Cryptosporidium parvum gp40/15 Is Associated with the Parasitophorous Vacuole Membrane and Is a Potential Vaccine Target


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Parasite and Cell Lines
2.3. Cpgp40/15 and Cpgp40 Cloning, Expression, and Purification
2.4. Preparation of Cpgp40/15 and Cpgp40 Polyclonal Antibodies
2.5. Western Blot Analysis of Native Cpgp40/15 and Cpgp40
2.6. Indirect Immunofluorescence Microscopy
2.7. Examination of Cpgp40/15 Expression by qPCR
2.8. In Vitro Neutralization of Sporozoite Invasion
3. Results
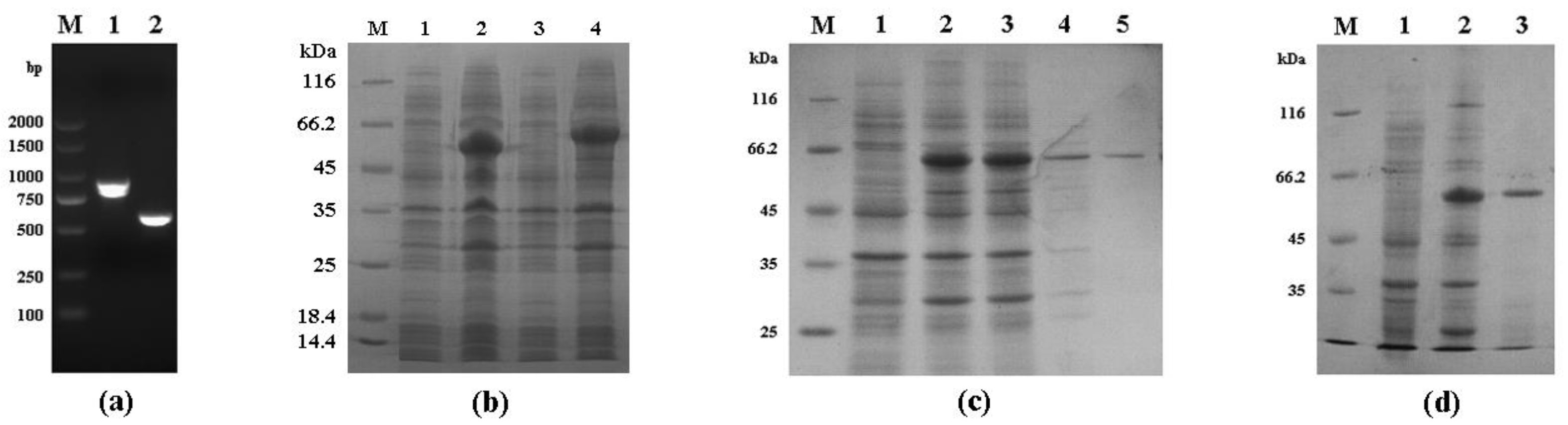
3.1. Expression of Recombinant Cpgp40/15 and Cpgp40
3.2. Identification of Native Cpgp40/15 and Cpgp40
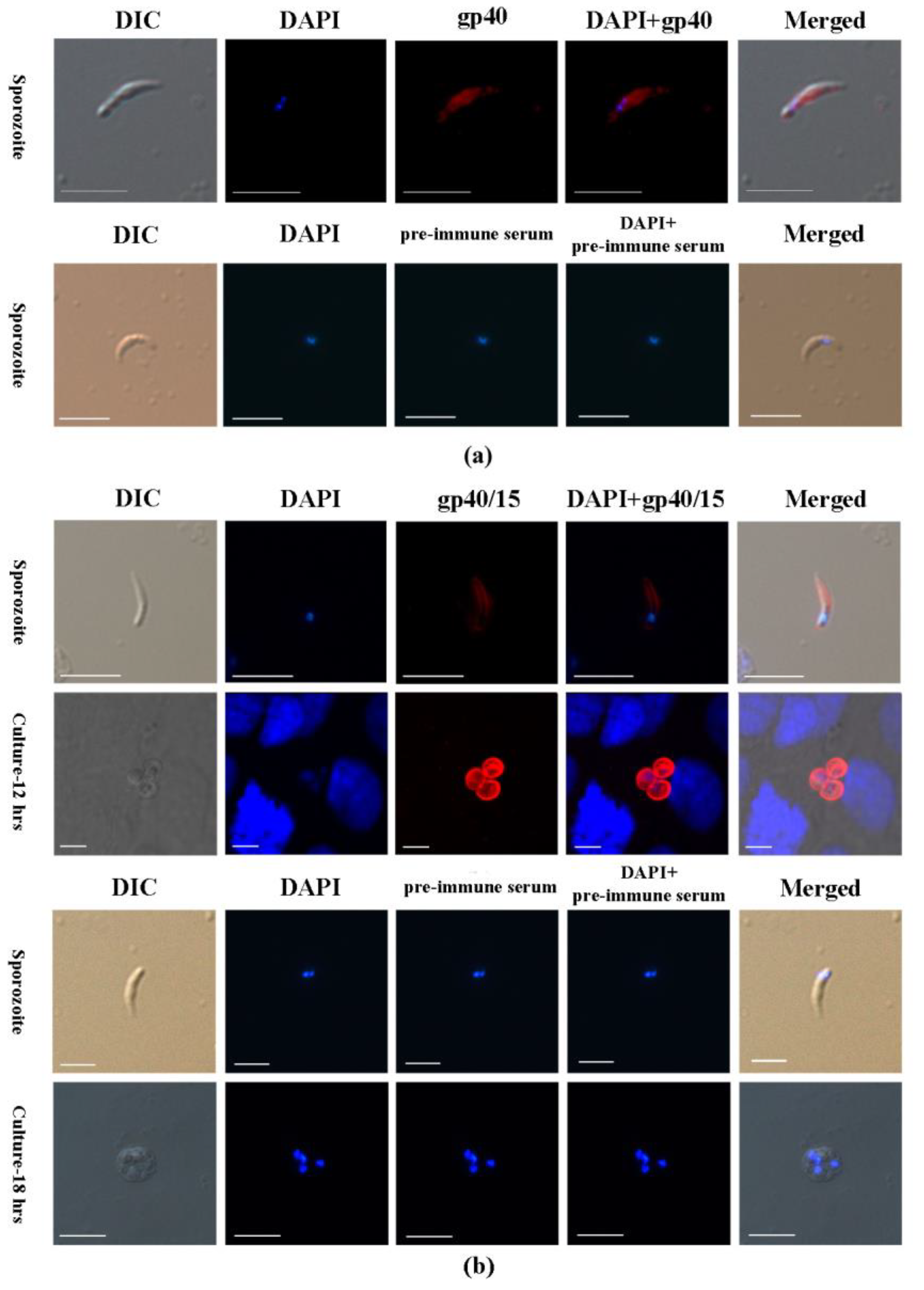
3.3. Cpgp40/15 Is Associated with the PVM during Intracellular Development
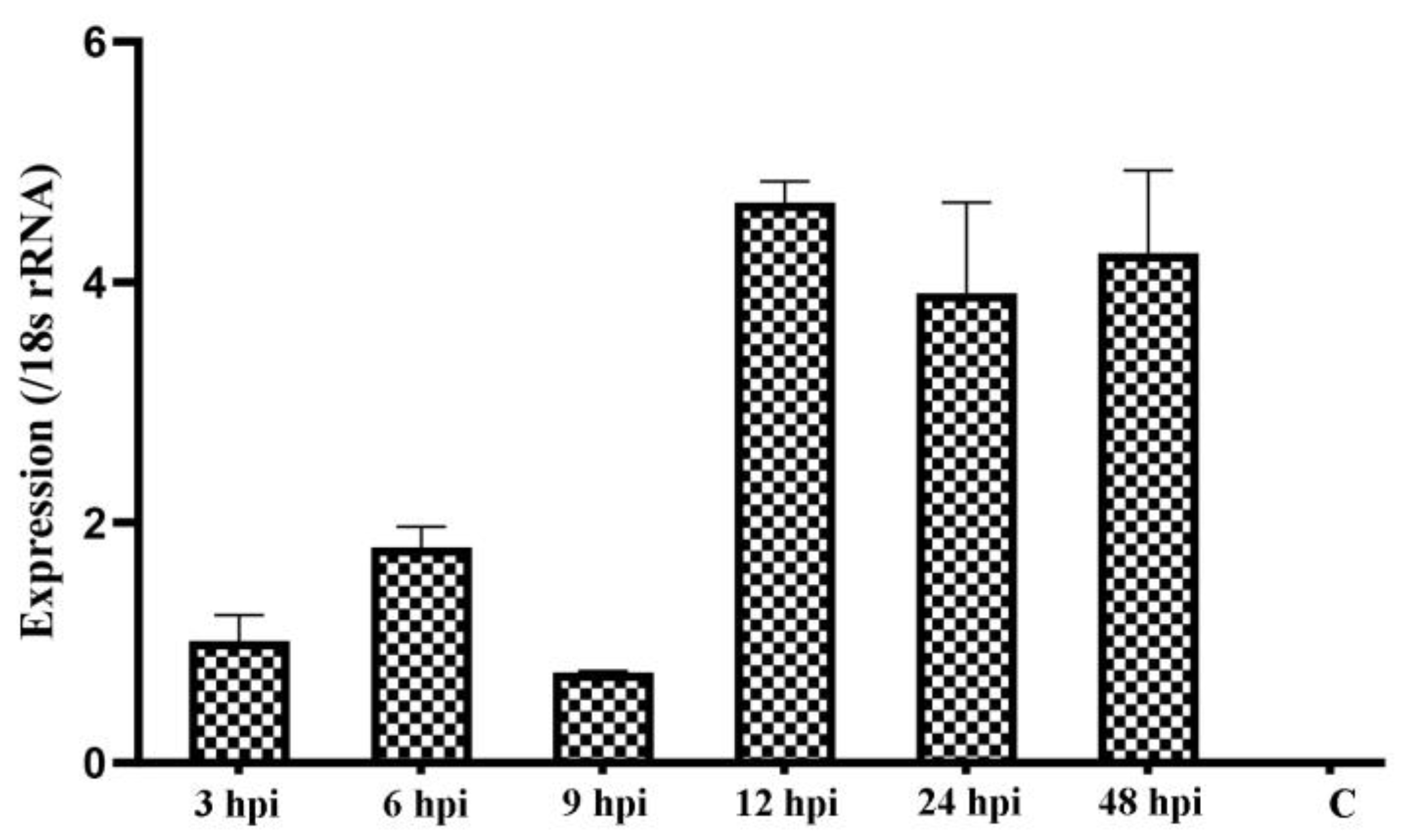
3.4. Cpgp40/15 Is Expressed throughout Intracellular Development
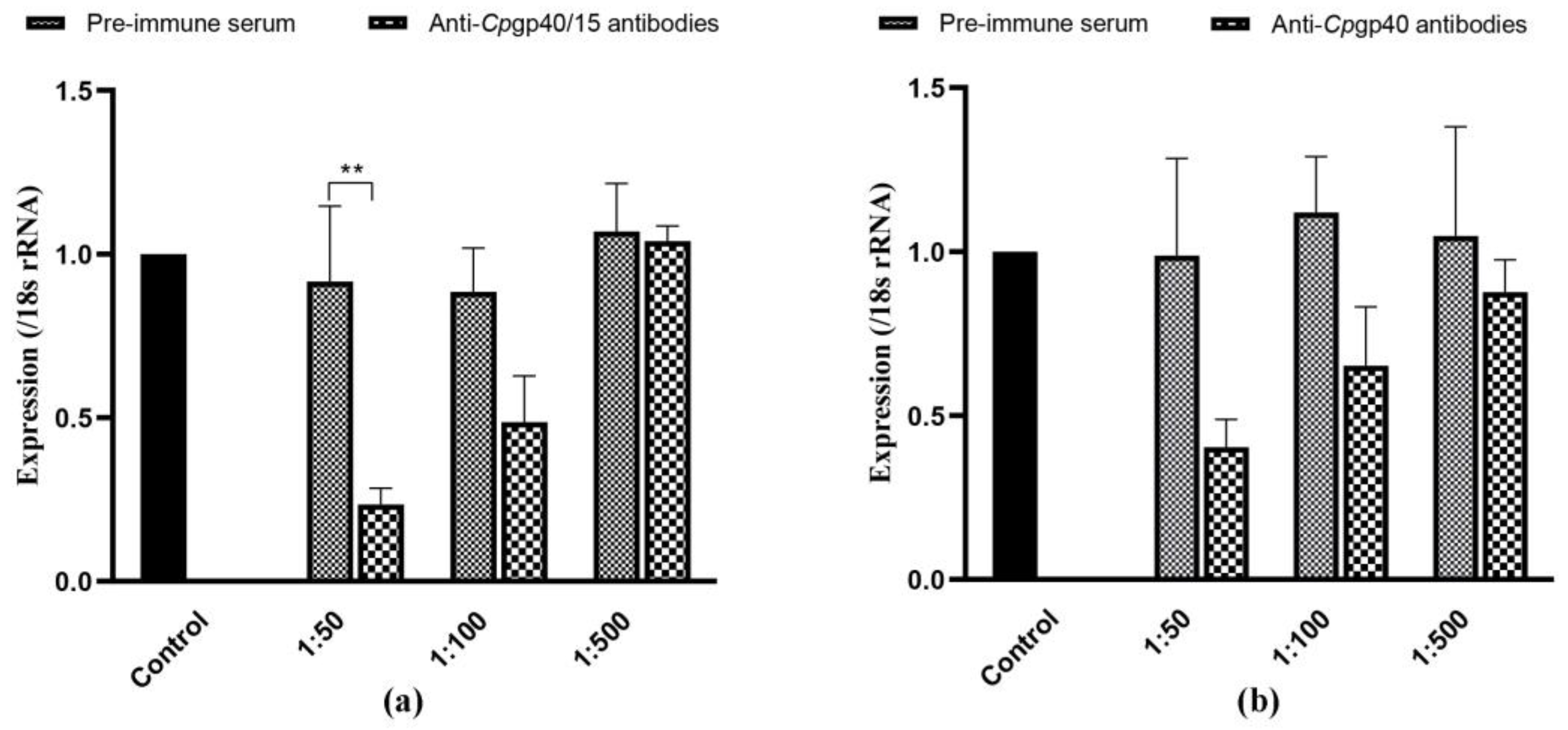
3.5. Cpgp40/15 and Cpgp40 Antibodies Inhibit Cryptosporidium Infection In Vitro
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kotloff, K.L.; Nataro, J.P.; Blackwelder, W.C.; Nasrin, D.; Farag, T.H.; Panchalingam, S.; Wu, Y.; Sow, S.O.; Sur, D.; Breiman, R.F.; et al. Burden and aetiology of diarrhoeal disease in infants and young children in developing countries (the Global Enteric Multicenter Study, GEMS): A prospective, case-control study. Lancet 2013, 382, 209–222. [Google Scholar] [CrossRef]
- O’Connor, R.M.; Shaffie, R.; Kang, G.; Ward, H.D. Cryptosporidiosis in patients with HIV/AIDS. Aids 2011, 25, 549–560. [Google Scholar] [CrossRef] [PubMed]
- Savioli, L.; Smith, H.; Thompson, A. Giardia and Cryptosporidium join the ‘Neglected Diseases Initiative’. Trends Parasitol. 2006, 22, 203–208. [Google Scholar] [CrossRef] [PubMed]
- Kelly, P. Treatment and prevention of cryptosporidiosis: What options are there for a country like Zambia? Parasitology 2011, 138, 1488–1491. [Google Scholar] [CrossRef]
- Gargala, G.; Delaunay, A.; Li, X.; Brasseur, P.; Favennec, L.; Ballet, J.J. Efficacy of nitazoxanide, tizoxanide and tizoxanide glucuronide against Cryptosporidium parvum development in sporozoite-infected HCT-8 enterocytic cells. J. Antimicrob. Chemother. 2000, 46, 57–60. [Google Scholar] [CrossRef]
- Certad, G.; Viscogliosi, E.; Chabé, M.; Cacciò, S.M. Pathogenic Mechanisms of Cryptosporidium and Giardia. Trends Parasitol. 2017, 33, 561–576. [Google Scholar] [CrossRef]
- Xu, R.; Guo, Y.; Li, N.; Zhang, Q.; Wu, H.; Ryan, U.; Feng, Y.; Xiao, L. Characterization of INS-15, A Metalloprotease Potentially Involved in the Invasion of Cryptosporidium parvum. Microorganisms 2019, 7, 452. [Google Scholar] [CrossRef] [Green Version]
- Singh, P.; Mirdha, B.R.; Srinivasan, A.; Rukmangadachar, L.A.; Singh, S.; Sharma, P.; Hariprasad, G.; Gururao, H.; Luthra, K. Identification of invasion proteins of Cryptosporidium parvum. World J. Microbiol. Biotechnol. 2015, 31, 1923–1934. [Google Scholar] [CrossRef]
- Bouzid, M.; Hunter, P.R.; Chalmers, R.M.; Tyler, K.M. Cryptosporidium pathogenicity and virulence. Clin. Microbiol. Rev. 2013, 26, 115–134. [Google Scholar] [CrossRef] [Green Version]
- Wanyiri, J.; Ward, H. Molecular basis of Cryptosporidium-host cell interactions: Recent advances and future prospects. Future Microbiol. 2006, 1, 201–208. [Google Scholar] [CrossRef]
- Okhuysen, P.C.; Chappell, C.L. Cryptosporidium virulence determinants—Are we there yet? Int. J. Parasitol. 2002, 32, 517–525. [Google Scholar] [CrossRef]
- Zhang, H.; Guo, F.; Zhu, G. Cryptosporidium Lactate Dehydrogenase Is Associated with the Parasitophorous Vacuole Membrane and Is a Potential Target for Developing Therapeutics. PLoS Pathog. 2015, 11, e1005250. [Google Scholar] [CrossRef] [PubMed]
- Strong, W.B.; Gut, J.; Nelson, R.G. Cloning and sequence analysis of a highly polymorphic Cryptosporidium parvum gene encoding a 60-kilodalton glycoprotein and characterization of its 15- and 45-kilodalton zoite surface antigen products. Infect. Immun. 2000, 68, 4117–4134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cevallos, A.M.; Zhang, X.; Waldor, M.K.; Jaison, S.; Zhou, X.; Tzipori, S.; Neutra, M.R.; Ward, H.D. Molecular cloning and expression of a gene encoding Cryptosporidium parvum glycoproteins gp40 and gp15. Infect. Immun. 2000, 68, 4108–4116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Connor, R.M.; Kim, K.; Khan, F.; Ward, H.D. Expression of Cpgp40/15 in Toxoplasma gondii: A surrogate system for the study of Cryptosporidium glycoprotein antigens. Infect. Immun. 2003, 71, 6027. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Winter, G.; Gooley, A.A.; Williams, K.L.; Slade, M.B. Characterization of a major sporozoite surface glycoprotein of Cryptosporidum parvum. Funct. Integr. Genom. 2000, 1, 207–217. [Google Scholar] [CrossRef] [PubMed]
- O’Connor, R.M.; Wanyiri, J.W.; Cevallos, A.M.; Priest, J.W.; Ward, H.D. Cryptosporidium parvum glycoprotein gp40 localizes to the sporozoite surface by association with gp15. Mol. Biochem. Parasitol. 2007, 156, 80–83. [Google Scholar] [CrossRef] [Green Version]
- Xiao, L.; Feng, Y. Molecular epidemiologic tools for waterborne pathogens Cryptosporidium spp. and Giardia duodenalis. Food Waterborne Parasitol. 2017, 8–9, 14–32. [Google Scholar] [CrossRef]
- Cai, X.; Woods, K.M.; Upton, S.J.; Zhu, G. Application of quantitative real-time reverse transcription-PCR in assessing drug efficacy against the intracellular pathogen Cryptosporidium parvum in vitro. Antimicrob. Agents Chemother. 2005, 49, 4437–4442. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Wanyiri, J.W.; O’Connor, R.; Allison, G.; Kim, K.; Kane, A.; Qiu, J.; Plaut, A.G.; Ward, H.D. Proteolytic processing of the Cryptosporidium glycoprotein gp40/15 by human furin and by a parasite-derived furin-like protease activity. Infect. Immun. 2007, 75, 184–192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chatterjee, A.; Banerjee, S.; Steffen, M.; O’Connor, R.M.; Ward, H.D.; Robbins, P.W.; Samuelson, J. Evidence for mucin-like glycoproteins that tether sporozoites of Cryptosporidium parvum to the inner surface of the oocyst wall. Eukaryot. Cell 2010, 9, 84–96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeng, B.; Cai, X.; Zhu, G. Functional characterization of a fatty acyl-CoA-binding protein (ACBP) from the apicomplexan Cryptosporidium parvum. Microbiology 2006, 152, 2355–2363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fritzler, J.M.; Millership, J.J.; Zhu, G. Cryptosporidium parvum long-chain fatty acid elongase. Eukaryot. Cell 2007, 6, 2018–2028. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mauzy, M.; Enomoto, S.; Lancto, C.; Abrahamsen, M.; Rutherford, M. The Cryptosporidium parvum transcriptome during in vitro development. PLoS ONE 2012, 7, e31715. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lippuner, C.; Ramakrishnan, C.; Basso, W.; Schmid, M.; Okoniewski, M.; Smith, N.; Hässig, M.; Deplazes, P.; Hehl, A. RNA-Seq analysis during the life cycle of Cryptosporidium parvum reveals significant differential gene expression between proliferating stages in the intestine and infectious sporozoites. Int. J. Parasitol. 2018, 48, 413–422. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mirhashemi, M.; Noubary, F.; Chapman-Bonofiglio, S.; Tzipori, S.; Huggins, G.; Widmer, G. Transcriptome analysis of pig intestinal cell monolayers infected with Cryptosporidium parvum asexual stages. Parasites Vectors 2018, 11, 176. [Google Scholar] [CrossRef]
- Isaza, J.; Galván, A.; Polanco, V.; Huang, B.; Matveyev, A.; Serrano, M.; Manque, P.; Buck, G.; Alzate, J. Revisiting the reference genomes of human pathogenic Cryptosporidium species: Reannotation of C. parvum Iowa and a new C. hominis reference. Sci. Rep. 2015, 5, 16324. [Google Scholar] [CrossRef]
- Matos, L.; McEvoy, J.; Tzipori, S.; Bresciani, K.; Widmer, G. The transcriptome of Cryptosporidium oocysts and intracellular stages. Sci. Rep. 2019, 9, 7856. [Google Scholar] [CrossRef] [Green Version]
- Tandel, J.; English, E.; Sateriale, A.; Gullicksrud, J.; Beiting, D.; Sullivan, M.; Pinkston, B.; Striepen, B. Life cycle progression and sexual development of the apicomplexan parasite Cryptosporidium parvum. Nat. Microbiol. 2019, 4, 2226–2236. [Google Scholar] [CrossRef] [Green Version]
- Chavez, M.A.; White, A.C., Jr. Novel treatment strategies and drugs in development for cryptosporidiosis. Expert Rev. Anti-Infect. Ther. 2018, 16, 655–661. [Google Scholar] [CrossRef] [PubMed]
- Boulter-Bitzer, J.I.; Lee, H.; Trevors, J.T. Molecular targets for detection and immunotherapy in Cryptosporidium parvum. Biotechnol. Adv. 2007, 25, 13–44. [Google Scholar] [CrossRef] [PubMed]
- Crabb, J.H. Antibody-based immunotherapy of cryptosporidiosis. Adv. Parasitol. 1998, 40, 121–149. [Google Scholar] [CrossRef]
- Avendano, C.; Jenkins, M.; Mendez-Callejas, G.; Oviedo, J.; Guzman, F.; Patarroyo, M.A.; Sanchez-Acedo, C.; Quilez, J. Cryptosporidium spp. CP15 and CSL protein-derived synthetic peptides’ immunogenicity and in vitro seroneutralisation capability. Vaccine 2018, 36, 6703–6710. [Google Scholar] [CrossRef] [PubMed]
- Riggs, M.; Stone, A.; Yount, P.; Langer, R.; Arrowood, M.; Bentley, D. Protective monoclonal antibody defines a circumsporozoite-like glycoprotein exoantigen of Cryptosporidium parvum sporozoites and merozoites. J. Immunol. 1997, 158, 1787–1795. [Google Scholar]
- Manjunatha, U.; Vinayak, S.; Zambriski, J.; Chao, A.; Sy, T.; Noble, C.; Bonamy, G.; Kondreddi, R.; Zou, B.; Gedeck, P. A Cryptosporidium PI(4)K inhibitor is a drug candidate for cryptosporidiosis. Nature 2017, 546, 376–380. [Google Scholar] [CrossRef]
- Arnold, S.; Choi, R.; Hulverson, M.; Schaefer, D.; Vinayak, S.; Vidadala, R.; McCloskey, M.; Whitman, G.; Huang, W.; Barrett, L. Necessity of bumped kinase inhibitor gastrointestinal exposure in treating Cryptosporidium infection. J. Infect. Dis. 2017, 216, 55–63. [Google Scholar] [CrossRef]





© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cui, Z.; Wang, L.; Wang, Y.; Li, J.; Wang, R.; Sun, M.; Zhang, L. Cryptosporidium parvum gp40/15 Is Associated with the Parasitophorous Vacuole Membrane and Is a Potential Vaccine Target. Microorganisms 2020, 8, 363. https://doi.org/10.3390/microorganisms8030363
Cui Z, Wang L, Wang Y, Li J, Wang R, Sun M, Zhang L. Cryptosporidium parvum gp40/15 Is Associated with the Parasitophorous Vacuole Membrane and Is a Potential Vaccine Target. Microorganisms. 2020; 8(3):363. https://doi.org/10.3390/microorganisms8030363
Chicago/Turabian StyleCui, Zhaohui, Luyang Wang, Yuexin Wang, Juan Li, Rongjun Wang, Mingfei Sun, and Longxian Zhang. 2020. "Cryptosporidium parvum gp40/15 Is Associated with the Parasitophorous Vacuole Membrane and Is a Potential Vaccine Target" Microorganisms 8, no. 3: 363. https://doi.org/10.3390/microorganisms8030363
APA StyleCui, Z., Wang, L., Wang, Y., Li, J., Wang, R., Sun, M., & Zhang, L. (2020). Cryptosporidium parvum gp40/15 Is Associated with the Parasitophorous Vacuole Membrane and Is a Potential Vaccine Target. Microorganisms, 8(3), 363. https://doi.org/10.3390/microorganisms8030363