-cell spread is facilitated by IcsB and VirA. mBio 2015, 6, e02567-14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meijer, B.M.; Jang, S.M.; Guerrera, I.C.; Chhuon, C.; Lipecka, J.; Reisacher, C.; Baleux, F.; Sansonetti, P.J.; Muchardt, C.; Arbibe, L. Threonine eliminylation by bacterial phosphothreonine lyases rapidly causes cross-linking of mitogen-activated protein kinase (MAPK) in live cells. J. Biol. Chem. 2017, 292, 7784–7794. [Google Scholar] [CrossRef] [Green Version]
- Grassart, A.; Malardé, V.; Gobba, S.; Sartori-Rupp, A.; Kerns, J.; Karalis, K.; Marteyn, B.; Sansonetti, P.; Sauvonnet, N. Bioengineered human organ-on-chip reveals intestinal microenvironment and mechanical forces impacting Shigella infection. Cell Host Microbe 2019, 26, 435–444. [Google Scholar] [CrossRef] [PubMed]
- Xu, D.; Liao, C.; Zhang, B.; Tolbert, W.D.; He, W.; Dai, Z.; Zhang, W.; Yuan, W.; Pazgier, M.; Liu, J.; et al. Human enteric α-defensin 5 promotes Shigella infection by enhancing bacterial adhesion and invasion. Immunity 2018, 48, 1233–1244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tinevez, J.-Y.; Arena, E.T.; Anderson, M.; Nigro, G.; Injarabian, L.; André, A.; Ferrari, M.; Campbell-Valois, F.-X.; Devin, A.; Shorte, S.L.; et al. Shigella-mediated oxygen depletion is essential for intestinal mucosa colonization. Nat. Microbiol. 2019, 4, 2001–2009. [Google Scholar] [CrossRef] [PubMed]
- Nigro, G.; Arena, E.T.; Sachse, M.; Moya-Nilges, M.; Marteyn, B.S.; Sansonetti, P.J.; Campbell-Valois, F.-X. Mapping of Shigella flexneri’s tissue distribution and type III secretion apparatus activity during infection of the large intestine of guinea pigs. Pathog. Dis. 2019, 77, ftz054. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmutz, C.; Ahrné, E.; Kasper, C.A.; Tschon, T.; Sorg, I.; Dreier, R.F.; Schmidt, A.; Arrieumerlou, C. Systems-level overview of host protein phosphorylation during Shigella flexneri infection revealed by phosphoproteomics. Mol. Cell Proteom. 2013, 12, 2952–2968. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spinnenhirn, V.; Farhan, H.; Basler, M.; Aichem, A.; Canaan, A.; Groettrup, M. The ubiquitin-like modifier FAT10 decorates autophagy-targeted Salmonella and contributes to Salmonella resistance in mice. J. Cell Sci. 2014, 127, 4883–4893. [Google Scholar] [CrossRef] [Green Version]
- Röder, J.; Hensel, M. Presence of SopE and mode of infection result in increased Salmonella-containing vacuole damage and cytosolic release during host cell infection by Salmonella enterica. Cell. Microbiol. 2020, 22, e13155. [Google Scholar] [CrossRef] [Green Version]
- Finn, C.E.; Chong, A.; Cooper, K.G.; Starr, T.; Steele-Mortimer, O. A second wave of Salmonella T3SS1 activity prolongs the lifespan of infected epithelial cells. PLoS Pathog. 2017, 13, e1006354. [Google Scholar] [CrossRef]
- Morgan, J.M.; Lam, H.N.; Delgado, J.; Luu, J.; Mohammadi, S.; Isberg, R.R.; Wang, H.; Auerbuch, V. An experimental pipeline for initial characterization of bacterial type III secretion system inhibitor mode of action using enteropathogenic yersinia. Front. Cell. Infect. Microbiol. 2018, 8, 1988–2017. [Google Scholar] [CrossRef]
- Davis, K.M.; Mohammadi, S.; Isberg, R.R. Community behavior and spatial regulation within a bacterial microcolony in deep tissue sites serves to protect against host attack. Cell Host Microbe 2015, 17, 21–31. [Google Scholar] [CrossRef] [Green Version]
- Jacobi, C.A.; Roggenkamp, A.; Rakin, A.; Zumbihl, R.; Leitritz, L.; Heesemann, J. In vitro and in vivo expression studies of yopE from Yersinia enterocolitica using the gfp reporter gene. Mol. Microbiol. 1998, 30, 865–882. [Google Scholar] [CrossRef] [PubMed]
- Cabantous, S.; Terwilliger, T.C.; Waldo, G.S. Protein tagging and detection with engineered self-assembling fragments of green fluorescent protein. Nat. Biotechnol. 2005, 23, 102–107. [Google Scholar] [CrossRef] [PubMed]
- Van Engelenburg, S.B.; Palmer, A.E. Imaging type-III secretion reveals dynamics and spatial segregation of Salmonella effectors. Nat. Methods 2010, 7, 325–330. [Google Scholar] [CrossRef] [PubMed]
- Jo, S.H.; Lee, J.; Park, E.; Kim, D.W.; Lee, D.H.; Ryu, C.M.; Choi, D.; Park, J.M. A human pathogenic bacterium Shigella proliferates in plants through adoption of type III effectors for shigellosis. Plant. Cell Environ. 2019, 42, 2962–2978. [Google Scholar] [CrossRef] [PubMed]
- Gawthorne, J.A.; Audry, L.; McQuitty, C.; Dean, P.; Christie, J.M.; Enninga, J.; Roe, A.J. Visualizing the translocation and localization of bacterial type III effector proteins by using a genetically encoded reporter system. Appl. Environ. Microbiol. 2016, 82, 2700–2708. [Google Scholar] [CrossRef] [Green Version]
- Peron-Cane, C.; Fernandez, J.-C.; Leblanc, J.; Wingertsmann, L.; Gautier, A.; Desprat, N.; Lebreton, A. Fluorescent secreted bacterial effectors reveal active intravacuolar proliferation of Listeria monocytogenes in epithelial cells. PLoS Pathog. 2020, 16, e1009001. [Google Scholar] [CrossRef]
- Göser, V.; Kommnick, C.; Liss, V.; Hensel, M. Self-Labeling enzyme tags for analyses of translocation of type III secretion system effector proteins. mBio 2019, 10, 1306. [Google Scholar] [CrossRef] [Green Version]
- Enninga, J.; Mounier, J.; Sansonetti, P.; Tran Van Nhieu, G. Secretion of type III effectors into host cells in real time. Nat. Methods 2005, 2, 959–965. [Google Scholar] [CrossRef]
- Praveschotinunt, P.; Dorval Courchesne, N.M.; Den Hartog, I.; Lu, C.; Kim, J.J.; Nguyen, P.Q.; Joshi, N.S. Tracking of engineered bacteria in vivo using nonstandard amino acid incorporation. ACS Synth. Biol. 2018, 7, 1640–1650. [Google Scholar] [CrossRef]
- Ambrosi, C.; Pompili, M.; Scribano, D.; Limongi, D.; Petrucca, A.; Cannavacciuolo, S.; Schippa, S.; Zagaglia, C.; Grossi, M.; Nicoletti, M. The Shigella flexneri OspB effector: An early immunomodulator. Int. J. Med. Microbiol. 2015, 305, 75–84. [Google Scholar] [CrossRef] [PubMed]
- Baxt, L.A.; Goldberg, M.B. Host and bacterial proteins that repress recruitment of LC3 to Shigella early during infection. PLoS ONE 2014, 9, e94653. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanenbaum, M.E.; Gilbert, L.A.; Qi, L.S.; Weissman, J.S.; Vale, R.D. A protein-tagging system for signal amplification in gene expression and fluorescence imaging. Cell 2014, 159, 635–646. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, W.; Zhou, Y.; Peng, T.; Zhou, P.; Ding, X.; Li, Z.; Zhong, H.; Xu, Y.; Chen, S.; Hang, H.C.; et al. Nε-fatty acylation of multiple membrane-associated proteins by Shigella IcsB effector to modulate host function. Nat. Microbiol. 2018, 996–1009. [Google Scholar] [CrossRef]
- Li, P.; Jiang, W.; Yu, Q.; Liu, W.; Zhou, P.; Li, J.; Xu, J.; Xu, B.; Wang, F.; Shao, F. Ubiquitination and degradation of GBPs by a Shigella effector to suppress host defence. Nature 2017, 551, 378–383. [Google Scholar] [CrossRef]
- Pinaud, L.; Samassa, F.; Porat, Z.; Ferrari, M.L.; Belotserkovsky, I.; Parsot, C.; Sansonetti, P.J.; Campbell-Valois, F.-X.; Phalipon, A. Injection of T3SS effectors not resulting in invasion is the main targeting mechanism of Shigella toward human lymphocytes. Proc. Natl. Acad. Sci. USA 2017, 13, 201707098. [Google Scholar] [CrossRef] [Green Version]
- Belotserkovsky, I.; Brunner, K.; Pinaud, L.; Rouvinski, A.; Dellarole, M.; Baron, B.; Dubey, G.; Samassa, F.; Parsot, C.; Sansonetti, P.; et al. Glycan-Glycan interaction determines shigella tropism toward human T lymphocytes. mBio 2018, 9, 858. [Google Scholar] [CrossRef] [Green Version]
- Konradt, C.; Frigimelica, E.; Nothelfer, K.; Puhar, A.; Salgado-Pabon, W.; di Bartolo, V.; Scott-Algara, D.; Rodrigues, C.D.; Sansonetti, P.J.; Phalipon, A. The Shigella flexneri type three secretion system effector IpgD inhibits T cell migration by manipulating host phosphoinositide metabolism. Cell Host Microbe 2011, 9, 263–272. [Google Scholar] [CrossRef]
- O’Boyle, N.; Connolly, J.P.R.; Roe, A.J. Tracking elusive cargo: Illuminating spatio-temporal Type 3 effector protein dynamics using reporters. Cell. Microbiol. 2018, 20, e12797. [Google Scholar] [CrossRef] [Green Version]
- Pinaud, L.; Sansonetti, P.J.; Phalipon, A. Host cell targeting by enteropathogenic bacteria T3SS effectors. Trends Microbiol. 2018, 26, 266–283. [Google Scholar] [CrossRef]
- Bernardini, M.L.; Mounier, J.; d’Hauteville, H.; Coquis-Rondon, M.; Sansonetti, P.J. Identification of icsA, a plasmid locus of Shigella flexneri that governs bacterial intra- and intercellular spread through interaction with F-actin. Proc. Natl. Acad. Sci. USA 1989, 86, 3867–3871. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- High, N.; Mounier, J.; Prevost, M.C.; Sansonetti, P.J. IpaB of Shigella flexneri causes entry into epithelial cells and escape from the phagocytic vacuole. EMBO J. 1992, 11, 1991–1999. [Google Scholar] [CrossRef] [PubMed]
- Page, A.L.; Ohayon, H.; Sansonetti, P.J.; Parsot, C. The secreted IpaB and IpaC invasins and their cytoplasmic chaperone IpgC are required for intercellular dissemination of Shigella flexneri. Cell. Microbiol. 1999, 1, 183–193. [Google Scholar] [CrossRef] [PubMed]
- Schuch, R.; Sandlin, R.C.; Maurelli, A.T. A system for identifying post-invasion functions of invasion genes: Requirements for the Mxi-Spa type III secretion pathway of Shigella flexneri in intercellular dissemination. Mol. Microbiol. 1999, 34, 675–689. [Google Scholar] [CrossRef] [PubMed]
- Duncan-Lowey, J.K.; Wiscovitch, A.L.; Wood, T.E.; Goldberg, M.B.; Russo, B.C. Shigella flexneri disruption of cellular tension promotes intercellular spread. Cell Rep. 2020, 33, 108409. [Google Scholar] [CrossRef] [PubMed]
- Du, J.; Reeves, A.Z.; Klein, J.A.; Twedt, D.J.; Knodler, L.A.; Lesser, C.F. The type III secretion system apparatus determines the intracellular niche of bacterial pathogens. Proc. Natl. Acad. Sci. USA 2016, 113, 4794–4799. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mellouk, N.; Weiner, A.; Aulner, N.; Schmitt, C.; Elbaum, M.; Shorte, S.L.; Danckaert, A.; Enninga, J. Shigella subverts the host recycling compartment to rupture its vacuole. Cell Host Microbe 2014, 16, 517–530. [Google Scholar] [CrossRef] [Green Version]
- Huang, J.; Brumell, J.H. Bacteria-autophagy interplay: A battle for survival. Nat. Rev. Microbiol. 2014, 12, 101–114. [Google Scholar] [CrossRef]
- Perrin, A.J.; Jiang, X.; Birmingham, C.L.; So, N.S.; Brumell, J.H. Recognition of bacteria in the cytosol of Mammalian cells by the ubiquitin system. Curr. Biol. 2004, 14, 806–811. [Google Scholar] [CrossRef] [Green Version]
- Thurston, T.L.; Ryzhakov, G.; Bloor, S.; von Muhlinen, N.; Randow, F. The TBK1 adaptor and autophagy receptor NDP52 restricts the proliferation of ubiquitin-coated bacteria. Nat. Immunol. 2009, 10, 1215–1221. [Google Scholar] [CrossRef]
- Thurston, T.L.; Wandel, M.P.; von Muhlinen, N.; Foeglein, A.; Randow, F. Galectin 8 targets damaged vesicles for autophagy to defend cells against bacterial invasion. Nature 2012, 482, 414–418. [Google Scholar] [CrossRef] [PubMed]
- Tattoli, I.; Sorbara, M.T.; Vuckovic, D.; Ling, A.; Soares, F.; Carneiro, L.A.M.; Yang, C.; Emili, A.; Philpott, D.J.; Girardin, S.E. Amino acid starvation induced by invasive bacterial pathogens triggers an innate host defense program. Cell Host Microbe 2012, 11, 563–575. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dupont, N.; Lacas-Gervais, S.; Bertout, J.; Paz, I.; Freche, B.; van Nhieu, G.T.; van der Goot, F.G.; Sansonetti, P.J.; Lafont, F. Shigella phagocytic vacuolar membrane remnants participate in the cellular response to pathogen invasion and are regulated by autophagy. Cell Host Microbe 2009, 6, 137–149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paz, I.; Sachse, M.; Dupont, N.; Mounier, J.; Cederfur, C.; Enninga, J.; Leffler, H.; Poirier, F.; Prevost, M.-C.; Lafont, F.; et al. Galectin-3, a marker for vacuole lysis by invasive pathogens. Cell. Microbiol. 2010, 12, 530–544. [Google Scholar] [CrossRef]
- Ogawa, M.; Yoshimori, T.; Suzuki, T.; Sagara, H.; Mizushima, N.; Sasakawa, C. Escape of intracellular Shigella from autophagy. Science 2005, 307, 727–731. [Google Scholar] [CrossRef]
- Mostowy, S.; Bonazzi, M.; Hamon, M.A.; Tham, T.N.; Mallet, A.; Lelek, M.; Gouin, E.; Demangel, C.; Brosch, R.; Zimmer, C.; et al. Entrapment of intracytosolic bacteria by septin cage-like structures. Cell Host Microbe 2010, 8, 433–444. [Google Scholar] [CrossRef]
- Krokowski, S.; Lobato-Márquez, D.; Chastanet, A.; Pereira, P.M.; Angelis, D.; Galea, D.; Larrouy-Maumus, G.; Henriques, R.; Spiliotis, E.T.; Carballido-López, R.; et al. Septins recognize and entrap dividing bacterial cells for delivery to lysosomes. Cell Host Microbe 2018, 24, 866–874. [Google Scholar] [CrossRef] [Green Version]
- Weddle, E.; Agaisse, H.J.I. Immunity spatial, temporal, and functional assessment of LC3-dependent autophagy in Shigella flexneri dissemination. Infect. Immun. 2018, 86, e00134-18. [Google Scholar] [CrossRef] [Green Version]
- Leung, Y.; Ally, S.; Goldberg, M.B. Bacterial actin assembly requires toca-1 to relieve N-wasp autoinhibition. Cell Host Microbe 2008, 3, 39–47. [Google Scholar] [CrossRef] [Green Version]
- Miller, K.A.; Garza-Mayers, A.C.; Leung, Y.; Goldberg, M.B. Identification of interactions among host and bacterial proteins and evaluation of their role early during Shigella flexneri infection. Microbiology 2018, 46, 251–311. [Google Scholar] [CrossRef]
- Kühn, S.; Bergqvist, J.; Gil, M.; Valenzuela, C.; Barrio, L.; Lebreton, S.; Zurzolo, C.; Enninga, J. Actin assembly around the Shigella-Containing vacuole promotes successful infection. Cell Rep. 2020, 31, 107638. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Huang, C.; Yin, L.; Wan, M.; Wang, X.; Li, L.; Liu, Y.; Wang, Z.; Fu, P.; Zhang, N.; et al. Nε-Fatty acylation of Rho GTPases by a MARTX toxin effector. Science 2017, 358, 528–531. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pei, J.; Grishin, N.V. The Rho GTPase inactivation domain in Vibrio cholerae MARTX toxin has a circularly permuted papain-like thiol protease fold. Proteins 2009, 77, 413–419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rathman, M.; Jouirhi, N.; Allaoui, A.; Sansonetti, P.; Parsot, C.; Tran Van Nhieu, G. The development of a FACS-based strategy for the isolation of Shigella flexneri mutants that are deficient in intercellular spread. Mol. Microbiol. 2000, 35, 974–990. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, Y.; Zhou, P.; Cheng, S.; Lu, Q.; Nowak, K.; Hopp, A.-K.; Li, L.; Shi, X.; Zhou, Z.; Gao, W.; et al. A bacterial effector reveals the V-ATPase-ATG16L1 Axis that initiates xenophagy. Cell 2019, 178, 552–566. [Google Scholar] [CrossRef]
- Dong, N.; Zhu, Y.; Lu, Q.; Hu, L.; Zheng, Y.; Shao, F. Structurally distinct bacterial TBC-like GAPs link Arf GTPase to Rab1 inactivation to counteract host defenses. Cell 2012, 150, 1029–1041. [Google Scholar] [CrossRef] [Green Version]
- Zoppino, F.C.M.; Militello, R.D.; Slavin, I.; Alvarez, C.; Colombo, M.I. Autophagosome formation depends on the small GTPase Rab1 and functional ER exit sites. Traffic 2010, 11, 1246–1261. [Google Scholar] [CrossRef]
- Kakuta, S.; Yamaguchi, J.; Suzuki, C.; Sasaki, M.; Kazuno, S.; Uchiyama, Y. Small GTPase Rab1B is associated with ATG9A vesicles and regulates autophagosome formation. FASEB J. 2017, 31, 3757–3773. [Google Scholar] [CrossRef] [Green Version]
- Wandel, M.P.; Pathe, C.; Werner, E.I.; Ellison, C.J.; Boyle, K.B.; von der Malsburg, A.; Rohde, J.; Randow, F. GBPs inhibit motility of shigella flexneri but are targeted for degradation by the bacterial ubiquitin ligase IpaH9.8. Cell Host Microbe 2017, 22, 507–518. [Google Scholar] [CrossRef] [Green Version]
- Piro, A.S.; Hernandez, D.; Luoma, S.; Feeley, E.M.; Finethy, R.; Yirga, A.; Frickel, E.M.; Lesser, C.F.; Coers, J. Detection of cytosolic Shigella flexneri via a C-terminal triple-arginine motif of GBP1 inhibits actin-based motility. mBio 2017, 8, e01979-17. [Google Scholar] [CrossRef] [Green Version]
- Ji, C.; Du, S.; Li, P.; Zhu, Q.; Yang, X.; Long, C.; Yu, J.; Shao, F.; Xiao, J. Structural mechanism for guanylate-binding proteins (GBPs) targeting by the Shigella E3 ligase IpaH9.8. PLoS Pathog. 2019, 15, e1007876. [Google Scholar] [CrossRef] [PubMed]
- Wandel, M.P.; Kim, B.-H.; Park, E.-S.; Boyle, K.B.; Nayak, K.; Lagrange, B.; Herod, A.; Henry, T.; Zilbauer, M.; Rohde, J.; et al. Guanylate-binding proteins convert cytosolic bacteria into caspase-4 signaling platforms. Nat. Immunol. 2020, 21, 880–891. [Google Scholar] [CrossRef] [PubMed]
- Sorbara, M.T.; Foerster, E.G.; Tsalikis, J.; Abdel-Nour, M.; Mangiapane, J.; Sirluck-Schroeder, I.; Tattoli, I.; van Dalen, R.; Isenman, D.E.; Rohde, J.R.; et al. Complement C3 drives autophagy-dependent restriction of cyto-invasive bacteria. Cell Host Microbe 2018, 23, 644–652. [Google Scholar] [CrossRef] [PubMed] [Green Version]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bajunaid, W.; Haidar-Ahmad, N.; Kottarampatel, A.H.; Ourida Manigat, F.; Silué, N.; F. Tchagang, C.; Tomaro, K.; Campbell-Valois, F.-X. The T3SS of Shigella: Expression, Structure, Function, and Role in Vacuole Escape. Microorganisms 2020, 8, 1933. https://doi.org/10.3390/microorganisms8121933
Bajunaid W, Haidar-Ahmad N, Kottarampatel AH, Ourida Manigat F, Silué N, F. Tchagang C, Tomaro K, Campbell-Valois F-X. The T3SS of Shigella: Expression, Structure, Function, and Role in Vacuole Escape. Microorganisms. 2020; 8(12):1933. https://doi.org/10.3390/microorganisms8121933
Chicago/Turabian StyleBajunaid, Waad, Nathaline Haidar-Ahmad, Anwer Hasil Kottarampatel, France Ourida Manigat, Navoun Silué, Caetanie F. Tchagang, Kyle Tomaro, and François-Xavier Campbell-Valois. 2020. "The T3SS of Shigella: Expression, Structure, Function, and Role in Vacuole Escape" Microorganisms 8, no. 12: 1933. https://doi.org/10.3390/microorganisms8121933
APA StyleBajunaid, W., Haidar-Ahmad, N., Kottarampatel, A. H., Ourida Manigat, F., Silué, N., F. Tchagang, C., Tomaro, K., & Campbell-Valois, F.-X. (2020). The T3SS of Shigella: Expression, Structure, Function, and Role in Vacuole Escape. Microorganisms, 8(12), 1933. https://doi.org/10.3390/microorganisms8121933