pH and Phosphate Induced Shifts in Carbon Flow and Microbial Community during Thermophilic Anaerobic Digestion


Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Setup
2.2. Physio-Chemical Analyses
2.3. 16S RNA Gene Library Preparation and Amplicon Sequencing
2.4. Data Analysis and Statistics
3. Results
3.1. Starting Conditions and Microbial Community of the Inoculum
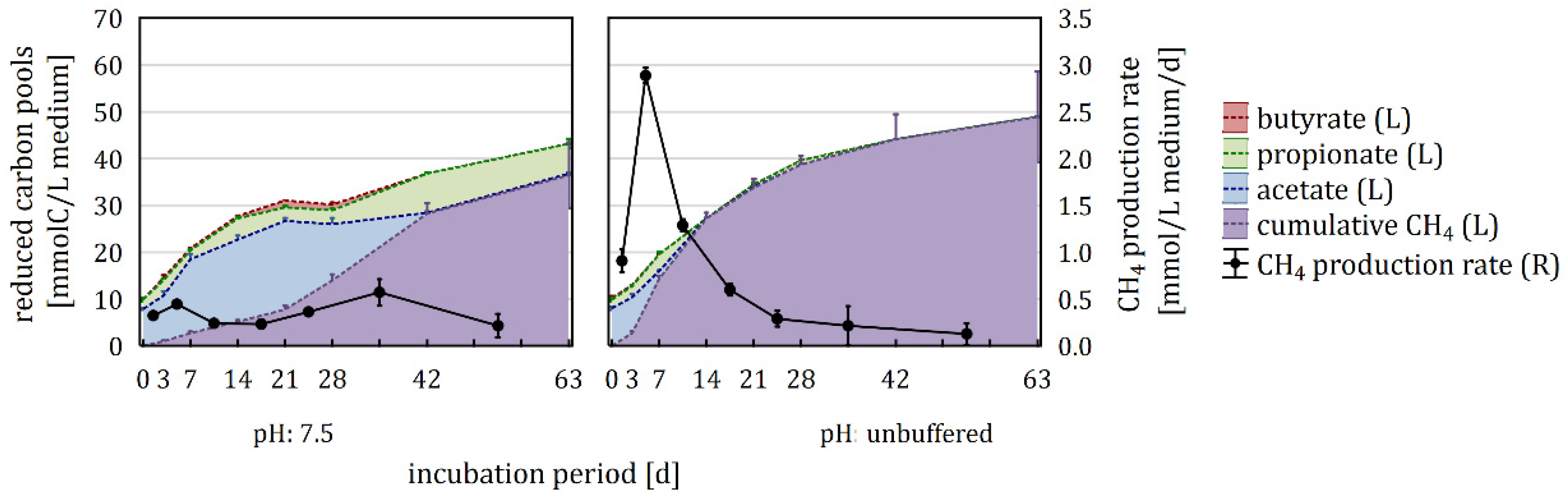
3.2. Impact of P-Buffer on the Carbon Flow
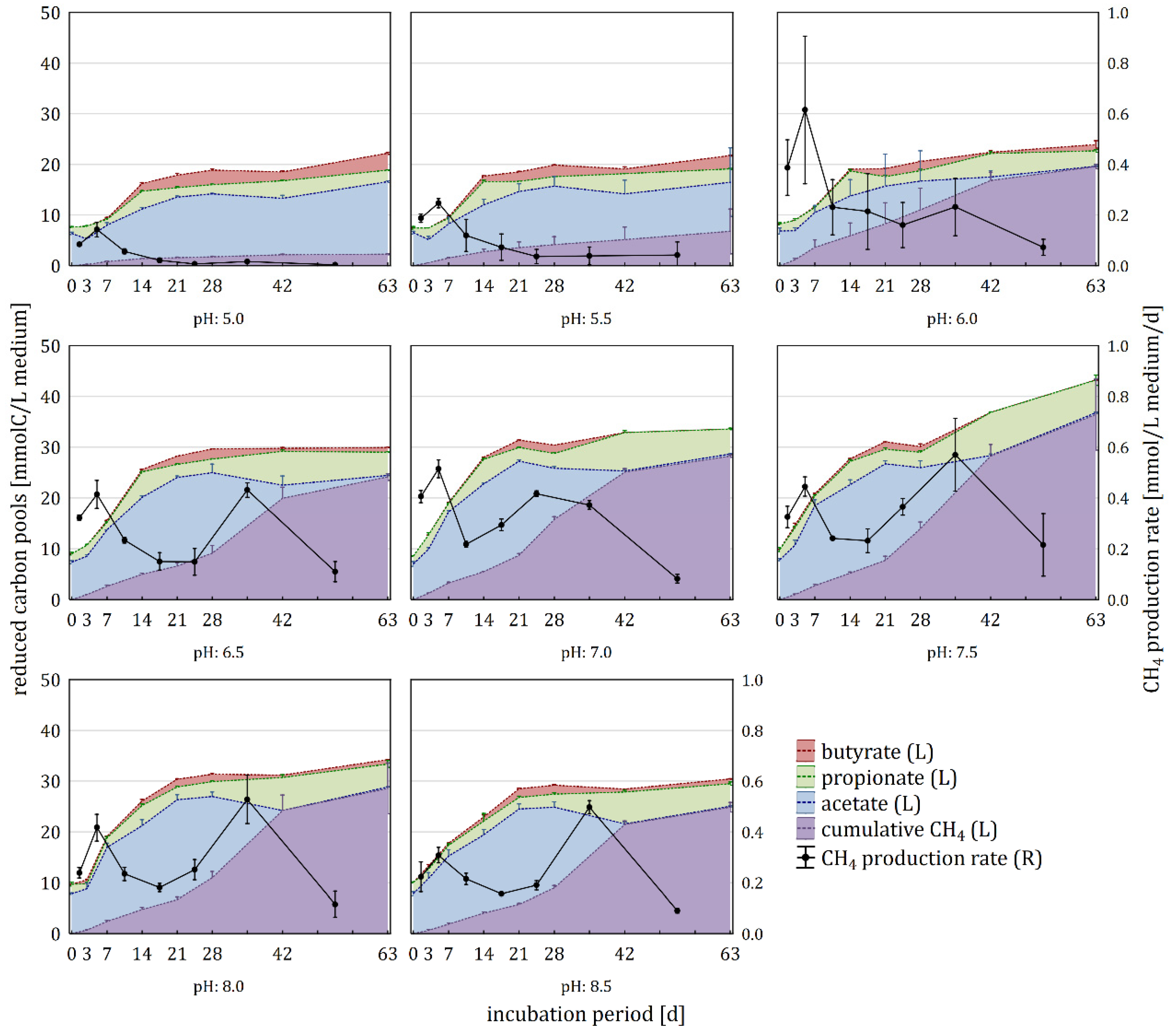
3.3. Effect of pH on Carbon Flow
3.4. Impact of the P-Buffer on the Microbial Community
3.5. Effect of pH on the Microbial Community
4. Discussion
4.1. Impact of P-Buffer
4.2. Effect of pH on Ecosystem Function and Microbial Diversity
4.3. Effect of pH on Physiochemical and Microbiological Aspects of the Anaerobic Digestion, Part 1: From Cellulose to Acetate
4.4. Effect of pH on Physiochemical and Microbiological Aspects of the Anaerobic Digestion, Part 2: From Acetate to CH4
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Siegrist, H.; Renggli, D.; Gujer, W. Mathematical Modelling of Anaerobic Mesophilic Sewage Sludge Treatment. Water Sci. Technol. 1993, 27, 25–36. [Google Scholar] [CrossRef]
- Lettinga, G. Anaerobic digestion and wastewater treatment systems. Antonie van Leeuwenhoek 1995, 67, 3–28. [Google Scholar] [CrossRef]
- Enzmann, F.; Mayer, F.; Rother, M.; Holtmann, D. Methanogens: Biochemical background and biotechnological applications. AMB Express 2018, 8, 1. [Google Scholar] [CrossRef] [PubMed]
- Miceli, J.F.; Torres, C.I.; Krajmalnik-Brown, R. Shifting the balance of fermentation products between hydrogen and volatile fatty acids: Microbial community structure and function. FEMS Microbiol. Ecol. 2016, 92. [Google Scholar] [CrossRef] [PubMed]
- Hoelzle, R.D.; Virdis, B.; Batstone, D.J. Regulation mechanisms in mixed and pure culture microbial fermentation. Biotechnol. Bioeng. 2014, 111, 2139–2154. [Google Scholar] [CrossRef]
- Levén, L.; Eriksson, A.R.B.; Schnürer, A. Effect of process temperature on bacterial and archaeal communities in two methanogenic bioreactors treating organic household waste. FEMS Microbiol. Ecol. 2007, 59, 683–693. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Sun, L.; Nordberg, Å.; Schnürer, A. Substrate-induced response in biogas process performance and microbial community relates back to inoculum source. Microorganisms 2018, 6, 80. [Google Scholar] [CrossRef] [Green Version]
- Campanaro, S.; Treu, L.; Kougias, P.G.; Luo, G.; Angelidaki, I. Metagenomic binning reveals the functional roles of core abundant microorganisms in twelve full-scale biogas plants. Water Res. 2018, 140, 123–134. [Google Scholar] [CrossRef]
- Nakamura, K.; Takahashi, A.; Mori, C.; Tamaki, H.; Mochimaru, H.; Nakamura, K.; Takamizawa, K.; Kamagata, Y. Methanothermobacter tenebrarum sp. nov., a hydrogenotrophic, thermophilic methanogen isolated from gas-associated formation water of a natural gas field. Int. J. Syst. Evol. Microbiol. 2013, 63, 715–722. [Google Scholar] [CrossRef]
- Smith, M.R.; Mah, R.A. Growth and methanogenesis by Methanosarcina strain 227 on acetate and methanol. Appl. Environ. Microbiol. 1978, 36, 870–879. [Google Scholar] [CrossRef] [Green Version]
- Witney, F.R.; Failla, M.L.; Weinberg, E.D. Phosphate inhibition of secondary metabolism in Serratia marcescens. Appl. Environ. Microbiol. 1977, 33, 1042–1046. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suzuki, S.; Shintani, M.; Sanchez, Z.K.; Kimura, K.; Numata, M.; Yamazoe, A.; Kimbara, K. Effects of phosphate addition on methane fermentation in the batch and upflow anaerobic sludge blanket (UASB) reactors. Appl. Microbiol. Biotechnol. 2015, 99, 10457–10466. [Google Scholar] [CrossRef] [PubMed]
- Lei, Z.; Chen, J.; Zhang, Z.; Sugiura, N. Methane production from rice straw with acclimated anaerobic sludge: Effect of phosphate supplementation. Bioresour. Technol. 2010, 101, 4343–4348. [Google Scholar] [CrossRef] [PubMed]
- Türker, M.; Celen, I. Removal of ammonia as struvite from anaerobic digester effluents and recycling of magnesium and phosphate. Bioresour. Technol. 2007, 98, 1529–1534. [Google Scholar] [CrossRef] [PubMed]
- Slonczewski, J.L.; Fujisawa, M.; Dopson, M.; Krulwich, T.A. Cytoplasmic pH Measurement and Homeostasis in Bacteria and Archaea; Elsevier: Amsterdam, The Netherlands, 2009; pp. 1–317. ISBN 9780123747907. [Google Scholar]
- Mao, C.; Feng, Y.; Wang, X.; Ren, G. Review on research achievements of biogas from anaerobic digestion. Renew. Sustain. Energy Rev. 2015, 45, 540–555. [Google Scholar] [CrossRef]
- Xu, F.; Li, Y.; Ge, X.; Yang, L.; Li, Y. Anaerobic digestion of food waste—Challenges and opportunities. Bioresour. Technol. 2018, 247, 1047–1058. [Google Scholar] [CrossRef]
- Graef, S.P.; Andrews, J.F. Stability and Control of Anaerobic Digestion. J. Water Pollut. Control Fed. 1974, 46, 666–683. [Google Scholar]
- Hao, L.-P.; Lü, F.; Li, L.; Shao, L.-M.; He, P.-J. Shift of pathways during initiation of thermophilic methanogenesis at different initial pH. Bioresour. Technol. 2012, 126, 418–424. [Google Scholar] [CrossRef]
- Zhai, N.; Zhang, T.; Yin, D.; Yang, G.; Wang, X.; Ren, G.; Feng, Y. Effect of initial pH on anaerobic co-digestion of kitchen waste and cow manure. Waste Manag. 2015, 38, 126–131. [Google Scholar] [CrossRef]
- Zhang, P.; Chen, Y.; Zhou, Q. Waste activated sludge hydrolysis and short-chain fatty acids accumulation under mesophilic and thermophilic conditions: Effect of pH. Water Res. 2009, 43, 3735–3742. [Google Scholar] [CrossRef]
- Trček, J.; Mira, N.P.; Jarboe, L.R. Adaptation and tolerance of bacteria against acetic acid. Appl. Microbiol. Biotechnol. 2015, 99, 6215–6229. [Google Scholar] [CrossRef] [PubMed]
- O’Flaherty, V.; Mahony, T.; O’Kennedy, R.; Colleran, E. Effect of pH on growth kinetics and sulphide toxicity thresholds of a range of methanogenic, syntrophic and sulphate-reducing bacteria. Process Biochem. 1998, 33, 555–569. [Google Scholar] [CrossRef]
- Zhou, J.; Zhang, R.; Liu, F.; Yong, X.; Wu, X.; Zheng, T.; Jiang, M.; Jia, H. Biogas production and microbial community shift through neutral pH control during the anaerobic digestion of pig manure. Bioresour. Technol. 2016, 217, 44–49. [Google Scholar] [CrossRef] [PubMed]
- Boaro, A.A.; Kim, Y.-M.; Konopka, A.E.; Callister, S.J.; Ahring, B.K. Integrated ‘omics analysis for studying the microbial community response to a pH perturbation of a cellulose-degrading bioreactor culture. FEMS Microbiol. Ecol. 2014, 90, 802–815. [Google Scholar] [CrossRef]
- Illmer, P.; Gstraunthaler, G. Effect of seasonal changes in quantities of biowaste on full scale anaerobic digester performance. Waste Manag. 2009, 29, 162–167. [Google Scholar] [CrossRef]
- Wagner, A.O.; Markt, R.; Mutschlechner, M.; Lackner, N.; Prem, E.M.; Praeg, N.; Illmer, P. Medium preparation for the cultivation of microorganisms under strictly anaerobic/anoxic conditions. J. Vis. Exp. 2019, 15. [Google Scholar] [CrossRef]
- Wagner, A.O.; Malin, C.; Lins, P.; Illmer, P. Effects of various fatty acid amendments on a microbial digester community in batch culture. Waste Manag. 2011, 31, 431–437. [Google Scholar] [CrossRef]
- Wagner, A.O.; Markt, R.; Puempel, T.; Illmer, P.; Insam, H.; Ebner, C. Sample preparation, preservation, and storage for volatile fatty acid quantification in biogas plants. Eng. Life Sci. 2017, 17, 132–139. [Google Scholar] [CrossRef]
- Wagner, A.O.; Prem, E.M.; Markt, R.; Kaufmann, R.; Illmer, P. Formation of phenylacetic acid and phenylpropionic acid under different overload conditions during mesophilic and thermophilic anaerobic digestion. Biotechnol. Biofuels 2019, 12, 359–383. [Google Scholar] [CrossRef] [PubMed]
- Apprill, A.; McNally, S.; Parsons, R.; Weber, L. Minor revision to V4 region SSU rRNA 806R gene primer greatly increases detection of SAR11 bacterioplankton. Aquat. Microb. Ecol. 2015, 75, 129–137. [Google Scholar] [CrossRef] [Green Version]
- Parada, A.E.; Needham, D.M.; Fuhrman, J.A. Every base matters: Assessing small subunit rRNA primers for marine microbiomes with mock communities, time series and global field samples. Environ. Microbiol. 2016, 18, 1403–1414. [Google Scholar] [CrossRef] [PubMed]
- Schloss, P.D.; Westcott, S.L.; Ryabin, T.; Hall, J.R.; Hartmann, M.; Hollister, E.B.; Lesniewski, R.A.; Oakley, B.B.; Parks, D.H.; Robinson, C.J.; et al. Introducing mothur: Open-source, platform-independent, community-supported software for describing and comparing microbial communities. Appl. Environ. Microbiol. 2009, 75, 7537–7541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2013, 41, D590–D596. [Google Scholar] [CrossRef] [PubMed]
- Yilmaz, P.; Parfrey, L.W.; Yarza, P.; Gerken, J.; Pruesse, E.; Quast, C.; Schweer, T.; Peplies, J.; Ludwig, W.; Glockner, F.O. The SILVA and “All-species living tree project (LTP)” taxonomic frameworks. Nucleic Acids Res. 2014, 42, D643–D648. [Google Scholar] [CrossRef] [Green Version]
- Rognes, T.; Flouri, T.; Nichols, B.; Quince, C.; Mahé, F. VSEARCH: A versatile open source tool for metagenomics. PeerJ 2016, 4, e2584. [Google Scholar] [CrossRef]
- Hammer, Ø.; Harper, D.A.T.; Ryan Paul, D. PAST: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 1–9. [Google Scholar]
- de Cáceres, M.; Legendre, P. Associations between species and groups of sites: Indices and statistical inference. Ecology 2009, 90, 3566–3574. [Google Scholar] [CrossRef]
- Huson, D.H.; Richter, D.C.; Rausch, C.; Dezulian, T.; Franz, M.; Rupp, R. Dendroscope: An interactive viewer for large phylogenetic trees. BMC Bioinform. 2007, 8. [Google Scholar] [CrossRef] [Green Version]
- Stoddard, S.F.; Smith, B.J.; Hein, R.; Roller, B.R.K.; Schmidt, T.M. rrnDB: Improved tools for interpreting rRNA gene abundance in bacteria and archaea and a new foundation for future development. Nucleic Acids Res. 2015, 43, D593–D598. [Google Scholar] [CrossRef] [Green Version]
- Wang, R.; Li, Y.; Wang, W.; Chen, Y.; Vanrolleghem, P.A. Effect of high orthophosphate concentration on mesophilic anaerobic sludge digestion and its modeling. Chem. Eng. J. 2015, 260, 791–800. [Google Scholar] [CrossRef]
- Conrad, R.; Klose, M.; Claus, P. Phosphate inhibits acetotrophic methanogenesis on rice roots. Appl. Environ. Microbiol. 2000, 66, 828–831. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paulo, P.L.; dos Santos, A.B.; Ide, C.N.; Lettinga, G. Phosphate inhibition on thermophilic acetoclastic methanogens: A warning. Water Sci. Technol. 2005, 52, 331–336. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Gil, G.; Kleerebezem, R.; Lettinga, G. Assessment of metabolic properties and kinetic parameters of methanogenic sludge by on-line methane production rate measurements. Appl. Microbiol. Biotechnol. 2002, 58, 248–254. [Google Scholar] [CrossRef]
- Kim, D.; Min, K.J.; Lee, K.; Yu, M.S.; Park, K.Y. Effects of pH, molar ratios and pre-treatment on phosphorus recovery through struvite crystallization from effluent of anaerobically digested swine wastewater. Environ. Eng. Res. 2017, 22, 12–18. [Google Scholar] [CrossRef]
- Wintsche, B.; Jehmlich, N.; Popp, D.; Harms, H.; Kleinsteuber, S. Metabolic adaptation of methanogens in anaerobic digesters upon trace element limitation. Front. Microbiol. 2018, 9, 405. [Google Scholar] [CrossRef]
- Choong, Y.Y.; Norli, I.; Abdullah, A.Z.; Yhaya, M.F. Impacts of trace element supplementation on the performance of anaerobic digestion process: A critical review. Bioresour. Technol. 2016, 209, 369–379. [Google Scholar] [CrossRef]
- Neubeck, A.; Sjöberg, S.; Price, A.; Callac, N.; Schnürer, A. Effect of nickel levels on hydrogen partial pressure and methane production in methanogens. PLoS ONE 2016, 11, e0168357. [Google Scholar] [CrossRef]
- Wintsche, B.; Glaser, K.; Sträuber, H.; Centler, F.; Liebetrau, J.; Harms, H.; Kleinsteuber, S. Trace elements induce predominance among methanogenic activity in anaerobic digestion. Front. Microbiol. 2016, 7, 2034. [Google Scholar] [CrossRef]
- Šafarič, L.; Shakeri Yekta, S.; Liu, T.; Svensson, B.H.; Schnürer, A.; Bastviken, D.; Björn, A. Dynamics of a perturbed microbial community during thermophilic anaerobic digestion of chemically definded soluble organic compounds. Microorganisms 2018, 6, 105. [Google Scholar] [CrossRef] [Green Version]
- Banks, C.J.; Zhang, Y.; Jiang, Y.; Heaven, S. Trace element requirements for stable food waste digestion at elevated ammonia concentrations. Bioresour. Technol. 2012, 104, 127–135. [Google Scholar] [CrossRef] [Green Version]
- Guo, X.; Wang, C.; Sun, F.; Zhu, W.; Wu, W. A comparison of microbial characteristics between the thermophilic and mesophilic anaerobic digesters exposed to elevated food waste loadings. Bioresour. Technol. 2014, 152, 420–428. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Han, G.; Shin, S.G.; Koo, T.; Cho, K.; Kim, W.; Hwang, S. Seasonal monitoring of bacteria and archaea in a full-scale thermophilic anaerobic digester treating food waste-recycling wastewater: Correlations between microbial community characteristics and process variables. Chem. Eng. J. 2016, 300, 291–299. [Google Scholar] [CrossRef]
- Liang, X.; Whitham, J.M.; Holwerda, E.K.; Shao, X.; Tian, L.; Wu, Y.-W.; Lombard, V.; Henrissat, B.; Klingeman, D.M.; Yang, Z.K.; et al. Development and characterization of stable anaerobic thermophilic methanogenic microbiomes fermenting switchgrass at decreasing residence times. Biotechnol. Biofuels 2018, 11, 243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maus, I.; Kim, Y.S.; Wibberg, D.; Stolze, Y.; Off, S.; Antonczyk, S.; Pühler, A.; Scherer, P.; Schlüter, A. Biphasic study to characterize agricultural biogas plants by high-throughput 16S rRNA gene amplicon sequencing and microscopic analysis. J. Microbiol. Biotechnol. 2017, 27, 321–334. [Google Scholar] [CrossRef] [Green Version]
- Zamanzadeh, M.; Hagen, L.H.; Svensson, K.; Linjordet, R.; Horn, S.J. Anaerobic digestion of food waste—Effect of recirculation and temperature on performance and microbiology. Water Res. 2016, 96, 246–254. [Google Scholar] [CrossRef]
- Fontana, A.; Patrone, V.; Puglisi, E.; Morelli, L.; Bassi, D.; Garuti, M.; Rossi, L.; Cappa, F. Effects of geographic area, feedstock, temperature, and operating time on microbial communities of six full-scale biogas plants. Bioresour. Technol. 2016, 218, 980–990. [Google Scholar] [CrossRef] [PubMed]
- Pap, B.; Györkei, Á.; Boboescu, I.Z.; Nagy, I.K.; Bíró, T.; Kondorosi, É.; Maróti, G. Temperature-dependent transformation of biogas-producing microbial communities points to the increased importance of hydrogenotrophic methanogenesis under thermophilic operation. Bioresour. Technol. 2015, 177, 375–380. [Google Scholar] [CrossRef]
- Parks, D.H.; Rinke, C.; Chuvochina, M.; Chaumeil, P.-A.; Woodcroft, B.J.; Evans, P.N.; Hugenholtz, P.; Tyson, G.W. Recovery of nearly 8,000 metagenome-assembled genomes substantially expands the tree of life. Nat. Microbiol. 2017, 2, 1533–1542. [Google Scholar] [CrossRef]
- McCann, K.S. The diversity–stability debate. Nature 2000, 405, 228–233. [Google Scholar] [CrossRef]
- Shade, A.; Peter, H.; Allison, S.D.; Baho, D.L.; Berga, M.; Bürgmann, H.; Huber, D.H.; Langenheder, S.; Lennon, J.T.; Martiny, J.B.H.; et al. Fundamentals of microbial community resistance and resilience. Front. Microbiol. 2012, 3, 417. [Google Scholar] [CrossRef] [Green Version]
- Maune, M.W.; Tanner, R.S. Description of Anaerobaculum hydrogeniformans sp. nov., an anaerobe that produces hydrogen from glucose, and emended description of the genus Anaerobaculum. Int. J. Syst. Evol. Microbiol. 2012, 62, 832–838. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Menes, R.J.; Muxí, L. Anaerobaculum mobile sp. nov., a novel anaerobic, moderately thermophilic, peptide-fermenting bacterium that uses crotonate as an electron acceptor, and emended description of the genus Anaerobaculum. Int. J. Syst. Evol. Microbiol. 2002, 52, 157–164. [Google Scholar] [CrossRef] [PubMed]
- Rees, G.N.; Patel, B.K.; Grassia, G.S.; Sheehy, A.J. Anaerobaculum thermoterrenum gen. nov., sp. nov., a novel, thermophilic bacterium which ferments citrate. Int. J. Syst. Bacteriol. 1997, 47, 150–154. [Google Scholar] [CrossRef]
- Zhang, X.; Tu, B.; Dai, L.-R.; Lawson, P.A.; Zheng, Z.-Z.; Liu, L.-Y.; Deng, Y.; Zhang, H.; Cheng, L. Petroclostridium xylanilyticum gen. nov., sp. nov., a xylan-degrading bacterium isolated from an oilfield, and reclassification of clostridial cluster III members into four novel genera in a new Hungateiclostridiaceae fam. nov. Int. J. Syst. Evol. Microbiol. 2018, 68, 3197–3211. [Google Scholar] [CrossRef] [PubMed]
- Hania, W.B.; Godbane, R.; Postec, A.; Hamdi, M.; Ollivier, B.; Fardeau, M.-L. Defluviitoga tunisiensis gen. nov., sp. nov., a thermophilic bacterium isolated from a mesothermic and anaerobic whey digester. Int. J. Syst. Evol. Microbiol. 2012, 62, 1377–1382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koeck, D.E.; Ludwig, W.; Wanner, G.; Zverlov, V.V.; Liebl, W.; Schwarz, W.H. Herbinix hemicellulosilytica gen. nov., sp. nov., a thermophilic cellulose-degrading bacterium isolated from a thermophilic biogas reactor. Int. J. Syst. Evol. Microbiol. 2015, 65, 2365–2371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koeck, D.E.; Hahnke, S.; Zverlov, V.V. Herbinix luporum sp. nov., a thermophilic cellulose-degrading bacterium isolated from a thermophilic biogas reactor. Int. J. Syst. Evol. Microbiol. 2016, 66, 4132–4137. [Google Scholar] [CrossRef]
- Slobodkin, A.I.; Tourova, T.P.; Kostrikina, N.A.; Lysenko, A.M.; German, K.E.; Bonch-Osmolovskaya, E.A.; Birkeland, N.-K. Tepidimicrobium ferriphilum gen. nov., sp. nov., a novel moderately thermophilic, Fe(III)-reducing bacterium of the order Clostridiales. Int. J. Syst. Evol. Microbiol. 2006, 56, 369–372. [Google Scholar] [CrossRef] [Green Version]
- Etchebehere, C.; Pavan, M.E.; Zorzópulos, J.; Soubes, M.; Muxí, L. Coprothermobacter platensis sp. nov., a new anaerobic proteolytic thermophilic bacterium isolated from an anaerobic mesophilic sludge. Int. J. Syst. Bacteriol. 1998, 48 Pt 4, 1297–1304. [Google Scholar] [CrossRef] [Green Version]
- Ollivier, B.M.; Mah, R.A.; Ferguson, T.J.; Boone, D.R.; Garcia, J.L.; Robinson, R. Emendation of the genus Thermobacteroides: Thermobacteroides proteolyticus sp. nov., a proteolytic acetogen from a methanogenic enrichment. Int. J. Syst. Bacteriol. 1985, 35, 425–428. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.; Dong, X. Proteiniphilum acetatigenes gen. nov., sp. nov., from a UASB reactor treating brewery wastewater. Int. J. Syst. Evol. Microbiol. 2005, 55, 2257–2261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kublanov, I.V.; Prokofeva, M.I.; Kostrikina, N.A.; Kolganova, T.V.; Tourova, T.P.; Wiegel, J.; Bonch-Osmolovskaya, E.A. Thermoanaerobacterium aciditolerans sp. nov., a moderate thermoacidophile from a Kamchatka hot spring. Int. J. Syst. Evol. Microbiol. 2007, 57, 260–264. [Google Scholar] [CrossRef] [PubMed]
- Shang, S.-M.; Qian, L.; Zhang, X.; Li, K.-Z.; Chagan, I. Themoanaerobacterium calidifontis sp. nov., a novel anaerobic, thermophilic, ethanol-producing bacterium from hot springs in China. Arch. Microbiol. 2013, 195, 439–445. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Romano, I.; Dipasquale, L.; Orlando, P.; Lama, L.; d’Ippolito, G.; Pascual, J.; Gambacorta, A. Thermoanaerobacterium thermostercus sp. nov., a new anaerobic thermophilic hydrogen-producing bacterium from buffalo-dung. Extremophiles 2010, 14, 233–240. [Google Scholar] [CrossRef]
- Liu, S.Y.; Rainey, F.A.; Morgan, H.W.; Mayer, F.; Wiegel, J. Thermoanaerobacterium aotearoense sp. nov., a slightly acidophilic, anaerobic thermophile isolated from various hot springs in new zealand, and emendation of the genus Thermoanaerobacterium. Int. J. Syst. Bacteriol. 1996, 46, 388–396. [Google Scholar] [CrossRef] [Green Version]
- Kim, B.-C.; Seung Jeon, B.; Kim, S.; Kim, H.; Um, Y.; Sang, B.-I. Caproiciproducens galactitolivorans gen. nov., sp. nov., a bacterium capable of producing caproic acid from galactitol, isolated from a wastewater treatment plant. Int. J. Syst. Evol. Microbiol. 2015, 65, 4902–4908. [Google Scholar] [CrossRef]
- Lee, Y.-J.; Mackie, R.I.; Cann, I.K.O.; Wiegel, J. Description of Caldanaerobius fijiensis gen. nov., sp. nov., an inulin-degrading, ethanol-producing, thermophilic bacterium from a Fijian hot spring sediment, and reclassification of Thermoanaerobacterium polysaccharolyticum and Thermoanaerobacterium zeae as Caldanaerobius polysaccharolyticus comb. nov. and Caldanaerobius zeae comb. nov. Int. J. Syst. Evol. Microbiol. 2008, 58, 666–670. [Google Scholar] [CrossRef] [Green Version]
- Cann, I.K.; Stroot, P.G.; Mackie, K.R.; White, B.A.; Mackie, R.I. Characterization of two novel saccharolytic, anaerobic thermophiles, Thermoanaerobacterium polysaccharolyticum sp. nov. and Thermoanaerobacterium zeae sp. nov., and emendation of the genus Thermoanaerobacterium. Int. J. Syst. Evol. Microbiol. 2001, 51, 293–302. [Google Scholar] [CrossRef] [Green Version]
- Morotomi, M.; Nagai, F.; Watanabe, Y. Description of Christensenella minuta gen. nov., sp. nov., isolated from human faeces, which forms a distinct branch in the order Clostridiales, and proposal of Christensenellaceae fam. nov. Int. J. Syst. Evol. Microbiol. 2012, 62, 144–149. [Google Scholar] [CrossRef]
- Yokoyama, H.; Wagner, I.D.; Wiegel, J. Caldicoprobacter oshimai gen. nov., sp. nov., an anaerobic, xylanolytic, extremely thermophilic bacterium isolated from sheep faeces, and proposal of Caldicoprobacteraceae fam. nov. Int. J. Syst. Evol. Microbiol. 2010, 60, 67–71. [Google Scholar] [CrossRef] [Green Version]
- Bouanane-Darenfed, A.; Ben Hania, W.; Cayol, J.-L.; Ollivier, B.; Fardeau, M.-L. Reclassification of Acetomicrobium faecale as Caldicoprobacter faecalis comb. nov. Int. J. Syst. Evol. Microbiol. 2015, 65, 3286–3288. [Google Scholar] [CrossRef] [PubMed]
- Bouanane-Darenfed, A.; Ben Hania, W.; Hacene, H.; Cayol, J.-L.; Ollivier, B.; Fardeau, M.-L. Caldicoprobacter guelmensis sp. nov., a thermophilic, anaerobic, xylanolytic bacterium isolated from a hot spring. Int. J. Syst. Evol. Microbiol. 2013, 63, 2049–2053. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bouanane-Darenfed, A.; Fardeau, M.-L.; Grégoire, P.; Joseph, M.; Kebbouche-Gana, S.; Benayad, T.; Hacene, H.; Cayol, J.-L.; Ollivier, B. Caldicoprobacter algeriensis sp. nov. a new thermophilic anaerobic, xylanolytic bacterium isolated from an Algerian hot spring. Curr. Microbiol. 2011, 62, 826–832. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Qiao, J.-T.; Yuan, X.-Z.; Guo, R.-B.; Qiu, Y.-L. Hydrogenispora ethanolica gen. nov., sp. nov., an anaerobic carbohydrate-fermenting bacterium from anaerobic sludge. Int. J. Syst. Evol. Microbiol. 2014, 64, 1756–1762. [Google Scholar] [CrossRef] [Green Version]
- Sun, L.; Toyonaga, M.; Ohashi, A.; Tourlousse, D.M.; Matsuura, N.; Meng, X.-Y.; Tamaki, H.; Hanada, S.; Cruz, R.; Yamaguchi, T.; et al. Lentimicrobium saccharophilum gen. nov., sp. nov., a strictly anaerobic bacterium representing a new family in the phylum Bacteroidetes, and proposal of Lentimicrobiaceae fam. nov. Int. J. Syst. Evol. Microbiol. 2016, 66, 2635–2642. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, C.; Liu, X.; Dong, X. Syntrophomonas cellicola sp. nov., a spore-forming syntrophic bacterium isolated from a distilled-spirit-fermenting cellar, and assignment of Syntrophospora bryantii to Syntrophomonas bryantii comb. nov. Int. J. Syst. Evol. Microbiol. 2006, 56, 2331–2335. [Google Scholar] [CrossRef] [PubMed]
- Stieb, M.; Schink, B. Anaerobic oxidation of fatty acids by Clostridium bryantii sp. nov., a sporeforming, obligately syntrophic bacterium. Arch. Microbiol. 1985, 140, 387–390. [Google Scholar] [CrossRef] [Green Version]
- McInerney, M.J.; Bryant, M.P.; Hespell, R.B.; Costerton, J.W. Syntrophomonas wolfei gen. nov. sp. nov., an anaerobic, syntrophic, fatty acid-oxidizing bacterium. Appl. Environ. Microbiol. 1981, 41, 1029–1039. [Google Scholar] [CrossRef] [Green Version]
- Hatamoto, M.; Imachi, H.; Fukayo, S.; Ohashi, A.; Harada, H. Syntrophomonas palmitatica sp. nov., an anaerobic, syntrophic, long-chain fatty-acid-oxidizing bacterium isolated from methanogenic sludge. Int. J. Syst. Evol. Microbiol. 2007, 57, 2137–2142. [Google Scholar] [CrossRef]
- Zhang, C.; Liu, X.; Dong, X. Syntrophomonas curvata sp. nov., an anaerobe that degrades fatty acids in co-culture with methanogens. Int. J. Syst. Evol. Microbiol. 2004, 54, 969–973. [Google Scholar] [CrossRef] [Green Version]
- Roy, F.; Samain, E.; Dubourguier, H.C.; Albagnac, G. Synthrophomonas sapovorans sp. nov., a new obligately proton reducing anaerobe oxidizing saturated and unsaturated long chain fatty acids. Arch. Microbiol. 1986, 145, 142–147. [Google Scholar] [CrossRef]
- Westerholm, M.; Roos, S.; Schnürer, A. Syntrophaceticus schinkii gen. nov., sp. nov., an anaerobic, syntrophic acetate-oxidizing bacterium isolated from a mesophilic anaerobic filter. FEMS Microbiol. Lett. 2010, 309, 100–104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manzoor, S.; Müller, B.; Niazi, A.; Schnürer, A.; Bongcam-Rudloff, E. Working draft genome sequence of the mesophilic acetate oxidizing bacterium Syntrophaceticus schinkii strain Sp3. Stand. Genom. Sci. 2015, 10, 99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zinder, S.H.; Mah, R.A. Isolation and characterization of a thermophilic strain of Methanosarcina unable to use H2-CO2 for methanogenesis. Appl. Environ. Microbiol. 1979, 38, 996–1008. [Google Scholar] [CrossRef] [Green Version]
- Sowers, K.R.; Baron, S.F.; Ferry, J.G. Methanosarcina acetivorans sp. nov., an acetoclastic methane-producing bacterium isolated from marine sediments. Appl. Environ. Microbiol. 1984, 47, 971–978. [Google Scholar] [CrossRef] [Green Version]
- Kern, T.; Fischer, M.A.; Deppenmeier, U.; Schmitz, R.A.; Rother, M. Methanosarcina flavescens sp. nov., a methanogenic archaeon isolated from a full-scale anaerobic digester. Int. J. Syst. Evol. Microbiol. 2016, 66, 1533–1538. [Google Scholar] [CrossRef]
- Wasserfallen, A.; Nölling, J.; Pfister, P.; Reeve, J.; Conway de Macario, E. Phylogenetic analysis of 18 thermophilic Methanobacterium isolates supports the proposals to create a new genus, Methanothermobacter gen. nov., and to reclassify several isolates in three species, Methanothermobacter thermautotrophicus comb. nov., Methanothermobacter wolfeii comb. nov., and Methanothermobacter marburgensis sp. nov. Int. J. Syst. Evol. Microbiol. 2000, 50 Pt 1, 43–53. [Google Scholar] [CrossRef]
- Cheng, L.; Qiu, T.-L.; Li, X.; Wang, W.-D.; Deng, Y.; Yin, X.-B.; Zhang, H. Isolation and characterization of Methanoculleus receptaculi sp. nov. from Shengli oil field, China. FEMS Microbiol.Lett. 2008, 285, 65–71. [Google Scholar] [CrossRef] [Green Version]
- Rivard, C.J.; Smith, P.H. Isolation and characterization of a thermophilic marine methanogenic bacterium, Methanogenium thermophilicum sp. nov. Int. J. Syst. Bacteriol. 1982, 32, 430–436. [Google Scholar] [CrossRef] [Green Version]
- Harned, H.S.; Ehlers, R.W. The dissociation constant of acetic acid from 0 to 60° centigrade. J. Am. Chem. Soc. 1933, 55, 652–656. [Google Scholar] [CrossRef]
- Rodriguez, J.; Kleerebezem, R.; Lema, J.M.; van Loosdrecht, M.C.M. Modeling product formation in anaerobic mixed culture fermentations. Biotechnol. Bioeng. 2006, 93, 592–606. [Google Scholar] [CrossRef] [PubMed]
- Gagliano, M.C.; Braguglia, C.M.; Petruccioli, M.; Rossetti, S. Ecology and biotechnological potential of the thermophilic fermentative Coprothermobacter spp. FEMS Microbiol. Ecol. 2015, 91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lü, F.; Bize, A.; Guillot, A.; Monnet, V.; Madigou, C.; Chapleur, O.; Mazéas, L.; He, P.; Bouchez, T. Metaproteomics of cellulose methanisation under thermophilic conditions reveals a surprisingly high proteolytic activity. ISME J. 2013, 8, 88–102. [Google Scholar] [CrossRef] [PubMed]
- Rainey, F.A.; Stackebrandt, E. Transfer of the type species of the genus Thermobacteroides to the genus Thermoanaerobacter as Thermoanaerobacter acetoethylicus (Ben-Bassat and Zeikus 1981) comb. nov., description of Coprothermobacter gen. nov., and reclassification of Thermobacteroides proteolyticus as Coprothermobacter proteolyticus (Ollivier et al. 1985) comb. nov. Int. J. Syst. Evol. Microbiol. 1993, 43, 857–859. [Google Scholar]
- Westerholm, M.; Roos, S.; Schnürer, A. Tepidanaerobacter acetatoxydans sp. nov., an anaerobic, syntrophic acetate-oxidizing bacterium isolated from two ammonium-enriched mesophilic methanogenic processes. Syst. Appl. Microbiol. 2011, 34, 260–266. [Google Scholar] [CrossRef] [PubMed]
- Westerholm, M.; Müller, B.; Singh, A.; Karlsson Lindsjö, O.; Schnürer, A. Detection of novel syntrophic acetate-oxidizing bacteria from biogas processes by continuous acetate enrichment approaches. Microb. Biotechnol. 2018, 11, 680–693. [Google Scholar] [CrossRef] [Green Version]
- Demirel, B.; Scherer, P. The roles of acetotrophic and hydrogenotrophic methanogens during anaerobic conversion of biomass to methane: A review. Rev. Environ. Sci. Biotechnol. 2008, 7, 173–190. [Google Scholar] [CrossRef]
- Illmer, P.; Reitschuler, C.; Wagner, A.O.; Schwarzenauer, T.; Lins, P. Microbial succession during thermophilic digestion: The potential of Methanosarcina sp. PLoS ONE 2014, 9, e86967. [Google Scholar] [CrossRef] [Green Version]
- Lins, P.; Reitschuler, C.; Illmer, P. Methanosarcina spp., the key to relieve the start-up of a thermophilic anaerobic digestion suffering from high acetic acid loads. Bioresour. Technol. 2014, 152, 347–354. [Google Scholar] [CrossRef]
- Hattori, S. Syntrophic acetate-oxidizing microbes in methanogenic environments. Microbes Environ. 2008, 23, 118–127. [Google Scholar] [CrossRef] [Green Version]
- Capson-Tojo, G.; Moscoviz, R.; Astals, S.; Robles, Á.; Steyer, J.-P. Unraveling the literature chaos around free ammonia inhibition in anaerobic digestion. Renew. Sustain. Energy Rev. 2020, 117, 109487. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Main Metabolic Task | OTU at Genus Level | This Study (data from day 63) | Literature | Source | ||
|---|---|---|---|---|---|---|
| Max. Rel. Abund. (respective pH) | Indicator for pH | pH Range * | Main Products ** | |||
| Hydrolysis of | Acetomicrobium | 9% (5.5) | 5.5 | 5–9 | Ac, H2 | [62,63,64] |
| cellulose and | Ruminiclostridium | 2% (6–7) | 6.0–6.5 | 5–8 | Ac, Pr, Bu, La, EtOH, H2 | [65] |
| acidogenesis | Defluviitoga | 39% (5.5) | 5.5–6.5 | 7–8 | Ac, H2 | [66] |
| Herbinix | 4% (7) | - | 7–10 | Ac, Pr, Bu, EtOH, H2 | [67,68] | |
| Hydrolysis of | Tepidimicrobium | 1% (5.5; 8.5) | - | 6–10 | Ac, Pr | [69] |
| proteins and | Coprothermobacter | 22% (5.5) | 5.5–6.5 | 4–9 | Ac, Pr, H2 | [70,71] |
| acidogenesis | Proteiniphilum | 6% (8.5) | - | 6–10 | Ac, Pr | [72] |
| Acidogenesis | Thermoanaerobacterium | 8% (5.5) | 5.5 | 3–8 | Ac, La, EtOH, H2 | [73,74,75,76] |
| Caproicipruducens | 2% (5.5) | 5.5 | 6–8 | Ac, Bu, Ca, EtOH, H2 | [77] | |
| Caldanerobius | 1% (5.5) | 5.5 | 4–8 | Fo, Ac, La, EtOH | [78,79] | |
| unc. Christensenellaceae | 3% (6.5) | - | 6–9 | Ac, Bu | [80] | |
| Caldicoprobacter | 12% (7.5) | 7.0–8.5 | 5–9 | Ac, La, EtOH, H2 | [81,82,83,84] | |
| Hydrogenispora | 5% (8.5) | 7.0–8.5 | 5–8 | Ac, EtOH, H2 | [85] | |
| Lentimicrobiaceae | 9% (8.5) | - | 7–9 | Fo, Ac, Pr, Ma, H2 | [86] | |
| Oxidation of VFA | unc. Syntrophomonadaceae | 1% (8.5) | 7.0–8.5 | 6–9 | Ac, Pr, H2 | [87,88,89,90,91,92] |
| SAO | Syntrophaceticus | 2% (7; 8.5) | 7.0–8.5 | 6–8 | Fo, H2 | [93,94] |
| AM | Methanosarcina | 16% (6) | 5.5–6.5 | 5–9 | CH4 | [95,96,97] |
| HM | Methanothermobacter | 4% (5.5) | - | 5–9 | CH4 | [9,98] |
| Methanoculleus | 8% (7.5) | - | 6–9 | CH4 | [99,100] | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lackner, N.; Wagner, A.O.; Markt, R.; Illmer, P. pH and Phosphate Induced Shifts in Carbon Flow and Microbial Community during Thermophilic Anaerobic Digestion. Microorganisms 2020, 8, 286. https://doi.org/10.3390/microorganisms8020286
Lackner N, Wagner AO, Markt R, Illmer P. pH and Phosphate Induced Shifts in Carbon Flow and Microbial Community during Thermophilic Anaerobic Digestion. Microorganisms. 2020; 8(2):286. https://doi.org/10.3390/microorganisms8020286
Chicago/Turabian StyleLackner, Nina, Andreas O. Wagner, Rudolf Markt, and Paul Illmer. 2020. "pH and Phosphate Induced Shifts in Carbon Flow and Microbial Community during Thermophilic Anaerobic Digestion" Microorganisms 8, no. 2: 286. https://doi.org/10.3390/microorganisms8020286
APA StyleLackner, N., Wagner, A. O., Markt, R., & Illmer, P. (2020). pH and Phosphate Induced Shifts in Carbon Flow and Microbial Community during Thermophilic Anaerobic Digestion. Microorganisms, 8(2), 286. https://doi.org/10.3390/microorganisms8020286