Analysis of the Bacterial Diversity of Paipa Cheese (a Traditional Raw Cow’s Milk Cheese from Colombia) by High-Throughput Sequencing

 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cheese Sampling
2.2. DNA Extraction
2.3. DNA Sequencing and Analysis
3. Results
3.1. Characteristics of Sample Sequence Reads
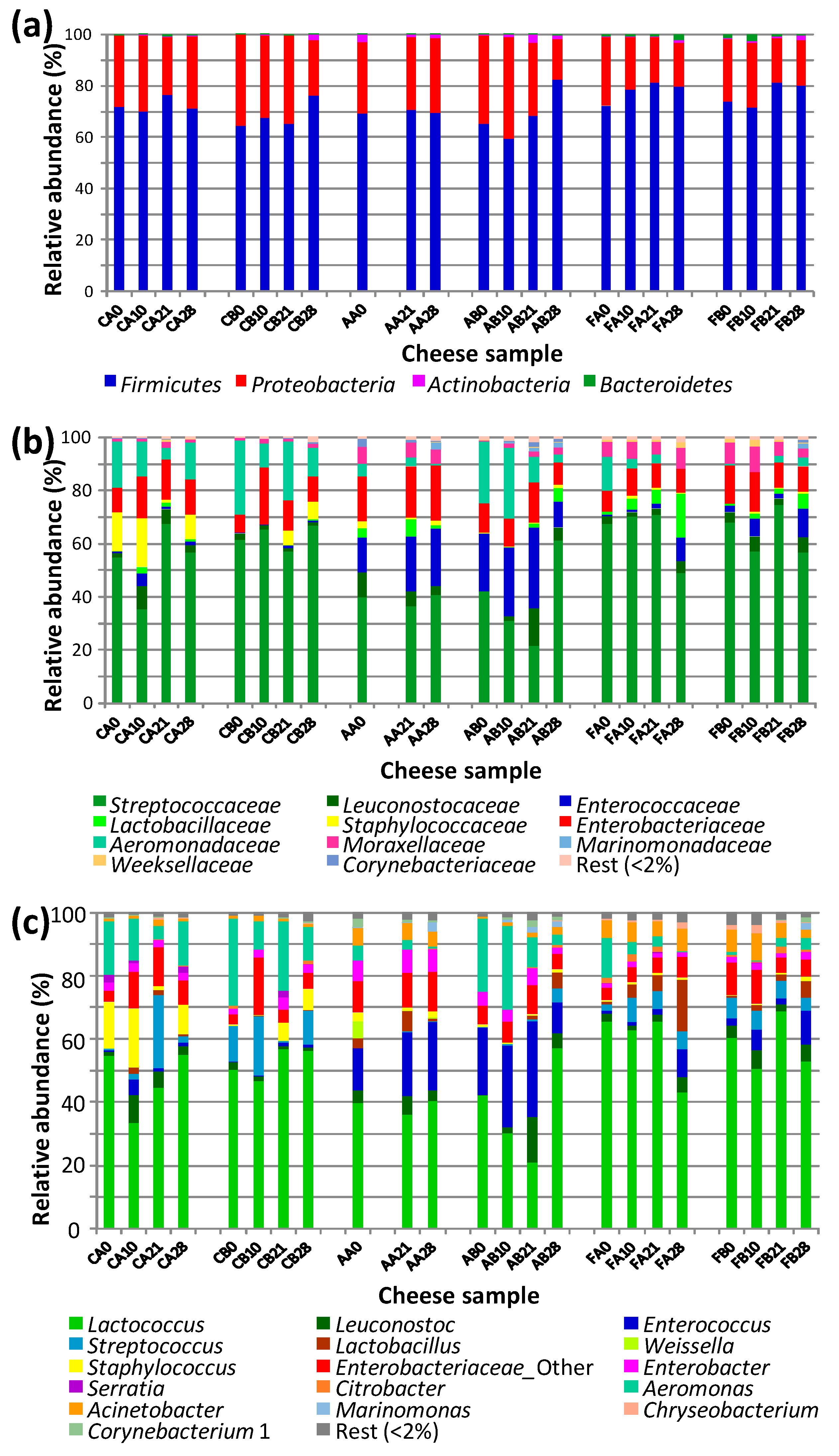
3.2. Bacterial Populations in Cheese Samples
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- O’Sullivan, D.J.; Cotter, P.D.; O’Sullivan, O.; Giblin, L.; McSweeney, P.L.; Sheehan, J.J. Temporal and spatial differences in microbial composition during the manufacture of a continental-type cheese. Appl. Environ. Microbiol. 2015, 81, 2525–2533. [Google Scholar] [CrossRef] [Green Version]
- Quigley, L.; O’Sullivan, O.; Beresford, T.P.; Ross, R.P.; Fitzgerald, G.F.; Cotter, P.D. High-throughput sequencing for detection of subpopulations of bacteria not previously associated with artisanal cheeses. Appl. Environ. Microbiol. 2012, 78, 5717–5723. [Google Scholar] [CrossRef] [Green Version]
- Yeluri Jonnala, B.R.; McSweeney, P.L.H.; Sheehan, J.J.; Cotter, P.D. Sequencing of the Cheese Microbiome and Its Relevance to Industry. Front. Microbiol. 2018, 9, 1020. [Google Scholar] [CrossRef] [Green Version]
- Frétin, M.; Martin, B.; Rifa, E.; Isabelle, V.M.; Pomiès, D.; Ferlay, A.; Montel, M.C.; Delbès, C. Bacterial community assembly from cow teat skin to ripened cheeses is influenced by grazing systems. Sci. Rep. 2018, 8, 200. [Google Scholar] [CrossRef] [Green Version]
- Quigley, L.; McCarthy, R.; O’Sullivan, O.; Beresford, T.P.; Fitzgerald, G.F.; Ross, R.P.; Stanton, C.; Cotter, P.D. The microbial content of raw and pasteurized cow milk as determined by molecular approaches. J. Dairy Sci. 2013, 96, 4928–4937. [Google Scholar] [CrossRef]
- Kable, M.E.; Srisengfa, Y.; Laird, M.; Zaragoza, J.; McLeod, J.; Heidenreich, J.; Marco, M.L. The Core and Seasonal Microbiota of Raw Bovine Milk in Tanker Trucks and the Impact of Transfer to a Milk Processing Facility. MBio 2016, 7, e00836-16. [Google Scholar] [CrossRef] [Green Version]
- Stellato, G.; De Filippis, F.; La Storia, A.; Ercolini, D. Coexistence of lactic acid bacteria and potential spoilage microbiota in a dairy processing environment. Appl. Environ. Microbiol. 2015, 81, 7893–7904. [Google Scholar] [CrossRef] [Green Version]
- Falardeau, J.; Keeney, K.; Trmčić, A.; Kitts, D.; Wang, S. Farm-to-fork profiling of bacterial communities associated with an artisan cheese production facility. Food Microbiol. 2019, 83, 48–58. [Google Scholar] [CrossRef]
- Giello, M.; La Storia, A.; Masucci, F.; Di Francia, A.; Ercolini, D.; Villani, F. Dynamics of bacterial communities during manufacture and ripening of traditional Caciocavallo of Castelfranco cheese in relation to cows’ feeding. Food Microbiol. 2017, 63, 170–177. [Google Scholar] [CrossRef]
- Bokulich, N.A.; Mills, D.A. Facility-specific “house” microbiome drives microbial landscapes of artisan cheesemaking plants. Appl. Environ. Microbiol. 2013, 79, 5214–5223. [Google Scholar] [CrossRef] [Green Version]
- Aldrete-Tapia, A.; Escobar-Ramírez, M.C.; Tamplin, M.L.; Hernández-Iturriaga, M. High-throughput sequencing of microbial communities in Poro cheese, an artisanal Mexican cheese. Food Microbiol. 2014, 44, 136–141. [Google Scholar] [CrossRef]
- Porcellato, D.; Skeie, S.B. Bacterial dynamics and functional analysis of microbial metagenomes during ripening of Dutch-type cheese. Int. Dairy J. 2016, 61, 182–188. [Google Scholar] [CrossRef]
- Ceugniez, A.; Taminiau, B.; Coucheney, F.; Jacques, P.; Delcenserie, V.; Daube, G.; Drider, D. Use of a metagenetic approach to monitor the bacterial microbiota of “Tomme d’Orchies” cheese during the ripening process. Int. J. Food Microbiol. 2017, 247, 65–69. [Google Scholar] [CrossRef]
- Riquelme, C.; Câmara, S.; Dapkevicius Mde, L.; Vinuesa, P.; da Silva, C.C.; Malcata, F.X.; Rego, O.A. Characterization of the bacterial biodiversity in Pico cheese (an artisanal Azorean food). Int. J. Food Microbiol. 2015, 192, 86–94. [Google Scholar] [CrossRef] [Green Version]
- Silvetti, T.; Capra, E.; Morandi, S.; Cremonesi, P.; Decimo, M.; Gavazzi, F.; Giannico, R.; De Noni, I.; Brasca, M. Microbial population profile during ripening of Protected Designation of Origin (PDO) Silter cheese, produced with and without autochthonous starter culture. LWT-Food Sci. Technol. 2017, 84, 821–831. [Google Scholar] [CrossRef]
- Dugat-Bony, E.; Garnier, L.; Denonfoux, J.; Ferreira, S.; Sarthou, A.S.; Bonnarme, P.; Irlinger, F. Highlighting the microbial diversity of 12 French cheese varieties. Int. J. Food Microbiol. 2016, 238, 265–273. [Google Scholar] [CrossRef]
- Delcenserie, V.; Taminiau, B.; Delhalle, L.; Nezer, C.; Doyen, P.; Crevecoeur, S.; Roussey, D.; Korsak, N.; Daube, G. Microbiota characterization of a Belgian protected designation of origin cheese, Herve cheese, using metagenomic analysis. J. Dairy Sci. 2014, 97, 6046–6056. [Google Scholar] [CrossRef] [Green Version]
- Xue, J.; Yang, Y.; Wang, Z.; Guo, Y.; Shao, Y. Bacterial Diversity in Chinese Rushan Cheese from Different Geographical Origins. Front. Microbiol. 2018, 9, 1920. [Google Scholar] [CrossRef]
- Kamimura, B.A.; De Filippis, F.; Sant’Ana, A.S.; Ercolini, D. Large-scale mapping of microbial diversity in artisanal Brazilian cheeses. Food Microbiol. 2019, 80, 40–49. [Google Scholar] [CrossRef]
- O’Sullivan, D.J.; Giblin, L.; McSweeney, P.L.H.; Sheehan, J.J.; Cotter, P.D. Nucleic acid-based approaches to investigate microbial-related cheese quality defects. Front. Microbiol. 2013, 4, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Dalmasso, A.; de los Dolores Soto del Rio, M.; Civera, T.; Pattono, D.; Cardazzo, B.; Bottero, M.T. Characterization of microbiota in Plaisentif cheese by high-throughput sequencing. LWT Food Sci. Technol. 2016, 69, 490–496. [Google Scholar] [CrossRef]
- Robayo, A.M.; Pachón, F.A. Characterization of Paipa and campesino cheeses chain in the framework of programa mercados campesinos: Two cases study. Rev. Med. Vet. Zootec. 2013, 63, 196–212. [Google Scholar]
- Superintendencia de Industria y Comercio de Colombia. Por la Cual se Concede la Protección con Denominación de Origen al Queso PAIPA; Resolución 0070-802; Superintendencia de Industria y Comercio de Colombia: Bogotá, Colombia, 6 December 2011. Available online: https://www.sic.gov.co/sites/default/files/files/Denominacion%20de%20Origen/Agro%20-%20Alimenticios/Queso%20Paipa/queso_paipa.pdf (accessed on 24 December 2019).
- Klindworth, A.; Pruesse, E.; Schweer, T.; Peplies, J.; Quast, C.; Horn, M.; Glöckner, F.O. Evaluation of general 16S ribosomal RNA gene PCR primers for classical and next-generation sequencing-based diversity studies. Nucleic Acids Res. 2013, 41, e1. [Google Scholar] [CrossRef]
- Schmieder, R.; Edwards, R. Quality control and preprocessing of metagenomic datasets. Bioinformatics 2011, 27, 863–864. [Google Scholar] [CrossRef] [Green Version]
- Caporaso, J.G.; Lauber, C.L.; Walters, W.A.; Berg-Lyons, D.; Lozupone, C.A.; Turnbaugh, P.J.; Fierer, N. Global patterns of 16S rRNA diversity at a depth of millions of sequences per sample. Proc. Natl. Acad. Sci. USA 2011, 108, 4516–4522. [Google Scholar] [CrossRef] [Green Version]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [Green Version]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2013, 41, 590–596. [Google Scholar] [CrossRef]
- Quigley, L.; O’Sullivan, O.; Beresford, T.P.; Ross, P.R.; Fitzgerald, G.F.; Cotter, P.D. A comparison of methods used to extract bacterial DNA from raw milk and raw milk cheese. J. Appl. Microbiol. 2012, 113, 96–105. [Google Scholar] [CrossRef]
- Lima, S.F.; de Souza Bicalho, M.L.; Bicalho, R.C. Evaluation of milk sample fractions for characterization of milk microbiota from healthy and clinical mastitis cows. PLoS ONE 2018, 13, e0193671. [Google Scholar] [CrossRef] [Green Version]
- Carafa, I.; Stocco, G.; Nardin, T.; Larcher, R.; Bittante, G.; Tuohy, K.; Franciosi, E. Production of Naturally γ–Aminobutyric Acid-Enriched Cheese Using the Dairy Strains Streptococcus thermophilus 84C and Lactobacillus brevis DSM 32386. Front. Microbiol. 2019, 10, 93. [Google Scholar] [CrossRef]
- Giraffa, G. Functionality of enterococci in dairy products. Int. J. Food Microbiol. 2003, 88, 215–222. [Google Scholar] [CrossRef]
- Foulquié-Moreno, M.R.; Sarantinopoulos, P.; Tsakalidou, E.; De Vuyst, L. The role and application of enterococci in food and health. Int. J. Food Microbiol. 2006, 106, 1–24. [Google Scholar] [CrossRef]
- Vandera, E.; Kakouri, A.; Koukkou, A.I.; Samelis, J. Major ecological shifts within the dominant nonstarter lactic acid bacteria in mature Greek Graviera cheese as affected by the starter culture type. Int. J. Food Microbiol. 2019, 290, 15–26. [Google Scholar] [CrossRef]
- Franz, C.M.A.P.; Van Belkum, M.J.; Holzapfel, W.H.; Abriouel, H.; Gálvez, A. Diversity of enterococcal bacteriocins and their grouping in a new classification scheme. FEMS Microbiol. Rev. 2007, 31, 293–310. [Google Scholar] [CrossRef]
- Hanchi, H.; Mottawea, W.; Sebei, K.; Hammami, R. The genus Enterococcus: Between probiotic potential and safety concerns—An update. Front. Microbiol. 2018, 9, 1791. [Google Scholar] [CrossRef]
- Barbieri, F.; Montanari, C.; Gardini, F.; Tabanelli, G. Biogenic amine production by lactic acid bacteria: A review. Foods 2019, 8, 17. [Google Scholar] [CrossRef] [Green Version]
- García-Solache, M.; Rice, L.B. The Enterococcus: A model of adaptability to its environment. Clin. Microbiol. Rev. 2019, 32, e00058-18. [Google Scholar] [CrossRef] [Green Version]
- Jamet, E.; Akary, E.; Poisson, M.A.; Chamba, J.F.; Bertrand, X.; Serror, P. Prevalence and characterization of antibiotic resistant Enterococcus faecalis in French cheeses. Food Microbiol. 2012, 31, 191–198. [Google Scholar] [CrossRef]
- Gaglio, R.; Couto, N.; Marques, C.; de Fatima Silva Lopes, M.; Moschetti, G.; Pomba, C.; Settanni, L. Evaluation of antimicrobial resistance and virulence of enterococci from equipment surfaces, raw materials, and traditional cheeses. Int. J. Food Microbiol. 2016, 236, 107–114. [Google Scholar] [CrossRef] [Green Version]
- Silvetti, T.; Morandi, S.; Brasca, M. Does Enterococcus faecalis from traditional raw milk cheeses serve as a reservoir of antibiotic resistance and pathogenic traits? Foodborne Pathog. Dis. 2019, 16, 359–367. [Google Scholar] [CrossRef]
- Kummel, J.; Stessl, B.; Gonano, M.; Walcher, G.; Bereuter, O.; Fricker, M.; Grunert, T.; Wagner, M.; Ehling-Schulz, M. Staphylococcus aureus Entrance into the Dairy Chain: Tracking, S. aureus from Dairy Cow to Cheese. Front. Microbiol. 2016, 7, 1603. [Google Scholar] [CrossRef] [Green Version]
- Bellio, A.; Chiesa, F.; Gallina, S.; Bianchi, D.M.; Macori, G.; Bossi, D.; Nia, Y.; Mutel, I.; Messio, S.; Hennekinne, J.A.; et al. Insight Into the Distribution of Staphylococci and Their Enterotoxins in Cheeses Under Natural Conditions. Front. Microbiol. 2019, 9, 3233. [Google Scholar] [CrossRef]
- Herrera, F.C.; García-López, M.L.; Santos, J.A. Short communication: Characterization of methicillin-resistant Staphylococcus aureus isolated from raw milk fresh cheese in Colombia. J. Dairy Sci. 2016, 99, 7872–7876. [Google Scholar] [CrossRef] [Green Version]
- Adame-Gómez, R.; Toribio-Jimenez, J.; Vences-Velazquez, A.; Rodríguez-Bataz, E.; Santiago Dionisio, M.C.; Ramirez-Peralta, A. Methicillin-Resistant Staphylococcus aureus (MRSA) in Artisanal Cheeses in México. Int. J. Microbiol. 2018, 18, 8760357. [Google Scholar] [CrossRef]
- Gonzalez, A.G.M.; Marques, L.M.P.; Gomes, M.D.S.A.; Beltrão, J.C.D.C.; Pinheiro, M.G.; Esper, L.M.R.; Paula, G.R.; Teixeira, L.A.; Aguiar-Alves, F. Methicillin-resistant Staphylococcus aureus in minas frescal cheese: Evaluation of classic enterotoxin genes, antimicrobial resistance and clonal diversity. FEMS Microbiol. Lett. 2017, 364, fnx232. [Google Scholar] [CrossRef] [Green Version]
- Ahmed, A.A.-H.; Maharik, N.M.S.; Valero, A.; Kamal, S.M. Incidence of enterotoxigenic Staphylococcus aureus in milk and Egyptian artisanal dairy products. Food Control 2019, 104, 20–27. [Google Scholar] [CrossRef]
- Alnakip, M.E.; Quintela-Baluja, M.; Böhme, K.; Caamaño-Antelo, S.; Barros-Velázquez, J. Molecular characterisation and typing the methicillin resistance of Staphylococcus spp. isolated from raw milk and cheeses in Northwest Spain: A mini survey. Int. Dairy J. 2019, 89, 68–76. [Google Scholar] [CrossRef]
- Chajęcka-Wierzchowska, W.; Zadernowska, A.; Gajewska, J. S. epidermidis strains from artisanal cheese made from unpasteurized milk in Poland—Genetic characterization of antimicrobial resistance and virulence determinants. Int. J. Food Microbiol. 2019, 294, 55–59. [Google Scholar] [CrossRef]
- Hammer, P.; Jordan, J.; Jacobs, C.; Klempt, M. Characterization of coagulase-negative staphylococci from brining baths in Germany. J. Dairy Sci. 2019, 102, 8734–8744. [Google Scholar] [CrossRef]
- Bachmann, H.P.; Spahr, U. The fate of potentially pathogenic bacteria in Swiss hard and semihard cheeses made from raw milk. J. Dairy Sci. 1995, 78, 476–483. [Google Scholar] [CrossRef]
- Ahmed, N.I.; Abd El Aal, S.F.A.; Ayoub, M.A.; El Sayed, M.S. Enumeration and Characterization of Aeromonas spp. Isolated from Milk and Some Dairy Products in Sharkia Governorate Egypt. Alexandria. J. Vet. Sci. 2014, 40, 52–64. [Google Scholar]
- Melas, D.S.; Papageorgiou, D.K.; Mantis, A.I. Enumeration and confirmation of Aeromonas hydrophila, Aeromonas caviae, and Aeromonas sobria isolated from raw milk and other milk products in Northern Greece. J. Food Prot. 1999, 62, 463–466. [Google Scholar] [CrossRef] [PubMed]
- Uraz, G.; Coskun, S.; Ozer, B. Microflora and pathogen bacteria (Salmonella, Klebsiella, Yersinia, Pseudomonas, Aeromonas, Escherichia coli, Staphylococcus aureus) in Urfa cheese (a traditional white-brined Turkish cheese). Pak. J. Nutr. 2008, 7, 630–635. [Google Scholar] [CrossRef]
- Santos, J.A.; López-Díaz, T.M.; García-Fernández, M.C.; García-López, M.L.; Otero, A. Villalón, a fresh ewe’s milk Spanish cheese, as a source of potentially pathogenic Aeromonas strains. J. Food Prot. 1996, 59, 1288–1291. [Google Scholar] [CrossRef]
- Freitas, A.C.; Nunes, M.P.; Milhomem, A.M.; Ricciardi, I.D. Occurrence and characterization of Aeromonas species in pasteurized milk and white cheese in Rio de Janeiro, Brasil. J. Food Prot. 1993, 56, 62–65. [Google Scholar] [CrossRef]
- Araújo, V.S.; Pagliares, V.A.; Queiroz, M.L.P.; Freitas-Almeida, A.C. Occurrence of Staphylococcus and enteropathogens in soft cheese commercialized in the city of Rio de Janeiro, Brazil. J. Appl. Microbiol. 2002, 92, 1172–1177. [Google Scholar] [CrossRef] [Green Version]
- Cereser, N.D.; Rossi Júnior, O.D.; Martineli, T.M.; De Souza, V.; Rodrigues, L.B.; Cardozo, M.V. Aeromonas in processing line of minas frescal and colonial cheeses. ARS Vet. 2013, 29, 23–29. [Google Scholar] [CrossRef] [Green Version]
- Janda, J.M.; Abbott, S.L. The genus Aeromonas: Taxonomy, pathogenicity, and infection. Clin. Microbiol. Rev. 2010, 23, 35–73. [Google Scholar] [CrossRef] [Green Version]
- Schuetz, A.N. Emerging agents of gastroenteritis: Aeromonas, Plesiomonas, and the diarrheagenic pathotypes of Escherichia coli. Semin. Diagnoatic Pathol. 2019, 36, 187–192. [Google Scholar] [CrossRef]
- Cousin, M.A. Presence and activity of psychrotrophic microorganisms in milk and dairy-products—A review. J. Food Prot. 1982, 45, 172–207. [Google Scholar] [CrossRef]
- Addis, E.; Fleet, G.H.; Cox, J.M.; Kolak, D.; Leung, T. The growth, properties and interactions of yeasts and bacteria associated with the maturation of Camembert and blue-veined cheeses. Int. J. Food Microbiol. 2001, 69, 25–36. [Google Scholar] [CrossRef]
- Baruzzi, F.; Lagonigro, R.; Quintieri, L.; Morea, M.; Caputo, L. Occurrence of non-lactic acid bacteria populations involved in protein hydrolysis of cold-stored high moisture Mozzarella cheese. Food Microbiol. 2012, 30, 37–44. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Sample | N°Reads | N°Observations | Chao1 | ACE | Shannon | Simpson |
|---|---|---|---|---|---|---|
| CA0 | 119123.00 | 32.00 | 32.00 | 32.00 | 1.50 | 0.65 |
| CA10 | 117789.00 | 28.00 | 28.00 | 28.00 | 1.98 | 0.81 |
| CA21 | 100681.00 | 33.00 | 33.00 | 33.00 | 1.73 | 0.73 |
| CA28 | 106138.00 | 32.00 | 32.00 | 32.00 | 1.64 | 0.66 |
| CB0 | 101205.00 | 25.00 | 25.00 | 25.00 | 1.41 | 0.66 |
| CB10 | 105216.00 | 27.00 | 27.00 | 27.00 | 1.55 | 0.71 |
| CB21 | 112479.00 | 32.00 | 32.00 | 32.00 | 1.45 | 0.62 |
| CB28 | 104831.00 | 31.00 | 31.00 | 31.00 | 1.65 | 0.66 |
| AA0 | 124879.00 | 33.00 | 33.00 | 34.08 | 2.10 | 0.80 |
| AA10 | 6744.00 | 13.00 | 13.00 | 13.00 | 1.71 | 0.67 |
| AA21 | 130445.00 | 30.00 | 30.00 | 30.00 | 1.99 | 0.80 |
| AA28 | 125251.00 | 35.00 | 35.00 | 35.00 | 1.89 | 0.77 |
| AB0 | 189360.00 | 26.00 | 26.00 | 26.00 | 1.52 | 0.72 |
| AB10 | 153619.00 | 31.00 | 31.00 | 31.35 | 1.74 | 0.77 |
| AB21 | 158773.00 | 34.00 | 34.00 | 34.44 | 2.05 | 0.82 |
| AB28 | 157004.00 | 41.00 | 41.00 | 41.00 | 1.74 | 0.66 |
| FA0 | 161174.00 | 41.00 | 41.00 | 41.00 | 1.40 | 0.55 |
| FA10 | 128947.00 | 37.00 | 37.00 | 37.00 | 1.57 | 0.59 |
| FA21 | 151273.00 | 39.00 | 39.00 | 39.00 | 1.50 | 0.56 |
| FA28 | 151121.00 | 45.00 | 45.00 | 45.00 | 1.91 | 0.76 |
| FB0 | 152092.00 | 44.00 | 44.00 | 44.00 | 1.59 | 0.61 |
| FB10 | 162325.00 | 51.00 | 51.00 | 51.00 | 1.86 | 0.71 |
| FB21 | 177231.00 | 45.00 | 45.00 | 45.00 | 1.41 | 0.52 |
| FB28 | 160890.00 | 44.00 | 44.00 | 44.00 | 1.86 | 0.70 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Castellanos-Rozo, J.; Pérez Pulido, R.; Grande, M.J.; Lucas, R.; Gálvez, A. Analysis of the Bacterial Diversity of Paipa Cheese (a Traditional Raw Cow’s Milk Cheese from Colombia) by High-Throughput Sequencing. Microorganisms 2020, 8, 218. https://doi.org/10.3390/microorganisms8020218
Castellanos-Rozo J, Pérez Pulido R, Grande MJ, Lucas R, Gálvez A. Analysis of the Bacterial Diversity of Paipa Cheese (a Traditional Raw Cow’s Milk Cheese from Colombia) by High-Throughput Sequencing. Microorganisms. 2020; 8(2):218. https://doi.org/10.3390/microorganisms8020218
Chicago/Turabian StyleCastellanos-Rozo, José, Rubén Pérez Pulido, Mª. José Grande, Rosario Lucas, and Antonio Gálvez. 2020. "Analysis of the Bacterial Diversity of Paipa Cheese (a Traditional Raw Cow’s Milk Cheese from Colombia) by High-Throughput Sequencing" Microorganisms 8, no. 2: 218. https://doi.org/10.3390/microorganisms8020218
APA StyleCastellanos-Rozo, J., Pérez Pulido, R., Grande, M. J., Lucas, R., & Gálvez, A. (2020). Analysis of the Bacterial Diversity of Paipa Cheese (a Traditional Raw Cow’s Milk Cheese from Colombia) by High-Throughput Sequencing. Microorganisms, 8(2), 218. https://doi.org/10.3390/microorganisms8020218