Eukaryotic Cell Permeabilisation to Identify New Putative Chlamydial Type III Secretion System Effectors Secreted within Host Cell Cytoplasm



Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture and Bacterial Strains
2.2. W. chondrophila Infection Procedure
2.3. C. trachomatis Infection Procedure
2.4. Selective Permeabilisation with Perfringolysin o and Mass Spectrometry Analysis
2.5. Yersinia enterocolitica Type III Secretion Assays
2.6. Ectopic Expression in HEK 293T Cells
2.7. Purification of Wcw_1706 for Pull Down Experiments and Antibodies Production
2.8. Pull Down Experiments
2.9. Immunoblots
2.9.1. Y. enterocolitica Secretion Assays
2.9.2. W. chondrophila Infected Cells
2.9.3. Pull Down Experiments
2.10. Gene Expression
- 16S for: 5′ GGCCCTTGGGTCGTAAAGTTCT 3′
- 16S rev: 5′ CGGAGTTAGCCGGTGCTTCT 3′
- wcw_1706 for: 5’ TTGACGCTTGTCGAGGTTCA 3’
- wcw_1706 rev: 5’ GCAAAAACTCCGGCACTTCC 3’
- wcw_0499 for: 5’ TGTGCGTGAGTTTTCAGAGGA 3’
- wcw_0499 rev: 5’ TTTATTGGTTTGCAGGGCGC 3’
2.11. Immunofluorescence and Confocal Microscopy
3. Results
3.1. Selective Eukaryotic Membrane Permeabilisation Identifies Secreted Bacterial Proteins
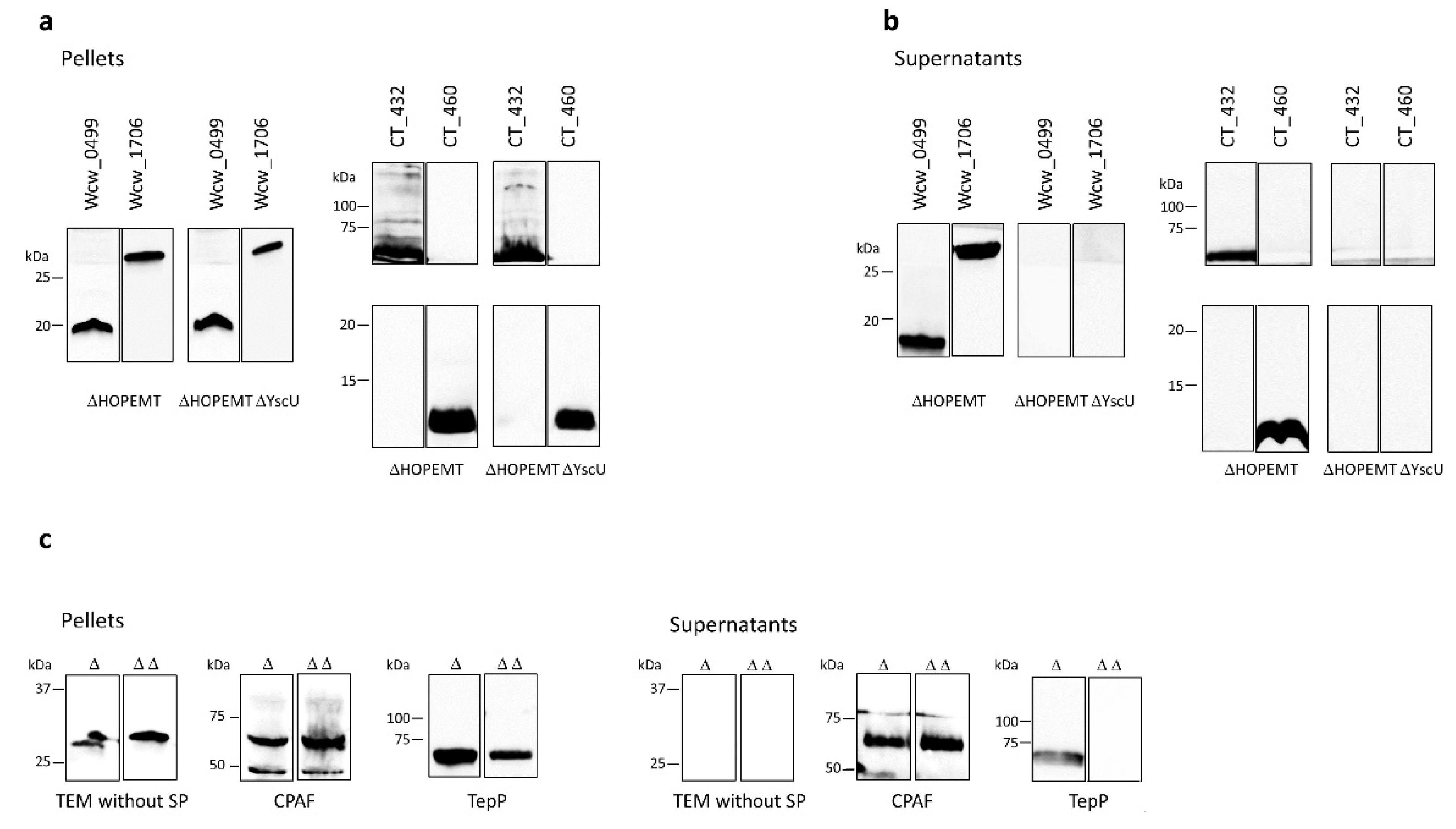
3.2. Four Newly Identified Putative Chlamydial Effector Proteins Are Secreted by Yersinia enterocolitica T3SS
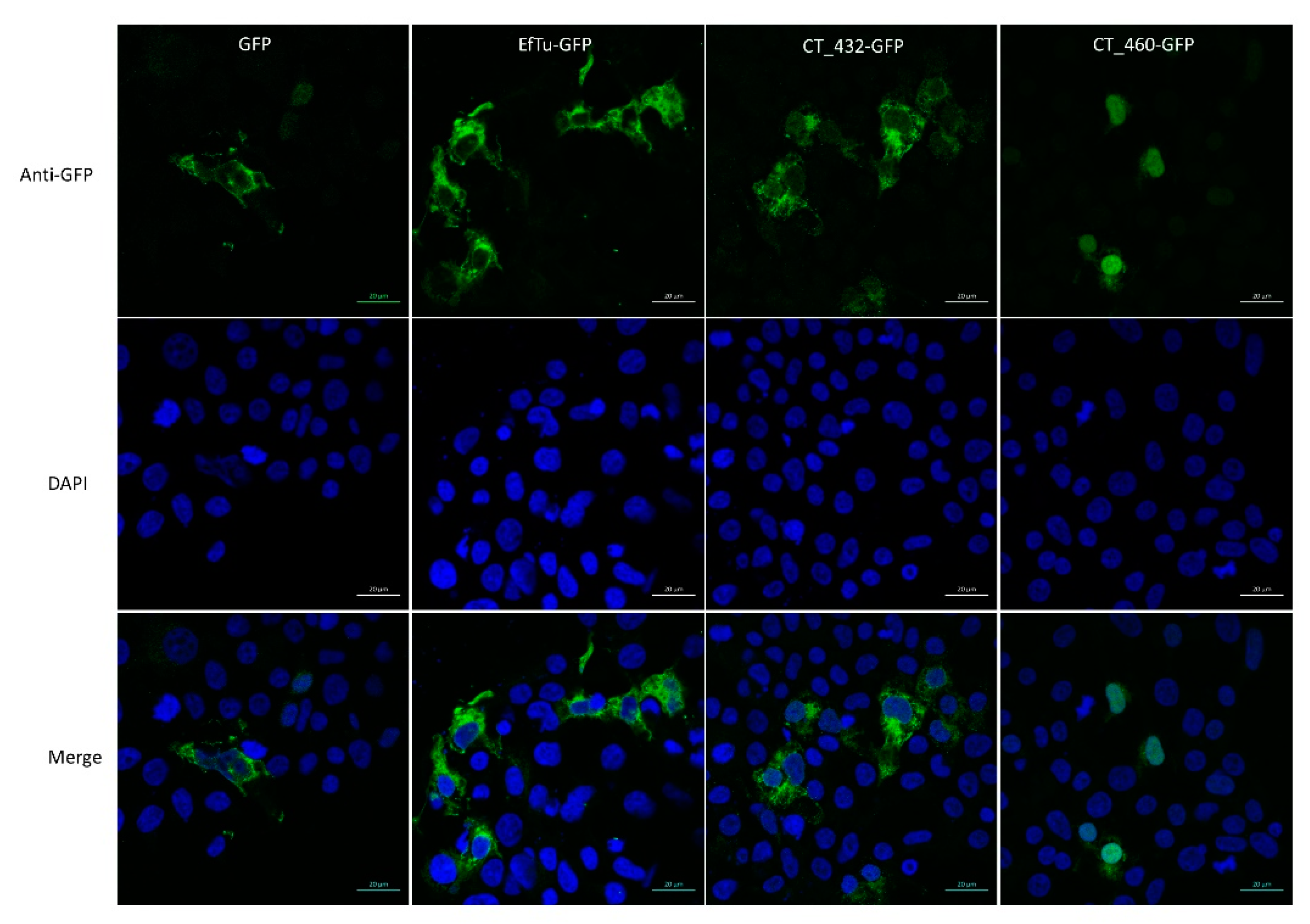
3.3. CT460 Localises in Host Cell Nucleus upon Transfection
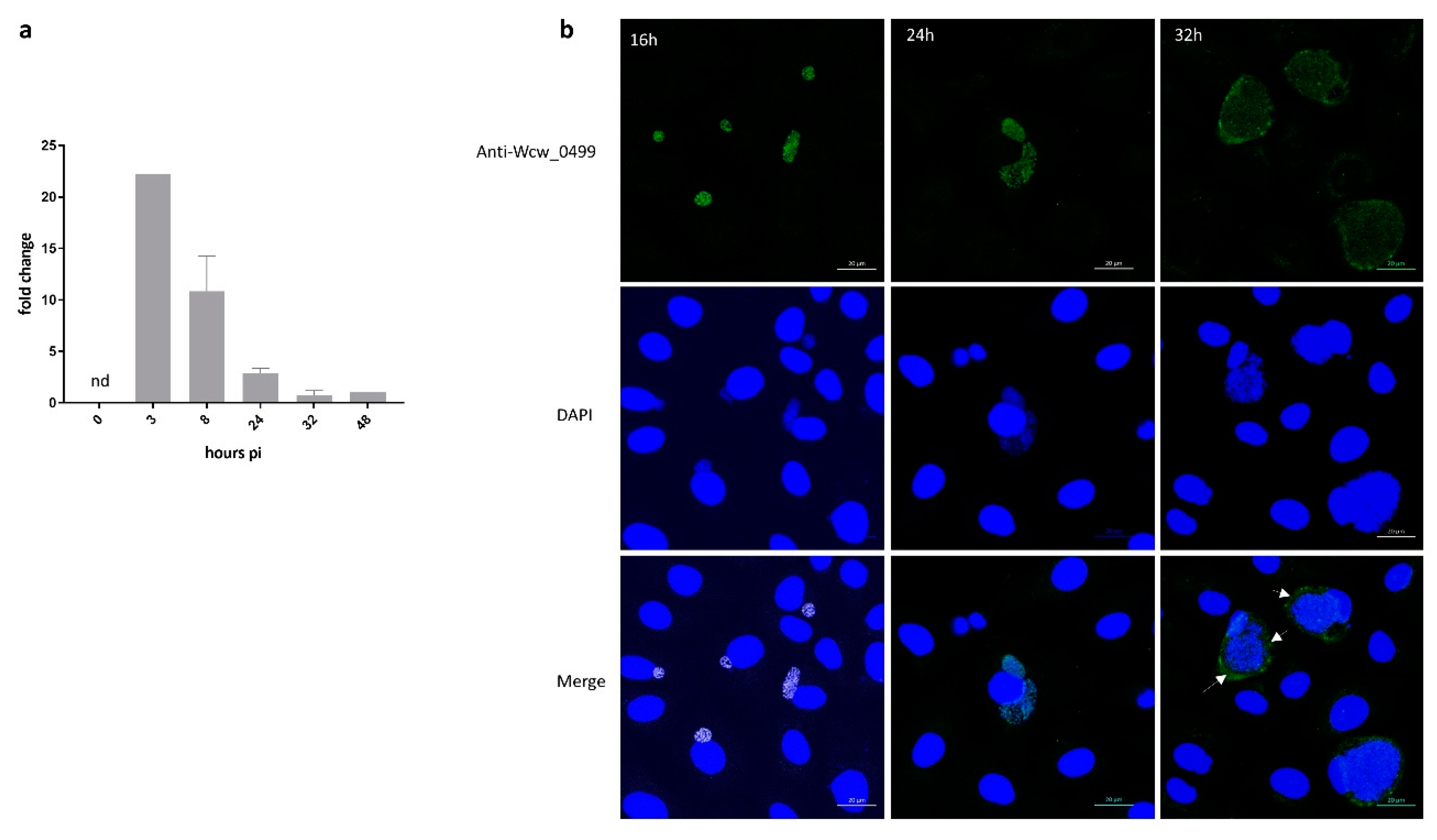
3.4. Wcw_0499 Is an Early to Mid-Cycle Effector Protein
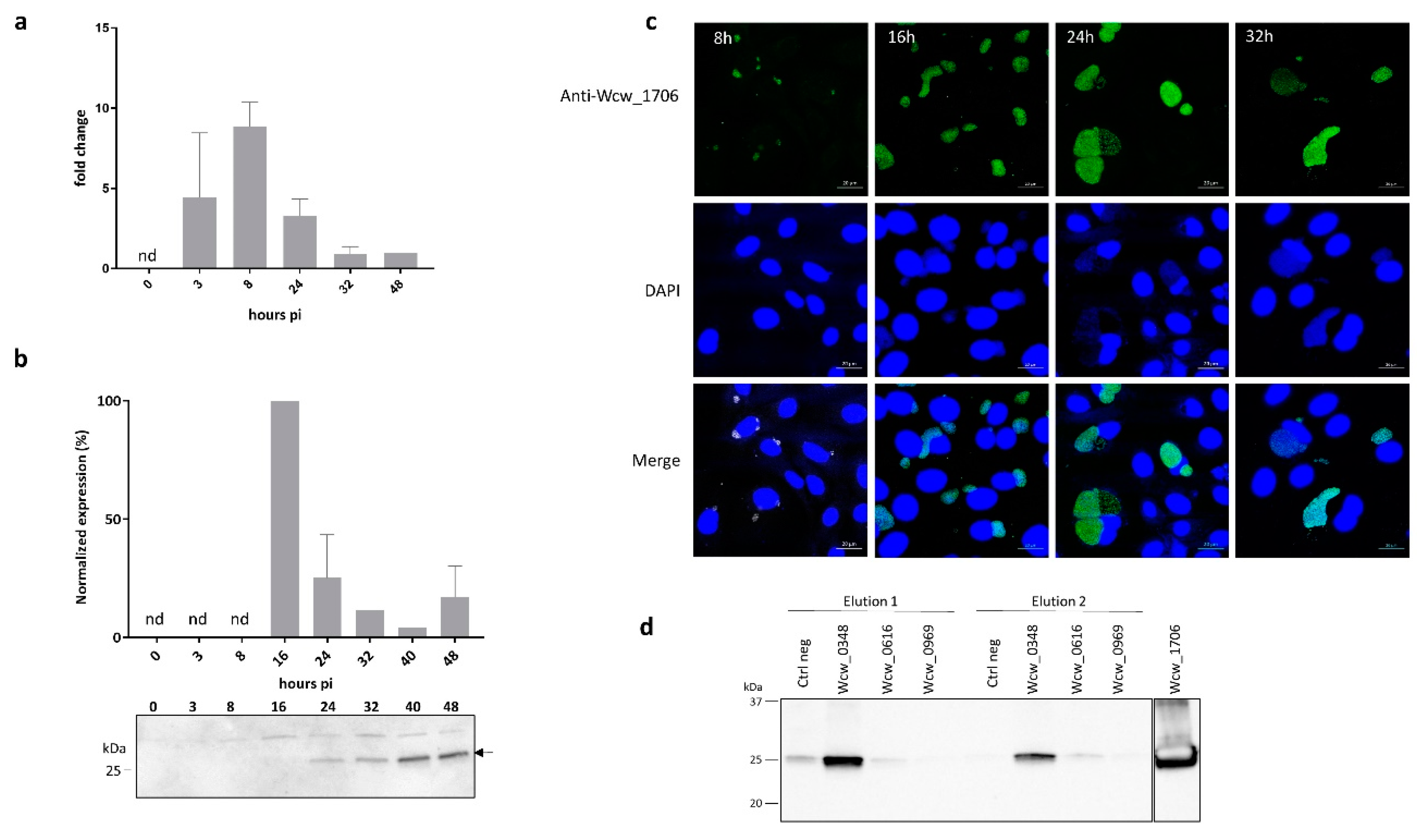
3.5. Wcw_1706 Is a Mid-Cycle Effector Protein Interacting with a T3SS Class II Chaperone
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Data Availability Statement
References
- O’Connell, C.M.; Ferone, M.E. Chlamydia trachomatis Genital Infections. Microb. Cell 2016, 3, 390–403. [Google Scholar] [CrossRef] [PubMed]
- Taylor, H.R.; Burton, M.J.; Haddad, D.; West, S.; Wright, H. Trachoma. Lancet 2014, 384, 2142–2152. [Google Scholar] [CrossRef]
- Baud, D.; Goy, G.; Gerber, S.; Vial, Y.; Hohlfeld, P.; Greub, G. Evidence of maternal-fetal transmission of Parachlamydia acanthamoebae. Emerg. Infect. Dis. 2009, 15, 120–121. [Google Scholar] [CrossRef] [PubMed]
- Baud, D.; Goy, G.; Osterheld, M.C.; Croxatto, A.; Borel, N.; Vial, Y.; Pospischil, A.; Greub, G. Role of Waddlia chondrophila placental infection in miscarriage. Emerg. Infect. Dis. 2014, 20, 460–464. [Google Scholar] [CrossRef]
- Baud, D.; Regan, L.; Greub, G. Emerging role of Chlamydia and Chlamydia-like organisms in adverse pregnancy outcomes. Curr. Opin. Infect. Dis. 2008, 21, 70–76. [Google Scholar] [CrossRef]
- Baud, D.; Thomas, V.; Arafa, A.; Regan, L.; Greub, G. Waddlia chondrophila, a potential agent of human fetal death. Emerg. Infect. Dis. 2007, 13, 1239–1243. [Google Scholar] [CrossRef]
- Hornung, S.; Thuong, B.C.; Gyger, J.; Kebbi-Beghdadi, C.; Vasilevsky, S.; Greub, G.; Baud, D. Role of Chlamydia trachomatis and emerging Chlamydia-related bacteria in ectopic pregnancy in Vietnam. Epidemiol. Infect. 2015, 143, 2635–2638. [Google Scholar] [CrossRef] [Green Version]
- Verweij, S.P.; Kebbi-Beghdadi, C.; Land, J.A.; Ouburg, S.; Morre, S.A.; Greub, G. Waddlia chondrophila and Chlamydia trachomatis antibodies in screening infertile women for tubal pathology. Microbes Infect. 2015, 17, 745–748. [Google Scholar] [CrossRef]
- Goy, G.; Croxatto, A.; Posfay-Barbe, K.M.; Gervaix, A.; Greub, G. Development of a real-time PCR for the specific detection of Waddlia chondrophila in clinical samples. Eur. J. Clin. Microbiol. Infect. Dis. 2009, 28, 1483–1486. [Google Scholar] [CrossRef] [Green Version]
- Haider, S.; Collingro, A.; Walochnik, J.; Wagner, M.; Horn, M. Chlamydia-like bacteria in respiratory samples of community-acquired pneumonia patients. Fems Microbiol Lett 2008, 281, 198–202. [Google Scholar] [CrossRef]
- Barkallah, M.; Gharbi, Y.; Hassena, A.B.; Slima, A.B.; Mallek, Z.; Gautier, M.; Greub, G.; Gdoura, R.; Fendri, I. Survey of infectious etiologies of bovine abortion during mid- to late gestation in dairy herds. PLoS ONE 2014, 9, e91549. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barkallah, M.; Fendri, I.; Dhieb, A.; Gharbi, Y.; Greub, G.; Gdoura, R. First detection of Waddlia chondrophila in Africa using SYBR Green real-time PCR on veterinary samples. Vet. Microbiol. 2013, 164, 101–107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blumer, S.; Greub, G.; Waldvogel, A.; Hassig, M.; Thoma, R.; Tschuor, A.; Pospischil, A.; Borel, N. Waddlia, Parachlamydia and Chlamydiaceae in bovine abortion. Vet. Microbiol. 2011, 152, 385–393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Henning, K.; Schares, G.; Granzow, H.; Polster, U.; Hartmann, M.; Hotzel, H.; Sachse, K.; Peters, M.; Rauser, M. Neospora caninum and Waddlia chondrophila strain 2032/99 in a septic stillborn calf. Vet. Microbiol. 2002, 85, 285–292. [Google Scholar] [CrossRef]
- Rurangirwa, F.R.; Dilbeck, P.M.; Crawford, T.B.; McGuire, T.C.; McElwain, T.F. Analysis of the 16S rRNA gene of micro-organism WSU 86-1044 from an aborted bovine foetus reveals that it is a member of the order Chlamydiales: Proposal of Waddliaceae fam. nov., Waddlia chondrophila gen. nov., sp. nov. Int. J. Syst. Bacteriol. 1999, 49 Pt 2, 577–581. [Google Scholar] [CrossRef] [PubMed]
- Croxatto, A.; Greub, G. Early intracellular trafficking of Waddlia chondrophila in human macrophages. Microbiology 2010, 156, 340–355. [Google Scholar] [CrossRef] [Green Version]
- Kebbi-Beghdadi, C.; Cisse, O.; Greub, G. Permissivity of Vero cells, human pneumocytes and human endometrial cells to Waddlia chondrophila. Microbes Infect. 2011, 13, 566–574. [Google Scholar] [CrossRef]
- Pillonel, T.; Bertelli, C.; Greub, G. Environmental Metagenomic Assemblies Reveal Seven New Highly Divergent Chlamydial Lineages and Hallmarks of a Conserved Intracellular Lifestyle. Front Microbiol. 2018, 9. [Google Scholar] [CrossRef]
- Elwell, C.; Mirrashidi, K.; Engel, J. Chlamydia cell biology and pathogenesis. Nat. Rev. Microbiol. 2016, 14, 385–400. [Google Scholar] [CrossRef]
- Bugalhao, J.N.; Mota, L.J. The multiple functions of the numerous Chlamydia trachomatis secreted proteins: The tip of the iceberg. Microb. Cell 2019, 6, 414–449. [Google Scholar] [CrossRef]
- Beeckman, D.S.; Vanrompay, D.C. Bacterial secretion systems with an emphasis on the chlamydial Type III secretion system. Curr. Issues Mol. Biol. 2010, 12, 17–41. [Google Scholar] [PubMed]
- Coulon, C.; Eterpi, M.; Greub, G.; Collignon, A.; McDonnell, G.; Thomas, V. Amoebal host range, host-free survival and disinfection susceptibility of environmental Chlamydiae as compared to Chlamydia trachomatis. Fems. Immunol. Med. Microbiol. 2012, 64, 364–373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elwell, C.; Engel, J.N. Drosophila melanogaster S2 cells: A model system to study Chlamydia interaction with host cells. Cell Microbiol. 2005, 7, 725–739. [Google Scholar] [CrossRef] [Green Version]
- Kebbi-Beghdadi, C.; Batista, C.; Greub, G. Permissivity of fish cell lines to three Chlamydia-related bacteria: Waddlia chondrophila, Estrella lausannensis and Parachlamydia acanthamoebae. Fems. Immunol. Med. Microbiol. 2011, 63, 339–345. [Google Scholar] [CrossRef] [Green Version]
- Kebbi-Beghdadi, C.; Fatton, M.; Greub, G. Permissivity of insect cells to Waddlia chondrophila, Estrella lausannensis and Parachlamydia acanthamoebae. Microbes Infect. 2015, 17, 749–754. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hackstadt, T.; Rockey, D.D.; Heinzen, R.A.; Scidmore, M.A. Chlamydia trachomatis interrupts an exocytic pathway to acquire endogenously synthesized sphingomyelin in transit from the Golgi apparatus to the plasma membrane. Embo. J. 1996, 15, 964–977. [Google Scholar] [CrossRef] [PubMed]
- Heuer, D.; Rejman Lipinski, A.; Machuy, N.; Karlas, A.; Wehrens, A.; Siedler, F.; Brinkmann, V.; Meyer, T.F. Chlamydia causes fragmentation of the Golgi compartment to ensure reproduction. Nature 2009, 457, 731–735. [Google Scholar] [CrossRef]
- Dumoux, M.; Clare, D.K.; Saibil, H.R.; Hayward, R.D. Chlamydiae assemble a pathogen synapse to hijack the host endoplasmic reticulum. Traffic 2012, 13, 1612–1627. [Google Scholar] [CrossRef] [Green Version]
- Jacquier, N.; Aeby, S.; Lienard, J.; Greub, G. Discovery of new intracellular pathogens by amoebal coculture and amoebal enrichment approaches. J. Vis. Exp. 2013, e51055. [Google Scholar] [CrossRef] [Green Version]
- Glomski, I.J.; Gedde, M.M.; Tsang, A.W.; Swanson, J.A.; Portnoy, D.A. The Listeria monocytogenes hemolysin has an acidic pH optimum to compartmentalize activity and prevent damage to infected host cells. J. Cell Biol. 2002, 156, 1029–1038. [Google Scholar] [CrossRef]
- Kleba, B.; Stephens, R.S. Chlamydial effector proteins localized to the host cell cytoplasmic compartment. Infect. Immun. 2008, 76, 4842–4850. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- da Cunha, M.; Milho, C.; Almeida, F.; Pais, S.V.; Borges, V.; Mauricio, R.; Borrego, M.J.; Gomes, J.P.; Mota, L.J. Identification of type III secretion substrates of Chlamydia trachomatis using Yersinia enterocolitica as a heterologous system. BMC Microbiol. 2014, 14, 40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kebbi-Beghdadi, C.; Pilloux, L.; Croxatto, A.; Tosetti, N.; Pillonel, T.; Greub, G. A predation assay using amoebae to screen for virulence factors unearthed the first W. chondrophila inclusion membrane protein. Sci. Rep. 2019, 9, 19485. [Google Scholar] [CrossRef] [PubMed]
- Radhakrishnan, S.K.; Pritchard, S.; Viollier, P.H. Coupling prokaryotic cell fate and division control with a bifunctional and oscillating oxidoreductase homolog. Dev. Cell 2010, 18, 90–101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pillonel, T.; Tagini, F.; Bertelli, C.; Greub, G. ChlamDB: A comparative genomics database of the phylum Chlamydiae and other members of the Planctomycetes-Verrucomicrobiae-Chlamydiae superphylum. Nucleic. Acids Res. 2019. [Google Scholar] [CrossRef] [PubMed]
- Kebbi-Beghdadi, C.; Domrose, A.; Becker, E.; Cisse, O.H.; Hegemann, J.H.; Greub, G. OmpA family proteins and Pmp-like autotransporter: New adhesins of Waddlia chondrophila. Pathog. Dis. 2015, 73. [Google Scholar] [CrossRef] [Green Version]
- Chomczynski, P.; Mackey, K. Short technical reports. Modification of the TRI reagent procedure for isolation of RNA from polysaccharide- and proteoglycan-rich sources. BioTechniques 1995, 19, 942–945. [Google Scholar] [PubMed]
- Kebbi-Beghdadi, C.; Lienard, J.; Uyttebroeck, F.; Baud, D.; Riederer, B.M.; Greub, G. Identification of immunogenic proteins of Waddlia chondrophila. PLoS ONE 2012, 7, e28605. [Google Scholar] [CrossRef] [Green Version]
- Lienard, J.; Croxatto, A.; Gervaix, A.; Posfay-Barbe, K.; Baud, D.; Kebbi Beghdadi, C.; Greub, G. Undressing of Waddlia chondrophila to enrich its outer membrane proteins to develop a new species-specific ELISA. New Microbe. New Infect. 2014, 2, 13–24. [Google Scholar] [CrossRef]
- Chen, D.; Lei, L.; Lu, C.; Flores, R.; DeLisa, M.P.; Roberts, T.C.; Romesberg, F.E.; Zhong, G. Secretion of the chlamydial virulence factor CPAF requires the Sec-dependent pathway. Microbiology 2010, 156, 3031–3040. [Google Scholar] [CrossRef] [Green Version]
- Shaw, A.C.; Vandahl, B.B.; Larsen, M.R.; Roepstorff, P.; Gevaert, K.; Vandekerckhove, J.; Christiansen, G.; Birkelund, S. Characterization of a secreted Chlamydia protease. Cell Microbiol. 2002, 4, 411–424. [Google Scholar] [CrossRef] [PubMed]
- Snavely, E.A.; Kokes, M.; Dunn, J.D.; Saka, H.A.; Nguyen, B.D.; Bastidas, R.J.; McCafferty, D.G.; Valdivia, R.H. Reassessing the role of the secreted protease CPAF in Chlamydia trachomatis infection through genetic approaches. Pathog. Dis. 2014, 71, 336–351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhong, G.; Fan, P.; Ji, H.; Dong, F.; Huang, Y. Identification of a chlamydial protease-like activity factor responsible for the degradation of host transcription factors. J. Exp. Med. 2001, 193, 935–942. [Google Scholar] [CrossRef] [PubMed]
- Dickinson, M.S.; Anderson, L.N.; Webb-Robertson, B.M.; Hansen, J.R.; Smith, R.D.; Wright, A.T.; Hybiske, K. Proximity-dependent proteomics of the Chlamydia trachomatis inclusion membrane reveals functional interactions with endoplasmic reticulum exit sites. PLoS Pathog. 2019, 15, e1007698. [Google Scholar] [CrossRef] [Green Version]
- Stenner-Liewen, F.; Liewen, H.; Zapata, J.M.; Pawlowski, K.; Godzik, A.; Reed, J.C. CADD, a Chlamydia protein that interacts with death receptors. J. Biol. Chem. 2002, 277, 9633–9636. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhong, G. Chlamydia trachomatis secretion of proteases for manipulating host signaling pathways. Front Microbiol. 2011, 2, 14. [Google Scholar] [CrossRef] [Green Version]
- Cruz-Fisher, M.I.; Cheng, C.; Sun, G.; Pal, S.; Teng, A.; Molina, D.M.; Kayala, M.A.; Vigil, A.; Baldi, P.; Felgner, P.L.; et al. Identification of immunodominant antigens by probing a whole Chlamydia trachomatis open reading frame proteome microarray using sera from immunized mice. Infect. Immun. 2011, 79, 246–257. [Google Scholar] [CrossRef] [Green Version]
- Bastidas, R.J.; Valdivia, R.H. Emancipating Chlamydia: Advances in the Genetic Manipulation of a Recalcitrant Intracellular Pathogen. Microbiol. Mol. Biol. Rev. 2016, 80, 411–427. [Google Scholar] [CrossRef] [Green Version]
- Qi, M.; Gong, S.; Lei, L.; Liu, Q.; Zhong, G. A Chlamydia trachomatis OmcB C-terminal fragment is released into host cell cytoplasm and is immunogenic in humans. Infect. Immun. 2011. [Google Scholar] [CrossRef] [Green Version]
- Jehl, M.A.; Arnold, R.; Rattei, T. Effective--a database of predicted secreted bacterial proteins. Nucleic Acids Res. 2011, 39, D591–D595. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Sun, M.; Bao, H.; White, A.P. T3_MM: A Markov model effectively classifies bacterial type III secretion signals. PLoS ONE 2013, 8, e58173. [Google Scholar] [CrossRef] [PubMed]
- Carpenter, V.; Chen, Y.S.; Dolat, L.; Valdivia, R.H. The Effector TepP Mediates Recruitment and Activation of Phosphoinositide 3-Kinase on Early Chlamydia trachomatis Vacuoles. mSphere 2017, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.S.; Bastidas, R.J.; Saka, H.A.; Carpenter, V.K.; Richards, K.L.; Plano, G.V.; Valdivia, R.H. The Chlamydia trachomatis type III secretion chaperone Slc1 engages multiple early effectors, including TepP, a tyrosine-phosphorylated protein required for the recruitment of CrkI-II to nascent inclusions and innate immune signaling. PLoS Pathog. 2014, 10, e1003954. [Google Scholar] [CrossRef] [PubMed]
- Agrain, C.; Sorg, I.; Paroz, C.; Cornelis, G.R. Secretion of YscP from Yersinia enterocolitica is essential to control the length of the injectisome needle but not to change the type III secretion substrate specificity. Mol. Microbiol. 2005, 57, 1415–1427. [Google Scholar] [CrossRef] [PubMed]
- de Barsy, M.; Herrgott, L.; Martin, V.; Pillonel, T.; Viollier, P.H.; Greub, G. Identification of new DNA-associated proteins from Waddlia chondrophila. Sci. Rep. 2019, 9, 4885. [Google Scholar] [CrossRef] [PubMed]
- Pennini, M.E.; Perrinet, S.; Dautry-Varsat, A.; Subtil, A. Histone methylation by NUE, a novel nuclear effector of the intracellular pathogen Chlamydia trachomatis. PLOS Pathog. 2010, 6, e1000995. [Google Scholar] [CrossRef]
- Brinkworth, A.J.; Malcolm, D.S.; Pedrosa, A.T.; Roguska, K.; Shahbazian, S.; Graham, J.E.; Hayward, R.D.; Carabeo, R.A. Chlamydia trachomatis Slc1 is a type III secretion chaperone that enhances the translocation of its invasion effector substrate TARP. Mol. Microbiol. 2011, 82, 131–144. [Google Scholar] [CrossRef] [Green Version]
- Pallen, M.J.; Francis, M.S.; Futterer, K. Tetratricopeptide-like repeats in type-III-secretion chaperones and regulators. FEMS Microbiol. Lett. 2003, 223, 53–60. [Google Scholar] [CrossRef] [Green Version]
- Fields, K.A.; Hackstadt, T. Evidence for the secretion of Chlamydia trachomatis CopN by a type III secretion mechanism. Mol. Microbiol. 2000, 38, 1048–1060. [Google Scholar] [CrossRef]
- Gong, S.; Lei, L.; Chang, X.; Belland, R.; Zhong, G. Chlamydia trachomatis secretion of hypothetical protein CT622 into host cell cytoplasm via a secretion pathway that can be inhibited by the type III secretion system inhibitor compound 1. Microbiology 2011, 157, 1134–1144. [Google Scholar] [CrossRef] [Green Version]
- Lei, L.; Qi, M.; Budrys, N.; Schenken, R.; Zhong, G. Localization of Chlamydia trachomatis hypothetical protein CT311 in host cell cytoplasm. Microb. Pathog. 2011, 51, 101–109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Z.; Chen, D.; Zhong, Y.; Wang, S.; Zhong, G. The chlamydial plasmid-encoded protein pgp3 is secreted into the cytosol of Chlamydia-infected cells. Infect. Immun. 2008, 76, 3415–3428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, C.; Lei, L.; Peng, B.; Tang, L.; Ding, H.; Gong, S.; Li, Z.; Wu, Y.; Zhong, G. Chlamydia trachomatis GlgA is secreted into host cell cytoplasm. PLoS ONE 2013, 8, e68764. [Google Scholar] [CrossRef] [PubMed]
- Muschiol, S.; Boncompain, G.; Vromman, F.; Dehoux, P.; Normark, S.; Henriques-Normark, B.; Subtil, A. Identification of a family of effectors secreted by the type III secretion system that are conserved in pathogenic Chlamydiae. Infect. Immun. 2011, 79, 571–580. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, X.; Lei, L.; Gong, S.; Chen, D.; Flores, R.; Zhong, G. The chlamydial periplasmic stress response serine protease cHtrA is secreted into host cell cytosol. BMC Microbiol. 2011, 11, 87. [Google Scholar] [CrossRef] [Green Version]
- Kuehn, M.J.; Kesty, N.C. Bacterial outer membrane vesicles and the host-pathogen interaction. Genes Dev. 2005, 19, 2645–2655. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walton, T.A.; Sousa, M.C. Crystal structure of Skp, a prefoldin-like chaperone that protects soluble and membrane proteins from aggregation. Mol. Cell 2004, 15, 367–374. [Google Scholar] [CrossRef] [PubMed]
- Olsen, A.W.; Follmann, F.; Hojrup, P.; Leah, R.; Sand, C.; Andersen, P.; Theisen, M. Identification of human T cell targets recognized during Chlamydia trachomatis genital infection. J. Infect. Dis. 2007, 196, 1546–1552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patton, D.L.; Teng, A.; Randall, A.; Liang, X.; Felgner, P.L.; de la Maza, L.M. Whole genome identification of C. trachomatis immunodominant antigens after genital tract infections and effect of antibiotic treatment of pigtailed macaques. J. Proteom. 2014, 108, 99–109. [Google Scholar] [CrossRef] [Green Version]
- Teng, A.; Cruz-Fisher, M.I.; Cheng, C.; Pal, S.; Sun, G.; Ralli-Jain, P.; Molina, D.M.; Felgner, P.L.; Liang, X.; de la Maza, L.M. Proteomic identification of immunodominant chlamydial antigens in a mouse model. J. Proteom. 2012, 77, 176–186. [Google Scholar] [CrossRef] [Green Version]
- Bertelli, C.; Collyn, F.; Croxatto, A.; Ruckert, C.; Polkinghorne, A.; Kebbi-Beghdadi, C.; Goesmann, A.; Vaughan, L.; Greub, G. The Waddlia genome: A window into chlamydial biology. PLoS ONE 2010, 5, e10890. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Griffiths, E.; Ventresca, M.S.; Gupta, R.S. BLAST screening of chlamydial genomes to identify signature proteins that are unique for the Chlamydiales, Chlamydiaceae, Chlamydophila and Chlamydia groups of species. BMC Genom. 2006, 7, 14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Subtil, A.; Delevoye, C.; Balana, M.E.; Tastevin, L.; Perrinet, S.; Dautry-Varsat, A. A directed screen for chlamydial proteins secreted by a type III mechanism identifies a translocated protein and numerous other new candidates. Mol. Microbiol. 2005, 56, 1636–1647. [Google Scholar] [CrossRef] [PubMed]
- Dill, B.D.; Dessus-Babus, S.; Raulston, J.E. Identification of iron-responsive proteins expressed by Chlamydia trachomatis reticulate bodies during intracellular growth. Microbiology 2009, 155, 210–219. [Google Scholar] [CrossRef] [Green Version]
- Belland, R.J.; Nelson, D.E.; Virok, D.; Crane, D.D.; Hogan, D.; Sturdevant, D.; Beatty, W.L.; Caldwell, H.D. Transcriptome analysis of chlamydial growth during IFN-gamma-mediated persistence and reactivation. Proc. Natl. Acad. Sci. USA 2003, 100, 15971–15976. [Google Scholar] [CrossRef] [Green Version]
- Brinkworth, A.J.; Wildung, M.R.; Carabeo, R.A. Genomewide Transcriptional Responses of Iron-Starved Chlamydia trachomatis Reveal Prioritization of Metabolic Precursor Synthesis over Protein Translation. mSystems 2018, 3. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Number | Identified Proteins | Gene | Molecular | Signal | T3SS Effectors | Previously Described |
|---|---|---|---|---|---|---|
| Name | Weight | Peptide | Prediction 1 | as | ||
| C. trachomatis | ||||||
| CT_610 | PqqC-like protein (CADD) | 27 kDa | NO | 0 | secreted [45] | |
| CT_460 | SWIB (YM74) complex protein | 10 kDa | NO | 1 | nd | |
| CT_112 | Oligoendopeptidase | pepF | 69 kDa | NO | 1 | nd |
| CT_539 | Thioredoxin | trxA | 11 kDa | NO | 1 | not secreted [47] |
| CT_771 | Hydrolase/phosphatase homolog | 17 kDa | NO | 0 | nd | |
| CT_242 | Skp-like protein | 19 kDa | YES | 1 | nd | |
| CT_707 | Trigger factor | tig | 50 kDa | NO | 2 | nd |
| CT_432 | Serine hydroxymethyltransferase | glyA | 54 kDa | NO | 1 | nd |
| CT_691 | Hypothetical protein | 25 kDa | YES | 1 | nd | |
| CT_505 | Glyceraldehyde-3-phosphate dehydrogenase | gapA | 36 kDa | NO | 0 | nd |
| CT_603 | Thio-specific Antioxidant (TSA) Peroxidase | ahpC | 22 kDa | NO | 0 | nd |
| CT_858 | Chlamydial protease-like activity factor | cpaf | 67 kDa | YES | 0 | secreted [42,48] |
| CT_507 | DNA-directed RNA polymerase subunit alpha | rpoA | 42 kDa | NO | 0 | nd |
| W. chondrophila | ||||||
| wcw_0501 | Hypothetical protein | 47 kDa | YES | 3 | nd | |
| wcw_0432 | DO serine protease | htrA3 | 52 kDa | YES | 0 | nd |
| wcw_0499 | Hypothetical protein | 18 kDa | NO | 1 | secreted (this study) | |
| wcw_1545 | Nucleoside diphosphate kinase | ndk2 | 18 kDa | YES | 0 | nd |
| wcw_0991 | Putative chlamydial protease-like activity factor | cpaf | 67 kDa | YES | 0 | nd |
| wcw_1192 | Putative Skp-like protein | ompH | 21 kDa | YES | 1 | nd |
| wcw_1543 | Nucleoside diphosphate kinase | ndk1 | 16 kDa | NO | 0 | nd |
| wcw_0657 | Hypothetical protein | 26 kDa | YES | 0 | nd | |
| wcw_0453 | Putative rhs family protein | rhs11 | 200 kDa | YES | 0 | nd |
| wcw_0967 | Hypothetical protein | 31 kDa | YES | 0 | nd | |
| wcw_0878 | NADPH-dependent FMN reductase | 23 kDa | YES | 0 | nd | |
| wcw_1529 | Peptidyl-prolyl cis-trans isomerase | mip3 | 28 kDa | YES | 0 | nd |
| wcw_0704 | Hypotheticalprotein | 526 kDa | NO | 1 | nd | |
| wcw_1706 | Hypothetical protein | 28 kDa | NO | 1 | secreted (this study) | |
| wcw_0680 | Hypothetical protein | 40 kDa | YES | 0 | nd | |
| wcw_0715 | RNA-binding protein | rbp | 10 kDa | NO | 0 | nd |
| wcw_0969 | Hypothetical protein | 18 kDa | NO | 0 | nd | |
| wcw_1068 | Peptidyl-prolyl cis-trans isomerase | ppiB | 22 kDa | YES | 0 | nd |
| wcw_1301 | Hypothetical protein | 17 kDa | YES | 1 | nd |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kebbi-Beghdadi, C.; Pilloux, L.; Martin, V.; Greub, G. Eukaryotic Cell Permeabilisation to Identify New Putative Chlamydial Type III Secretion System Effectors Secreted within Host Cell Cytoplasm. Microorganisms 2020, 8, 361. https://doi.org/10.3390/microorganisms8030361
Kebbi-Beghdadi C, Pilloux L, Martin V, Greub G. Eukaryotic Cell Permeabilisation to Identify New Putative Chlamydial Type III Secretion System Effectors Secreted within Host Cell Cytoplasm. Microorganisms. 2020; 8(3):361. https://doi.org/10.3390/microorganisms8030361
Chicago/Turabian StyleKebbi-Beghdadi, Carole, Ludovic Pilloux, Virginie Martin, and Gilbert Greub. 2020. "Eukaryotic Cell Permeabilisation to Identify New Putative Chlamydial Type III Secretion System Effectors Secreted within Host Cell Cytoplasm" Microorganisms 8, no. 3: 361. https://doi.org/10.3390/microorganisms8030361
APA StyleKebbi-Beghdadi, C., Pilloux, L., Martin, V., & Greub, G. (2020). Eukaryotic Cell Permeabilisation to Identify New Putative Chlamydial Type III Secretion System Effectors Secreted within Host Cell Cytoplasm. Microorganisms, 8(3), 361. https://doi.org/10.3390/microorganisms8030361