Untapped Potentials of Endophytic Fungi: A Review of Novel Bioactive Compounds with Biological Applications


Abstract
:1. Introduction
2. Biological Properties of Novel Bioactive Compounds from Endophytic Fungi
2.1. Bioactive Compounds as an Alternative Antibacterial Agents
2.2. Bioactive Compounds as an Alternative to Antifungal Agents
2.3. Bioactive Compounds for Treating Cancer Cells (Anticancer Activity)
2.4. Bioactive Compounds as a Potential Antioxidant Agent
2.5. Bioactive Compounds for Treating Infectious Parasites
2.6. Bioactive Compounds with the Potential of Serving as Immunosuppressive Drugs
2.7. Bioactive Compounds with Antiviral Properties
2.8. Bioactive Compounds as Potential Antitubercular Drugs
3. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ayukekbong, J.A.; Ntemgwa, M.; Atabe, A.N. The threat of antimicrobial resistance in developing countries: Causes and control strategies. Antimicrob. Resist. Infect. Control. 2017, 6, 47. [Google Scholar] [CrossRef] [PubMed]
- Dadgostar, P. Antimicrobial Resistance: Implications and Costs. Infect. Drug Resist. 2019, 12, 3903–3910. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vasan, N.; Baselga, J.; Hyman, D.M. A view on drug resistance in cancer. Nature 2019, 575, 299–309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prestinaci, F.; Pezzotti, P.; Pantosti, A. Antimicrobial resistance: A global multifaceted phenomenon. Pathog. Glob. Health 2015, 109, 309–318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- World Health Organization (WHO). Antimicrobial Resistance: Global Report on Surveillance; WHO: Geneva, Switzerland, 2014. [Google Scholar]
- World Health Organization (WHO). Traditional Medicine. World Health Organization. 2008. Available online: http://www.who.int/mediacentre/factsheets/2003/fs134/en/ (accessed on 11 September 2015).
- Mukherjee, P.K. Trichoderma–Genetics, Genomics and Beyond. Indian Phytopathol. 2015, 68, 1–7. [Google Scholar]
- Li, F.; Weng, J. Demystifying traditional herbal medicine with modern approach. Nat. Plants 2017, 3, 17109. [Google Scholar] [CrossRef]
- Othman, L.; Sleiman, A.; Abdel-Massih, R.M. Antimicrobial activity of polyphenols and alkaloids in middle eastern plants. Front. Microbiol. 2019, 10, 911. [Google Scholar] [CrossRef]
- Rodriguez, R.; Redman, R. More than 400 million years of evolution and some plants still can’t make it on their own: Plant stress tolerance via fungal symbiosis. J. Exp. Bot. 2008, 59, 1109–1123. [Google Scholar] [CrossRef]
- Hawksworth, D.L. The fungal dimension of biodiversity: Magnitude, significance, and conservation. Mycol. Res. 1991, 95, 641–655. [Google Scholar] [CrossRef]
- Kaul, S.; Gupta, S.; Ahmed, M.; Dhar, M.K. Endophytic fungi from medicinal plants: A treasure hunt for bioactive metabolites. Phytochem. Rev. 2012, 11, 487–505. [Google Scholar] [CrossRef]
- Rabiey, M.; Hailey, L.E.; Roy, S.R.; Grenz, K.; Al-Zadjali, M.A.S.; Barrett, G.A.; Jackson, R.W. Endophytes vs tree pathogens and pests: Can they be used as biological control agents to improve tree health? Eur. J. Plant Pathol. 2019, 155, 711–729. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Ren, L.; Li, C.; Gao, C.; Liu, X.; Wang, M.; Luo, L. Effects of endophytic fungi diversity in different coniferous species on the colonization of Sirex noctilio (Hymenoptera: Siricidae). Sci. Rep. 2019, 9, 5077. [Google Scholar] [CrossRef] [PubMed]
- Khare, E.; Mishra, J.; Arora, N.K. Multifaceted interactions between endophytes and plant: Developments and prospects. Front. Microbiol. 2018, 9, 2732. [Google Scholar] [CrossRef] [PubMed]
- Eid, A.M.; Salim, S.S.; Hassan, S.E.D.; Ismail, M.A.; Fouda, A. Role of endophytes in plant health and abiotic stress management. In Microbiome in Plant Health and Disease; Springer: Singapore, 2019; pp. 119–144. [Google Scholar]
- Tan, S.Y.; Tatsumura, Y. Alexander Fleming (1881–1955): Discoverer of penicillin. Singap. Med. J. 2015, 56, 366–367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, L.; Chen, L. Progress in research on paclitaxel and tumor immunotherapy. Cell Mol. Biol. Lett. 2019, 24, 40. [Google Scholar] [CrossRef] [Green Version]
- Fadiji, A.E.; Babalola, O.O. Elucidating mechanisms of endophytes used in plant protection and other bioactivities with multifunctional prospects. Front. Bioeng. Biotechnol. 2020, 8, 467. [Google Scholar] [CrossRef]
- Toghueo, R.M.K. Bioprospecting endophytic fungi from Fusarium genus as sources of bioactive metabolites. Mycology 2020, 11, 1–21. [Google Scholar] [CrossRef] [Green Version]
- Sengupta, S.; Chattopadhyay, M.K.; Grossart, H.P. The multifaceted roles of antibiotics and antibiotic resistance in nature. Front. Microbiol. 2013, 4, 47. [Google Scholar] [CrossRef] [Green Version]
- Ventola, C.L. The antibiotic resistance crisis: Part 1: Causes and treats. Pharm. Ther. 2015, 40, 277. [Google Scholar]
- Centers for Disease Control and Prevention, Office of Infectious Disease. Antibiotic Resistance Threats in the United States. Available online: http://www.cdc.gov/drugresistance/threat-report-2013 (accessed on 21 June 2020).
- World Health Organization (WHO). Fact Sheets. Antibiotic Resistance. Available online: http://www.who.int/news-room/fact-sheet/detail/antibiotic-resistance (accessed on 21 June 2020).
- Ravindra, P.A.; Suneel, K.; Sardul, S.S. Endophytic mycoflora as a source of biotherapeutic compounds for disease treatment. J. Appl. Pharm. Sci. 2016, 6, 242–254. [Google Scholar]
- Pavithra, G.; Sumant, B.; Meenakshi, R.; Seweta, S. Role of Endophytic Microbes against Plant Pathogens. Asian J. Plant Sci. 2020, 19, 54–62. [Google Scholar]
- Tirsit, T. Pharmaceutical effects of bioactive compounds from endophytes. Int. J. Innov. Pharm. Sci. Res. 2018, 1, 1–13. [Google Scholar]
- Chatterjee, S.; Ghosh, R.; Mandal, N.C. Production of bioactive compounds with bactericidal and antioxidant potential by endophytic fungus Alternaria alternata AE1 isolated from Azadirachta indica A. Juss. PLoS ONE 2019, 14, e0214744. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maria do Carmo, M.; Carvalho, C.M.; Faria, F.S.; Nobreza, A.M.; Thiago, M. Antibacterial Activity of Phytochemical extracts and endophytic fungi of Carapa guianensis against Enterococcus faecalis in endodontic infections an in vitro study. Open Dent. J. 2019, 13, 249–254. [Google Scholar]
- Manganyi, M.C.; Tchatchouang, C.D.K.; Regnier, T.; Bezuidenhout, C.C.; Ateba, C.N. Bioactive compound produced by endophytic fungi isolated from Pelargonium sidoides against selected bacteria of clinical importance. Mycobiology 2019, 47, 335–339. [Google Scholar] [CrossRef] [Green Version]
- Varkey, S.; Anith, K.N.; Narayana, R.; Aswini, S. A consortium of rhizobacteria and fungal endophyte suppress the root-knot nematode parasite in tomato. Rhizosphere 2018, 5, 38–42. [Google Scholar] [CrossRef]
- Kjer, J.; Wray, V.; Edrada-Ebel, R.; Ebel, R.; Pretsch, A.; Lin, W.H.; Proksch, P. Xanalteric acids I and II and related phenolic compounds from an endophytic Alternaria sp. isolated from the mangrove plant Sonneratia alba. J. Nat. Prod. 2009, 72, 2053–2057. [Google Scholar] [CrossRef]
- Uzma, F.; Mohan, C.D.; Siddaiah, C.N.; Chowdappa, S. Endophytic fungi: Promising source of novel bioactive compounds. In Advances in Endophytic Fungal Research; Springer: Cham, Switzerland, 2019; pp. 243–265. [Google Scholar]
- Ding, Z.; Tao, T.; Wang, L.; Zhao, Y.; Huang, H.; Zhang, D.; Liu, M.; Wang, Z.; Han, J. Bioprospecting of Novel and Bioactive Metabolites from Endophytic Fungi Isolated from Rubber Tree Ficus elastica Leaves. Microbiol. Biotechnol. 2019, 29, 731–738. [Google Scholar] [CrossRef] [Green Version]
- Findlay, J.A.; Li, G.; Johnson, J.A. Bioactive compounds from an endophytic fungus from eastern larch (Larix laricina) needles. Can. J. Chem. 1997, 75, 716–719. [Google Scholar] [CrossRef]
- Hawksworth, D.L.; Luecking, R. Fungal diversity revisited: 2.2 to 3.8 million species. Microbiol. Spectr. 2017, 5, 0052-2016. [Google Scholar]
- Almeida, F.; Rodrigues, M.L.; Coelho, C. The Still Underestimated Problem of Fungal Diseases Worldwide. Front. Microbiol. 2019, 10, 214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giacomazzi, J.; Baethgen, L.; Carneiro, L.C.; Millington, M.A.; Denning, D.W.; Colombo, A.L.; Pasqualotto, A.C. The burden of serious human fungal infections in Brazil. Mycoses 2015, 59, 145–150. [Google Scholar] [CrossRef] [PubMed]
- Peláez, F.; Cabello, A.; Platas, G.; Díez, M.T.; del Val, A.G.; Basilio, A.; Martán, I.; Vicente, F.; Bills, G.F.; Giacobbe, R.A.; et al. The discovery of enfumafungin, a novel antifungal compound produced by an endophytic Hormonema species biological activity and taxonomy of the producing organisms. Syst. Appl. Microbiol. 2000, 23, 333–343. [Google Scholar] [CrossRef]
- Chepkirui, C.; Richter, C.; Matasyoh, J.C.; Stadler, M. Monochlorinated calocerins A-D and 9-oxostrobilurin derivatives from the basidiomycete Favolaschia calocera. Phytochemistry 2016, 132, 95–101. [Google Scholar] [CrossRef] [PubMed]
- Haryani, Y.; Hilma, R.; Delfira, N.; Martalinda, T.; Puspita, F.; Friska, A.; Juwita, D.; Farniga, A.; Ardi, F. Potential Antibacterial Activity of Endophytic Fungi Penicillium sp. and Trichoderma sp. Derived From Mangrove Ceriops Tagal (Perr.) CB Robb and Bruguiera sp. J. Phys. Conf. Ser. 2019, 1351, 012100. [Google Scholar] [CrossRef]
- Li, E.; Jiang, L.; Guo, L.; Zhang, H.; Che, Y. Pestalachlorides A-C, antifungal metabolites from the plant endophytic fungus Pestalotiopsis adusta. Bioorg. Med. Chem. 2008, 16, 7894–7899. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, H.T.; Choi, S.; Kim, S.; Lee, J.H.; Park, A.R.; Yu, N.H.; Yoon, H.; Bae, C.H.; Yeo, J.H.; Choi, G.J.; et al. The Hsp90 Inhibitor, Monorden, Is a Promising Lead Compound for the Development of Novel Fungicides. Front. Plant Sci. 2020, 11, 371. [Google Scholar] [CrossRef] [PubMed]
- Ma, K.L.; Wei, W.J.; Li, H.Y.; Wang, L.D.; Dong, S.H.; Gao, K. Phomotide A, a novel polyketide, from the endophytic fungus Phomopsis sp. CFS42. Tetrahedron Lett. 2020, 61, 151468. [Google Scholar] [CrossRef]
- World Health Organization (WHO). Fact Sheets. Cancer. Available online: https://www.who.int/news-room/fact-sheets/detail/cancer (accessed on 24 June 2020).
- Batra, H.; Pawar, S.; Bahl, D. Curcumin in combination with anti-cancer drugs: A nanomedicine review. Pharmacol. Res. 2019, 139, 91–105. [Google Scholar] [CrossRef]
- Peng, C.; Xu, J.; Yu, M.; Ning, X.; Huang, Y.; Du, B.; Hernandez, E.; Kapur, P.; Hsieh, J.T.; Zheng, J. Tuning the in-Vivo Transport of Anticancer Drugs Using Renal-Clearable Gold Nanoparticles. Angew. Chem. 2019, 131, 8567–8571. [Google Scholar]
- Tewari, D.; Rawat, P.; Singh, P.K. Adverse drug reactions of anticancer drugs derived from natural sources. Food Chem. Toxicol. 2019, 123, 522–535. [Google Scholar] [CrossRef] [PubMed]
- Singh, M.; Kumar, A.; Singh, R.; Pandey, K.D. Endophytic bacteria: A new source of bioactive compounds. 3 Biotech 2017, 7, 315. [Google Scholar] [CrossRef]
- Das, A.; Rahman, M.I.; Ferdous, A.S.; Amin, A.; Rahman, M.M.; Nahar, N.; Uddin, M.A.; Islam, M.R.; Khan, H. An endophytic Basidiomycete, Grammothele lineata, isolated from Corchorus olitorius, produces paclitaxel that shows cytotoxicity. PLoS ONE 2017, 12, e0178612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qiao, W.; Tang, T.; Ling, F. Comparative transcriptome analysis of a taxol-producing endophytic fungus, Aspergillus aculeatinus Tax-6, and its mutant strain. Sci. Rep. 2020, 10, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Gill, H.; Vasundhara, M. Isolation of taxol producing endophytic fungus Alternaria brassicicola from non-Taxus medicinal plant Terminalia arjuna. World J. Microbiol. Biotechnol. 2019, 35, 74. [Google Scholar] [CrossRef] [PubMed]
- El-Bialy, H.A.; El-Bastawisy, H.S. Elicitors stimulate paclitaxel production by endophytic fungi isolated from ecologically altered Taxus baccata. J. Radiat. Res. Appl. 2020, 13, 79–87. [Google Scholar] [CrossRef] [Green Version]
- Stierle, A.; Strobel, G.; Stierle, D. Taxol and taxane production by Taxomyces andreanae, an endophytic fungus of Pacific yew. Science 1993, 260, 214–226. [Google Scholar] [CrossRef]
- Kumar, G.; Chandra, P.; Choudhary, M. Endophytic fungi: A potential source of bioactive compounds. Chem. Sci. Rev. Lett. 2017, 6, 2373–2381. [Google Scholar]
- Shweta, S.; Zuehlke, S.; Ramesha, B.T. Endophytic fungal strains of Fusarium solani, from Apodytes dimidiate E. Mey. ex Arn (Icacinaceae) produce camptothecin, 10-hydroxycamptothecin and 9-methoxycamptothecin. Phytochemistry 2010, 71, 117–122. [Google Scholar] [CrossRef]
- Wani, M.C.; Taylor, H.L.; Wall, M.E.; Coggon, P.; McPhail, A. Plant antitumor agents. VI. The isolation and structure of taxol, a novel anti leukemic and antitumor agent from Taxus brevifolia. J. Am. Chem. Soc. 1971, 93, 2325–2327. [Google Scholar] [CrossRef]
- Li, X.; Tian, Y.; Yang, S.-X.; Zhang, Y.-M.; Qin, J.-C. Cytotoxic azaphilone alkaloids from Chaetomium globosum TY1. Bioorg. Med. Chem. Lett. 2013, 23, 2945–2947. [Google Scholar] [CrossRef] [PubMed]
- Rana, K.L.; Kour, D.; Kaur, T.; Devi, R.; Negi, C.; Yadav, A.N.; Yadav, N.; Singh, K.; Saxena, A.K. Endophytic fungi from medicinal plants: Biodiversity and biotechnological applications. In Microbial Endophytes; Woodhead Publishing: Cambridge, UK, 2020; pp. 273–305. [Google Scholar]
- Wagenaar, M.M.; Corwin, J.; Strobel, G.; Clardy, J. Three new cytochalasins produced by an endophytic fungus in the genus Rhinocladiella. J. Nat. Prod. 2000, 63, 1692–1695. [Google Scholar] [CrossRef] [PubMed]
- Torres-Mendoza, D.; Ortega, H.E.; Cubilla-Rios, L. Patents on endophytic fungi related to secondary metabolites and biotransformation applications. J. Fungi 2020, 6, 58. [Google Scholar] [CrossRef] [PubMed]
- Puri, S.C.; Nazir, A.; Chawla, R.; Arora, R.; Riyaz-ul-Hasan, S.; Amna, T.; Ahmed, B.; Verma, V.; Singh, S.; Sagar, R.; et al. The endophytic fungus Trametes hirsuta as a novel alternative source of podophyllotoxin and related aryl tetralin lignans. J. Biotechnol. 2006, 122, 494–510. [Google Scholar] [CrossRef]
- Tidke, S.A.; Kumar, R.K.; Ramakrishna, D.; Kiran, S.; Kosturkova, G.; Gokare, R.A. Current understanding of endophytes: Their relevance, importance, and industrial potentials. J. Biotechnol. Biochem. 2017, 3, 43–59. [Google Scholar] [CrossRef]
- Baker, S.; Satish, S. Endophytes: Natural warehouse of bioactive compounds. Drug Invent. Today 2012, 4, 548–553. [Google Scholar]
- Sarika, G.; Amruta, N.; Kandikattu, H.K.; Basavaraju, G.V.; Suma, H.K.; Manjunath, B.L.; Sravani, C.H. Chemical profiling of camptothecin and methoxy camptothecin in Nothapodytes nimmoniana Grah.(Mabb.) during seed development, seed germination and their effects on seed-borne pathogens. S. Afr. J. Bot. 2019, 123, 113–123. [Google Scholar] [CrossRef]
- Neelam, K.A.; Sharma, K.K. Phenylpropanoids and its derivatives: Biological activities and its role in food, pharmaceutical and cosmetic industries. Crit. Rev. Food Sci. Nutr. 2019, 1–21. [Google Scholar]
- Cai, R.; Jiang, H.; Zang, Z.; Li, C.; She, Z. New benzofuranoids and phenylpropanoids from the mangrove endophytic fungus, Aspergillus sp. ZJ-68. Mar. Drugs 2019, 17, 478. [Google Scholar] [CrossRef] [Green Version]
- Adorisio, S.; Fierabracci, A.; Muscari, I.; Liberati, A.M.; Cannarile, L.; Thuy, T.T.; Sung, T.V.; Sohrab, H.; Hasan, C.M.; Ayroldi, E.; et al. Fusarubin and anhydrofusarubin isolated from a Cladosporium species inhibit cell growth in human cancer cell lines. Toxins 2019, 11, 503. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.W.; Zhang, Y.L.; Lin, F.C.; Hu, Y.Z.; Zhang, C.L. Natural products with antitumor activity from endophytic fungi. Mini Rev. Med. Chem. 2011, 11, 1056–1074. [Google Scholar] [CrossRef] [PubMed]
- Elochukwui, C. Generation and reaction of free radicals in the human body: A major cause of aging and chronic degenerative diseases. EC Nutr. 2015, 1, 132–136. [Google Scholar]
- Lobo, V.; Patil, A.; Phatak, A.; Chandra, N. Free radicals, antioxidants and functional foods: Impact on human health. Pharmacogn. Rev. 2010, 4, 118–126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rahal, A.; Kumar, A.; Singh, V.; Yadav, B.; Tiwari, R.; Chakraborty, S.; Dhama, K. Oxidative stress, prooxidants, and antioxidants: The interplay. BioMed Res. Int. 2014, 1, 1–19. [Google Scholar] [CrossRef] [Green Version]
- Nasri, H.; Rafieian-Kopaei, M. Oxidative stress and aging prevention. Int. J. Prev. Med. 2013, 4, 1101–1102. [Google Scholar]
- Patipong, T.; Hibino, T.; Waditee-Sirisattha, R.; Kageyama, H. Induction of antioxidative activity and antioxidant molecules in the halotolerant cyanobacterium Halothece sp. PCC7418 by temperature shift. Nat. Prod. Commun. 2019, 14, 1934578–19865680. [Google Scholar] [CrossRef] [Green Version]
- Smith, H.; Doyle, S.; Murphy, R. Filamentous fungi as a source of natural antioxidants. Food Chem. 2015, 185, 389–397. [Google Scholar] [CrossRef]
- Caicedo, N.H.; Davalos, A.F.; Puente, P.A.; Rodríguez, A.Y.; Caicedo, P.A. Antioxidant activity of exo-metabolites produced by Fusarium oxysporum: An endophytic fungus isolated from leaves of Otoba gracilipes. Microbiol. Open 2019, 8, 903. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Dong, M.; Chen, X.; Jiang, M.; Lv, X.; Yan, G. Antioxidant activity and phenolics of an endophytic Xylaria sp. from Ginkgo biloba. Food Chem. 2007, 105, 548–554. [Google Scholar] [CrossRef]
- Dulf, F.V.; Vodnar, D.C.; Dulf, E.H.; Pintea, A. Phenolic compounds, flavonoids, lipids and antioxidant potential of apricot (Prunus armeniaca L.) pomace fermented by two filamentous fungal strains in solid state system. Chem. Cen. J. 2017, 11, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Strobel, G.; Ford, E.; Worapong, J.; Harper, J.K.; Arif, A.M.; Grant, D.M.; Fung, P.C.W.; Ming Wah Chau, R. Isopestacin, an isobenzofuranone from Pestalotiopsis microspora, possessing antifungal and antioxidant activities. Phytochemistry 2002, 60, 179–183. [Google Scholar] [CrossRef]
- Li, P.; Luo, C.; Sun, W.; Lu, S.; Mou, Y.; Peng, Y.; Zhou, L. In-vitro antioxidant activities of polysaccharides from endophytic fungus Fusarium oxysporum Dzf17. Afr. J. Microbiol. Res. 2011, 5, 5994–5997. [Google Scholar]
- Srinivasan, K.; Jagadish, L.K.; Shenbhagaraman, R.; Muthumary, J. Antioxidant activity of endophytic fungus Phyllosticta sp. isolated from Guazuma tomentosa. J. Phytol. 2010, 1, 37–41. [Google Scholar]
- Huang, W.Y.; Cai, Y.Z.; Hyde, K.D.; Corke, H.; Sun, M. Endophytic fungi from Nerium oleander L (Apocynaceae): Main constituents and antioxidant activity. World J. Microbiol. Biotechnol. 2007, 23, 1253–1263. [Google Scholar] [CrossRef] [Green Version]
- Shen, L.; Ai, C.Z.; Song, Y.C.; Wang, F.W.; Jiao, R.H.; Zhang, A.H.; Man, H.Z.; Tan, R.X. Cytotoxic trichothecene macrolides produced by the endophytic Myrothecium roridum. J. Nat. Prod. 2019, 82, 1503–1509. [Google Scholar] [CrossRef]
- Torgerson, P.R.; Devleesschauwer, B.; Praet, N.; Speybroeck, N.; Willingham, A.L.; Kasuga, F.; Rokni, M.B.; Zhou, X.N.; Fèvre, E.M.; Sripa, B.; et al. World Health Organization estimates of the global and regional disease burden of 11 foodborne parasitic diseases, 2010: A data synthesis. PLoS Med. 2015, 12, e1001920. [Google Scholar] [CrossRef] [Green Version]
- Skinner-Adams, T.S.; Sumanadasa, S.D.; Fisher, G.M.; Davis, R.A.; Doolan, D.L.; Andrews, K.T. Defining the targets of antiparasitic compounds. Drug Discov. Today 2016, 21, 725–739. [Google Scholar] [CrossRef] [Green Version]
- Song, T.; Wang, C.; Tian, B. Modelling intra-host competition between malaria parasites strains. Comput. Appl. Math. 2020, 39, 48. [Google Scholar] [CrossRef]
- Vermelho, A.B.; Rodrigues, G.C.; Supuran, C.T. Why hasn’t there been more progress in new Chagas disease drug discovery? Expert Opin. Drug Discov. 2020, 15, 145–158. [Google Scholar] [CrossRef]
- Rottmann, M.; McNamara, C.; Yeung, B.K.; Lee, M.C.; Zou, B.; Russell, B.; Seitz, P.; Plouffe, D.M.; Dharia, N.V.; Tan, J.; et al. Spiroindolones, a potent compound class for the treatment of malaria. Science 2010, 329, 1175–1180. [Google Scholar] [CrossRef] [Green Version]
- Brissow, E.R.; da Silva, I.P.; de Siqueira, K.A.; Senabio, J.A.; Pimenta, L.P.; Januário, A.H.; Magalhães, L.G.; Furtado, R.A.; Tavares, D.C.; Junior, P.A.S.; et al. 18-Des-hydroxy Cytochalasin: An antiparasitic compound of Diaporthe phaseolorum-92C, an endophytic fungus isolated from Combretum lanceolatum Pohl ex Eichler. Parasit. Res. 2017, 116, 1823–1830. [Google Scholar] [CrossRef] [PubMed]
- Hayibor, K.; Kwain, S.; Osei, E.; Nartey, A.P.; Tetevi, G.M.; Owusu, K.B.A.; Camas, M.; Camas, A.S.; Kyeremeh, K. Ghanaian mangrove wetland endophytic fungus, Penicillium herquei strain BRS2A-AR produces (9Z, 11E)-13-oxooctadeca-9, 11-dienoic acid with activity against Trichomonas mobilensis. Int. J. Biol. Chem. Sci. 2019, 13, 1918–1937. [Google Scholar] [CrossRef] [Green Version]
- Shi, Y.N.; Pusch, S.; Shi, Y.M.; Richter, C.; Macia-Vicente, J.G.; Schwalbe, H.; Kaiser, M.; Opatz, T.; Bode, H.B. (±)-Alternarlactones A and B, Two Antiparasitic Alternariol-like Dimers from the Fungus Alternaria alternata P1210 isolated from the halophyte Salicornia sp. Org. Chem. 2019, 84, 11203–11209. [Google Scholar] [CrossRef] [PubMed]
- Mao, Z.; Wang, W.; Su, R.; Gu, G.; Liu, Z.L.; Lai, D.; Zhou, L. Hyalodendrins A and B, New decalin-type tetramic acid larvicides from the endophytic fungus Hyalodendriella sp. Ponipodef12. Molecules 2020, 25, 114. [Google Scholar] [CrossRef] [Green Version]
- Toghueo, R.M.K.; Kemgne, E.A.M.; Eke, P.; Kanko, M.I.M.; Dize, D.; Sahal, D.; Boyom, F.F. Antiplasmodial potential and GC-MS fingerprint of endophytic fungal extracts derived from Cameroonian Annona muricata. J. Ethnopharmacol. 2019, 235, 111–121. [Google Scholar] [CrossRef]
- Golias, H.C.; Polonio, J.C.; dos Santos Ribeiro, M.A.; Polli, A.D.; da Silva, A.A.; Bulla, A.M.; Volpato, H.; Nakamura, C.V.; Meurer, E.C.; Azevedo, J.L.; et al. Tibouchina granulosa (Vell.) Cogn (Melastomataceae) as source of endophytic fungi: Isolation, identification, and antiprotozoal activity of metabolites from Phyllosticta capitalensis. Braz. J. Microbiol. 2020, 51, 557–569. [Google Scholar] [CrossRef]
- Van Laar, J.M. Immunosuppressive Drugs. In Kelley and Firestein’s Textbook of Rheumatology, 10th ed.; Firestein, G.S., Budd, R.C., Gabriel, S.E., McInnes, I.B., O’Dell, J.R., Eds.; Elsevier: Amsterdam, The Netherlands, 2017; Chapter 62; pp. 983–998. [Google Scholar]
- Wang, W.X.; Cheng, G.G.; Li, Z.H.; Ai, H.L.; He, J.; Li, J.; Feng, T.; Liu, J.K. Curtachalasins, immunosuppressive agents from the endophytic fungus Xylaria cf. curta. Org. Biomol. Chem. 2019, 17, 7985–7994. [Google Scholar] [CrossRef]
- Liu, H.; Chen, S.; Liu, W.; Liu, Y.; Huang, X.; She, Z. Polyketides with immunosuppressive activities from mangrove endophytic fungus Penicillium sp. ZJ-SY2. Mar. Drugs 2016, 14, 217. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.W.; Wang, J.L.; Chen, J.; Chen, J.J.; Shen, J.W.; Feng, X.X.; Kubicek, C.P.; Lin, F.C.; Zhang, C.L.; Chen, F.Y. A novel derivative of (-) mycousnine produced by the endophytic fungus Mycosphaerella nawae, exhibits high and selective immunosuppressive activity on T cells. Front. Microbiol. 2017, 8, 1251. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.P.; Zhao, Z.Z.; Cheng, G.G.; Zhao, K.; Han, K.Y.; Zhou, L.; Feng, T.; Li, Z.H.; Liu, J.K. Immunosuppressive Nor-isopimarane Diterpenes from cultures of the Fungicolous Fungus Xylaria longipes HFG1018. J. Nat. Prod. 2020, 83, 401–412. [Google Scholar] [CrossRef]
- Andersen, P.I.; Ianevski, A.; Lysvand, H.; Vitkauskiene, A.; Oksenych, V.; Bjørås, M.; Telling, K.; Lutsar, I.; Dampis, U.; Irie, Y.; et al. Discovery and development of safe-in-man broad-spectrum antiviral agents. Int. J. Infec. Dis. 2020, 93, 268–276. [Google Scholar] [CrossRef] [PubMed]
- Linnakoski, R.; Reshamwala, D.; Veteli, P.; Cortina-Escribano, M.; Vanhanen, H.; Marjomäki, V. Antiviral agents from fungi: Diversity, mechanisms and potential applications. Front. Microbiol. 2018, 9, 2325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mason, S.; Devincenzo, J.P.; Toovey, S.; Wu, J.Z.; Whitley, R.J. Comparison of antiviral resistance across acute and chronic viral infections. Antivir. Res. 2018, 158, 103–112. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Sun, S.; Zhu, T.; Lin, Z.; Gu, J.; Li, D.; Gu, Q. Antiviral isoindolone derivatives from an endophytic fungus Emericella sp. associated with Aegiceras corniculatum. Phytochemistry 2011, 72, 1436–1442. [Google Scholar] [CrossRef]
- He, J.W.; Chen, G.D.; Gao, H.; Yang, F.; Li, X.X.; Peng, T.; Guo, L.D.; Yao, X.S. Heptaketides with antiviral activity from three endolichenic fungal strains Nigrospora sp., Alternaria sp. and Phialophora sp. Fitoterapia 2012, 83, 1087–1091. [Google Scholar] [CrossRef]
- Selim, K.A.; Elkhateeb, W.A.; Tawila, A.M.; El-Beih, A.A.; Abdel-Rahman, T.M.; El-Diwany, A.I.; Ahmed, E.F. Antiviral and antioxidant potential of fungal endophytes of Egyptian medicinal plants. Fermentation 2018, 4, 49. [Google Scholar] [CrossRef] [Green Version]
- Liu, S.S.; Jiang, J.X.; Huang, R.; Wang, Y.T.; Jiang, B.G.; Zheng, K.X.; Wu, S.H. A new antiviral 14-nordrimane sesquiterpenoid from an endophytic fungus Phoma sp. Phytochem. Lett. 2019, 29, 75–78. [Google Scholar] [CrossRef]
- Zhang, S.P.; Huang, R.; Li, F.F.; Wei, H.X.; Fang, X.W.; Xie, X.S.; Lin, D.G.; Wu, S.H.; He, J. Antiviral anthraquinones and azaphilones produced by an endophytic fungus Nigrospora sp. Aconitum carmichaeli. Fitoterapia 2016, 112, 85–89. [Google Scholar] [CrossRef]
- Raekiansyah, M.; Mori, M.; Nonaka, K.; Agoh, M.; Shiomi, K.; Matsumoto, A.; Morita, K. Identification of novel antiviral of fungus-derived brefeldin A against dengue viruses. Trop. Med. Health 2017, 45, 32. [Google Scholar] [CrossRef] [Green Version]
- Hawas, U.W.; Al-Farawati, R. Chemical constituents and antiviral activity from marine endophytic fungi from Red Sea alga Padina pavonica. J. Chem. Soc. Pak. 2017, 39, 478. [Google Scholar]
- Wellensiek, B.P.; Ramakrishnan, R.; Bashyal, B.P.; Eason, Y.; Gunatilaka, A.L.; Ahmad, N. Inhibition of HIV-1 replication by secondary metabolites from endophytic fungi of desert plants. Open Virol. J. 2013, 7, 72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Z.; Wu, K.; Xu, Y.; Xia, X.; Wang, X.; Ge, M.; Shao, L. Three novel chromanones with biological activities from the endophytic fungus Phomopsis CGMCC No. 5416. J. Antibiot. 2020, 73, 194–199. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. WHO Consolidated Guidelines on Tuberculosis: Tuberculosis Preventive Treatment: Module 1: Prevention: Tuberculosis Preventive Treatment. Available online: https://apps.who.int/iris/bitstream/handle/10665/331170/9789240001503-eng.pdf (accessed on 25 June 2020).
- Schön, T.; Miotto, P.; Köser, C.U.; Viveiros, M.; Böttger, E.; Cambau, E. Mycobacterium tuberculosis drug-resistance testing: Challenges, recent developments and perspectives. Clin. Microbiol. Infect. 2017, 23, 154–160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- World Health Organization (WHO). Fact sheets. Tuberculosis. Available online: https://www.who.int/news-room/fact-sheets/detail/tuberculosis (accessed on 27 June 2020).
- Uc-Cachón, A.H.; Gamboa-Angulo, M.; Borges-Argáez, R.; Reyes-Estebanez, M.; Said-Fernández, S.; Molina-Salinas, G.M. Antitubercular activity of the fungus Gliocladium sp. MR41 strain. Iran. J. Pharm. Res. IJPR 2019, 18, 860. [Google Scholar] [PubMed]
- Shah, A.; Rather, M.A.; Shah, A.M.; Mushtaq, S.; Hussain, A.; Rasool, S.; Parry, Z.A.; Qazi, P.H. Evaluating the in vitro antituberculosis, antibacterial and antioxidant potential of fungal endophytes isolated from Glycyrrhiza glabra L. Ann. Phytomed. 2016, 5, 140–146. [Google Scholar] [CrossRef]
- Mayer, A.; Guerrero, A.J.; Rodríguez, A.D.; Taglialatela-Scafati, O.; Nakamura, F.; Fusetani, N. Marine pharmacology in 2014–2015: Marine compounds with antibacterial, antidiabetic, antifungal, anti-inflammatory, antiprotozoal, antituberculosis, antiviral, and anthelmintic activities; affecting the immune and nervous systems, and other miscellaneous mechanisms of action. Mar. Drugs 2020, 18, 5. [Google Scholar]
- Ikram, M.; Ali, N.; Jan, G.; Hamayun, M.; Jan, F.G.; Iqbal, A. Novel antimicrobial and antioxidative activity by endophytic Penicillium roqueforti and Trichoderma reesei isolated from Solanum surattense. Acta Physiol. Plant. 2019, 41, 164. [Google Scholar] [CrossRef]
- Chaithra, M.; Vanitha, S.; Ramanathan, A.; Jegadeeshwari, V.; Rajesh, V.; Hegde, V.; Apshara, E. Profiling Secondary Metabolites of Cocoa (Theobroma cacao L.) Endophytic Fungi Lasiodiplodia pseudotheobromae PAK-7 and Lasiodiplodia theobromae TN-R-3 and their antimicrobial activities. Curr. J. Appl. Sci. Technol. 2020, 39, 47–56. [Google Scholar] [CrossRef] [Green Version]
- Wonglom, P.; Ito, S.I.; Sunpapao, A. Volatile organic compounds emitted from endophytic fungus Trichoderma asperellum T1 mediate antifungal activity, defense response and promote plant growth in lettuce (Lactuca sativa). Fungal Ecol. 2020, 43, 100867. [Google Scholar] [CrossRef]
- Yehia, R.S.; Osman, G.H.; Assaggaf, H.; Salem, R.; Mohamed, M.S. Isolation of potential antimicrobial metabolites from endophytic fungus Cladosporium cladosporioides from endemic plant Zygophyllum mandavillei. S. Afr. J. Bot. 2020, 129, 1–7. [Google Scholar] [CrossRef]
- Cao, X.; Shi, Y.; Wu, S.; Wu, X.; Wang, K.; Sun, H.; He, S.; Dickschat, J.S.; Wu, B. Polycyclic meroterpenoids, talaromyolides E−K for antiviral activity against pseudorabies virus from the endophytic fungus Talaromyces purpureogenus. Tetrahedron 2020, 76, 131349. [Google Scholar] [CrossRef]
- Tian, Y.Q.; Lin, X.P.; Wang, Z.; Zhou, X.F.; Qin, X.C.; Kaliyaperumal, K.; Zhang, T.Y.; Tu, Z.C.; Liu, Y. Asteltoxins with Antiviral Activities from the Marine Sponge-Derived Fungus Aspergillus sp. SCSIO XWS02F40. Molecules 2015, 21, 34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seetharaman, P.; Gnanasekar, S.; Chandrasekaran, R.; Chandrakasan, G.; Kadarkarai, M.; Sivaperumal, S. Isolation and characterization of anticancer flavone chrysin (5, 7-dihydroxy flavone)-producing endophytic fungi from Passiflora incarnata L. leaves. Ann. Microbiol. 2017, 67, 321–331. [Google Scholar] [CrossRef]
- Da Rosa, B.V.; Kuhn, K.R.; Ugalde, G.A.; Zabot, G.L.; Kuhn, R.C. Antioxidant compounds extracted from Diaporthe schini using supercritical CO 2 plus cosolvent. Bioprocess Biosyst. Eng. 2020, 43, 133–141. [Google Scholar] [CrossRef] [PubMed]
- Druzian, S.P.; Pinheiro, L.N.; Susin, N.M.B.; Dal Prá, V.; Mazutti, M.A.; Kuhn, R.C.; de Marsillac Terra, L. Production of metabolites with antioxidant activity by Botryosphaeria dothidea in submerged fermentation. Bioprocess Biosyst. Eng. 2020, 43, 13–20. [Google Scholar] [CrossRef]
- Ran, X.; Zhang, G.; Li, S.; Wang, J. Characterization and antitumor activity of camptothecin from endophytic fungus Fusarium solani isolated from Camptotheca acuminate. Afr. Health Sci. 2017, 566–574. [Google Scholar] [CrossRef] [Green Version]
- Shah, A.; Rather, M.A.; Hassan, Q.P.; Aga, M.A.; Mushtaq, S.; Shah, A.M.; Hussain, A.; Baba, S.A.; Ahmad, Z. Discovery of anti-microbial and anti-tubercular molecules from Fusarium solani: An endophyte of Glycyrrhiza glabra. J. Appl. Microbiol. 2017, 122, 1168–1176. [Google Scholar] [CrossRef]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| OXIDATIVE STRESS | |
|---|---|
| Neurological | Multi-System Effects |
| Attention-deficit/hyperactivity disorder (ADHD) Alzheimer’s disease Anxiety and depression Asperger syndrome Autism Multiple sclerosis Parkinson’s disease | Diabetes Cancer Inflammation Fibromyalgia Lyme disease Chronic fatigue syndrome Metabolic syndrome Anxiety Hyperthyroidism Sleep apnea |
| Cardiovascular | Joints/Skin |
| Cardiovascular Disease Angina Pectoris Hypertension Atherosclerosis | Gout Dermatitis Rheumatoid arthritis Carpal tunnel syndrome |
| Gastrointestinal Disorder | Respiratory |
| Crohn’s Disease Gastroesophageal reflux disease (GERD) Gastric ulcers Celiac disease Functional dyspepsia | Chronic obstructive pulmonary disease (COPD) Asthma |
| Endophytic Fungi | Host Plant | Bioactive Compounds | Biological Properties | Activity Level | Ref. |
|---|---|---|---|---|---|
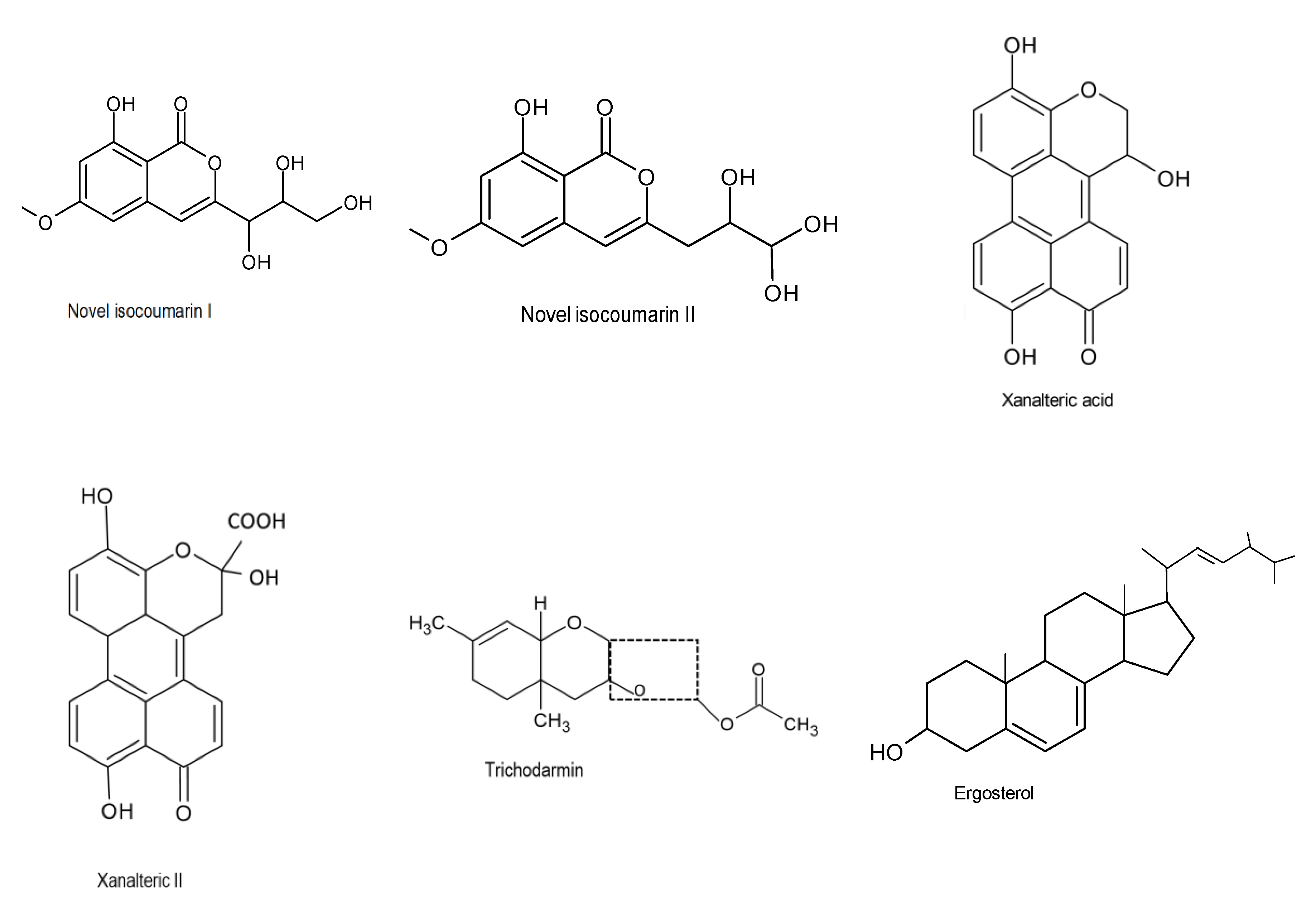
| Penicillium funiculosum Fes1711 and Trichoderma harzianum Fes1712 | Ficus elastica | Isocoumarin derivatives | Antibacterial activity | MIC = 32 μg/mL | [34] |
| Phomopsis sp. CFS42 | Cephalotaxus fortunei | Polyketides | Antifungal activity | MIC = 2.5 μg/mL | [44] |
| Chaetomium globosum | Ginkgo biloba | Azaphilone alkaloids | Anticancer activity | IC50 = 53.4 μM | [58] |
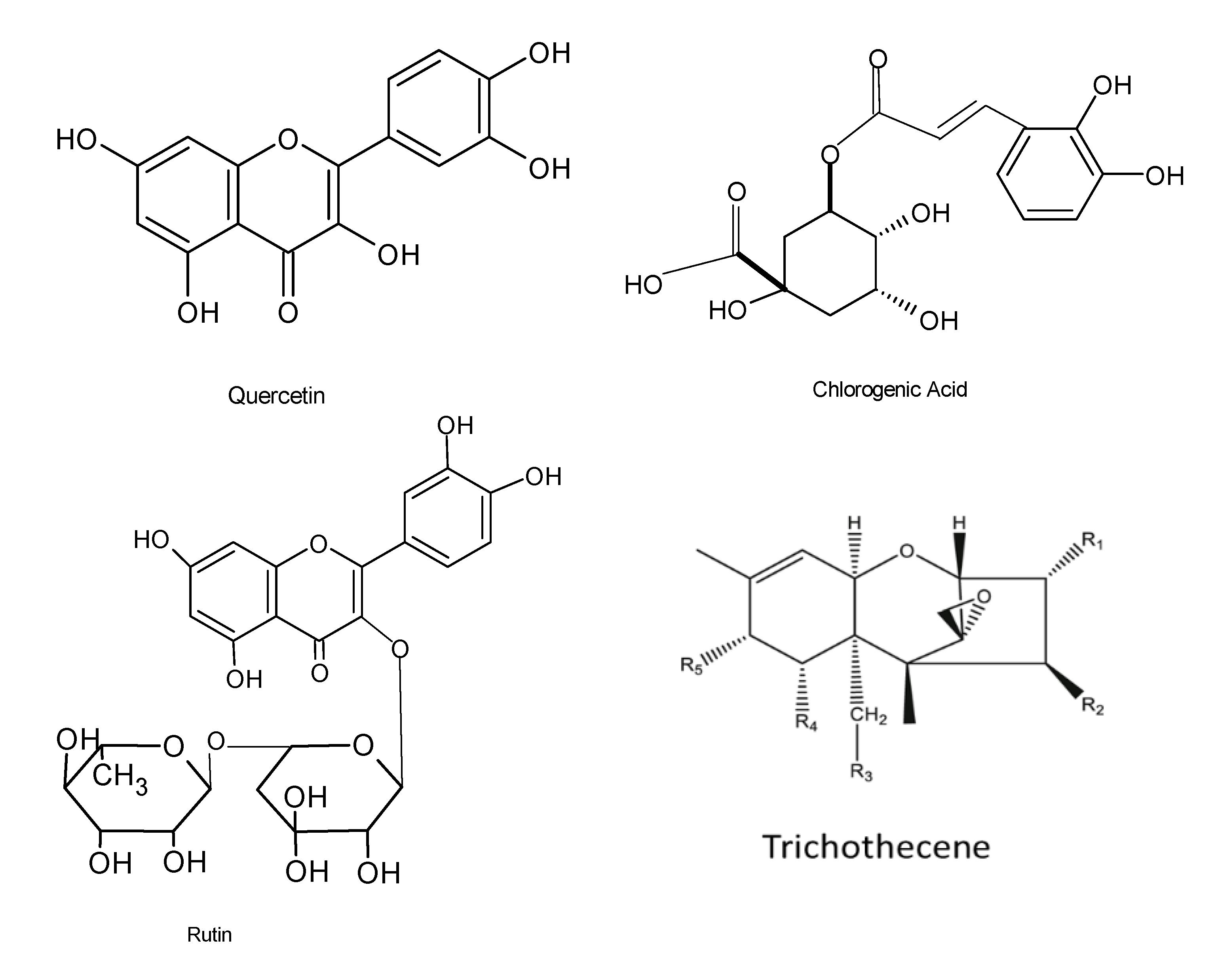
| Alternaria alternata AE1 | Azadirachta indica | Phenolics and flavonoids | Antioxidant properties | IC50 = 38 μg/mL | [28] |
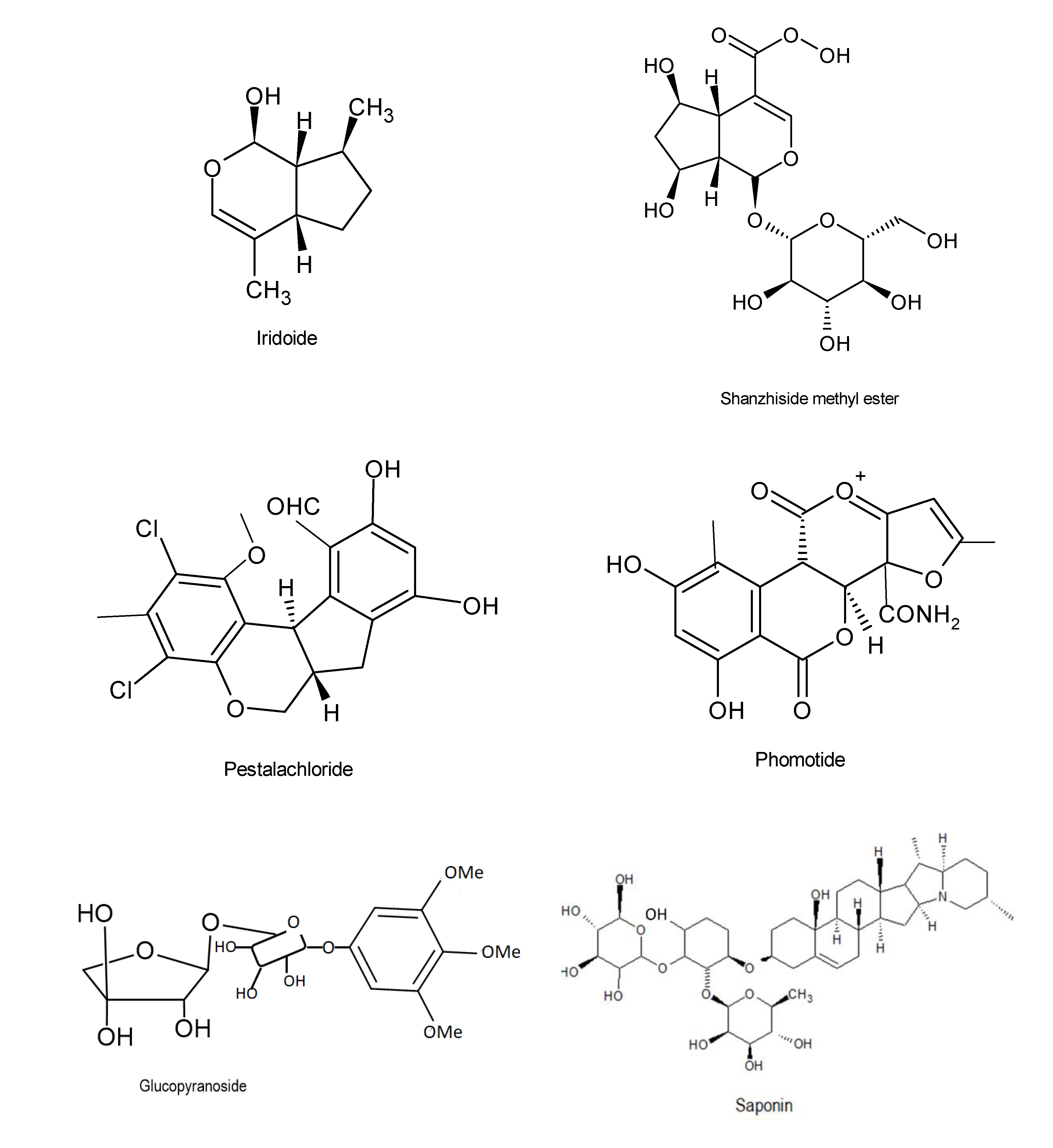
| Mycosphaerella nawae ZJLQ129 | Smilax china | Amide derivative | Immunosuppressant activity | 30 and 300 nM | [98] |
| Phomopsis sp. CGMCC No. 5416 | Achyranthes bidentata | Chromanones | Antiviral activity | IC50 =32.5 μg/ ml | [111] |
| Gliocladium sp. MR41 | Culture collection | Polyols | Antitubercular properties | MIC = 3.13 µg/mL | [116] |
| Penicillium roqueforti and Trichoderma reesei | Solanum surattense | Ferulic acid, cinnamic acid, quercetin, and rutin | Antibacterial activity | MBC = 2.5 µg/mL | [118] |
| Lasiodiplodia pseudotheobromae PAK-7 and L. theobromae TN-R-3 | Theobroma cacao L. | dl-Mevalonic acid lactone, Methyl 6-O-[1-methylpropyl]-á-d-galactopyranoside | Antibacterial activity | MIC= 21 mm | [119] |
| Trichoderma asperellum T1 | Culture collection | 6-pentyl-2H-pyran-2-one (6-PP) | Antifungal and plant promoting properties | 61.31% Inhibition | [120] |
| Cladosporium cladosporioides | Zygophyllum mandavillei | 3-phenylpropionic acid, 5′-hydroxyasperentin | Antifungal activity | MIC = 15.62 μg/mL | [121] |
| Talaromyces purpureogenus | Grateloupia filicina | Talaromyolide K | Antiviral activity | 60.11% Inhibition | [122] |
| Aspergillus sp. SCSIO XWS02F40 | Callyspongia sp. | Asteltoxins | Antiviral activity | IC50 = 3.5 μg/mL | [123] |
| Diaporthe schini | Solanum americanum | 1,4-diaza-2,5-dioxo-3-isobutyl bicyclo[4.3.0]nonane and benzeneethanol | Antioxidant activity | DPPH radical = 96.62% | [124] |
| Botryosphaeria dothidea | Pampa and Atlantic Forest Plants | Hexahydropyrrolizin-3-one and (2-methylpropyl) ester | Antioxidant activity | IC50 = 0.206 mg/mL | [125] |
| Fusarium solani S-019 | Camptotheca acuminate | Camptothecin | Anticancer activity | 50 µg/mL | [126] |
| Alternaria alternata KT380662 | Passiflora incarnata L. | Flavone chrysin (5,7-dihydroxy flavone) | Anticancer activity | IC50 = 37.97 μg/mL | [127] |
| Diaporthe phaseolorum 92C | Combretum lanceolatum | 18-Des-hydroxy Cytochalasin | Antiparasitic activity | IC50 = 50 μg/mL | [124] |
| Phyllosticta capitalensi | Tibouchina granulosa | Brefeldin and heptelidic acid | Antiparasitic activity | IC50 = 50.13 μg/mL, | [89] |
| Fusarium solani | Glycyrrhiza glabra | Fusarubin, 3-O-methylfusarubin, and javanicin | Antitubercular activity | MIC = 8 μg/mL | [128] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Manganyi, M.C.; Ateba, C.N. Untapped Potentials of Endophytic Fungi: A Review of Novel Bioactive Compounds with Biological Applications. Microorganisms 2020, 8, 1934. https://doi.org/10.3390/microorganisms8121934
Manganyi MC, Ateba CN. Untapped Potentials of Endophytic Fungi: A Review of Novel Bioactive Compounds with Biological Applications. Microorganisms. 2020; 8(12):1934. https://doi.org/10.3390/microorganisms8121934
Chicago/Turabian StyleManganyi, Madira Coutlyne, and Collins Njie Ateba. 2020. "Untapped Potentials of Endophytic Fungi: A Review of Novel Bioactive Compounds with Biological Applications" Microorganisms 8, no. 12: 1934. https://doi.org/10.3390/microorganisms8121934
APA StyleManganyi, M. C., & Ateba, C. N. (2020). Untapped Potentials of Endophytic Fungi: A Review of Novel Bioactive Compounds with Biological Applications. Microorganisms, 8(12), 1934. https://doi.org/10.3390/microorganisms8121934