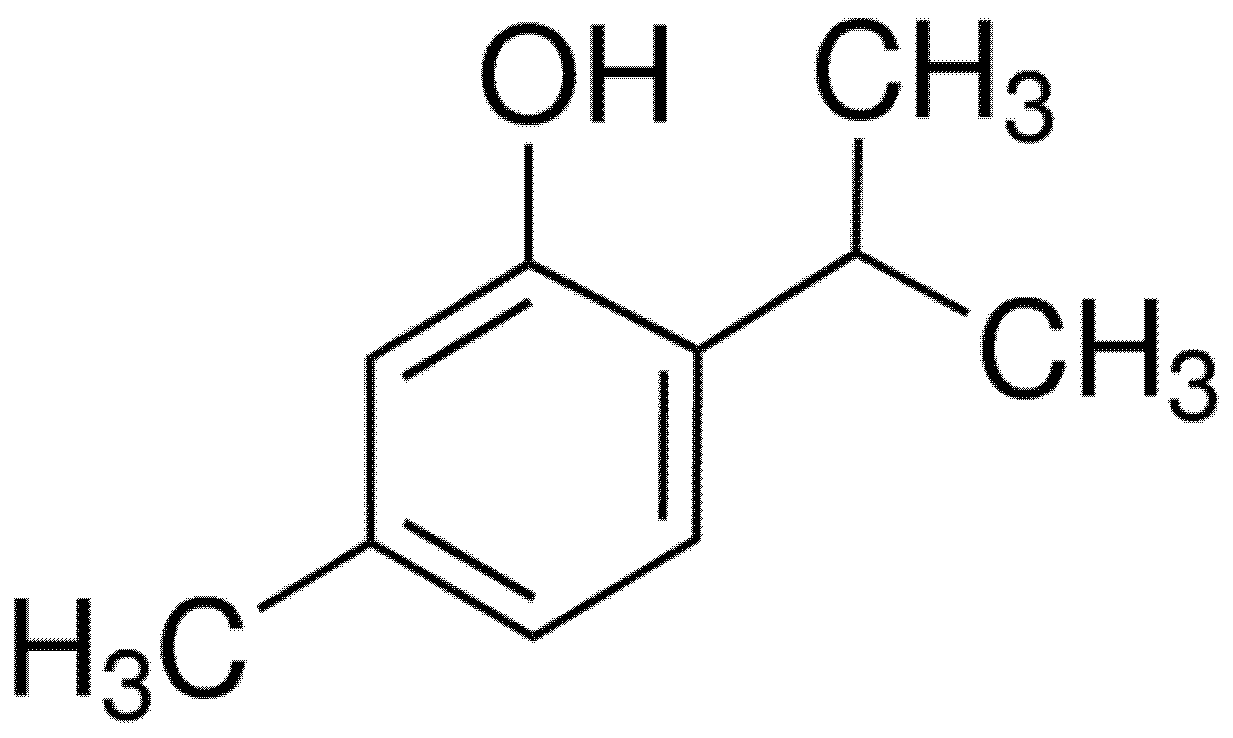
Thymol Protects Channel Catfish from Aeromonas hydrophila Infection by Inhibiting Aerolysin Expression and Biofilm Formation

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Microorganism and Reagents
2.2. Determination of Minimal Inhibitory Concentrations
2.3. Growth Curves
2.4. Hemolytic Activity Assay
2.5. Western Blot Analysis
2.6. Biofilm Formation Assay
2.7. RNA Extraction and Real-Time PCR
2.8. Cell Viability Assays
2.9. Channel Catfish Model of A. hydrophila Infection
2.10. Statistical Analysis
3. Results
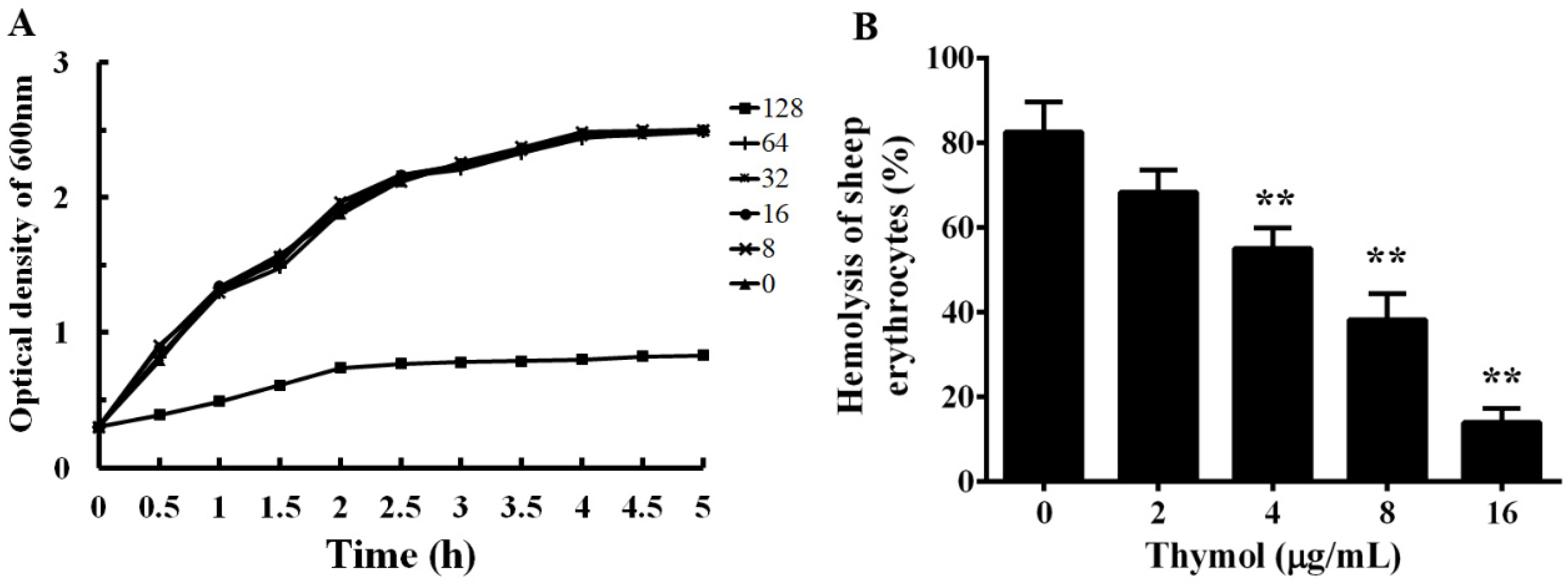
3.1. Thymol has Little Influence on the Growth of A. hydrophila
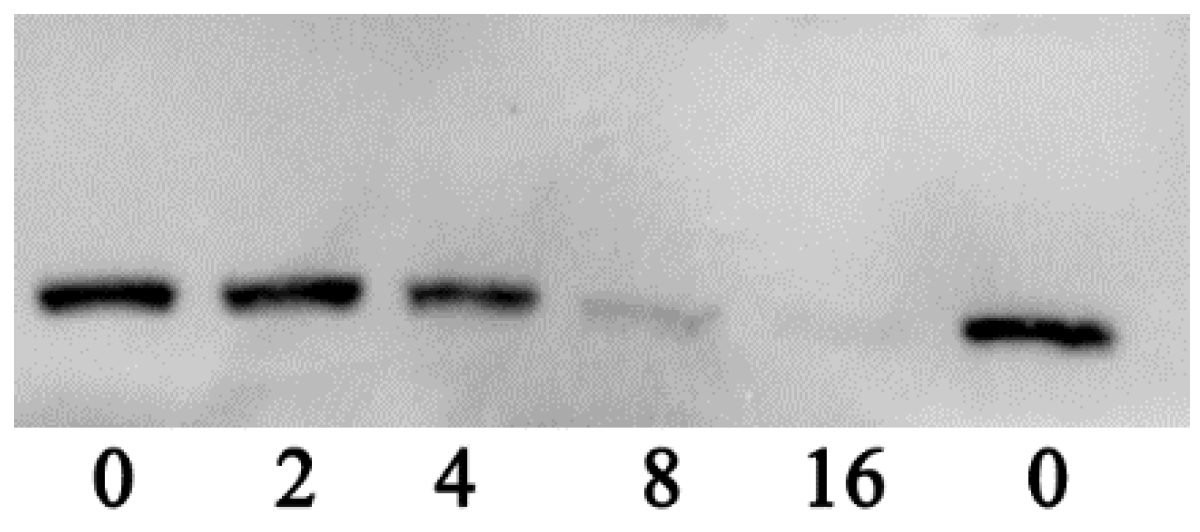
3.2. Thymol Inhibits the Hemolytic Activity by Decreasing the Production of Aerolysin
3.3. Thymol Reduces the Formation of Biofilm
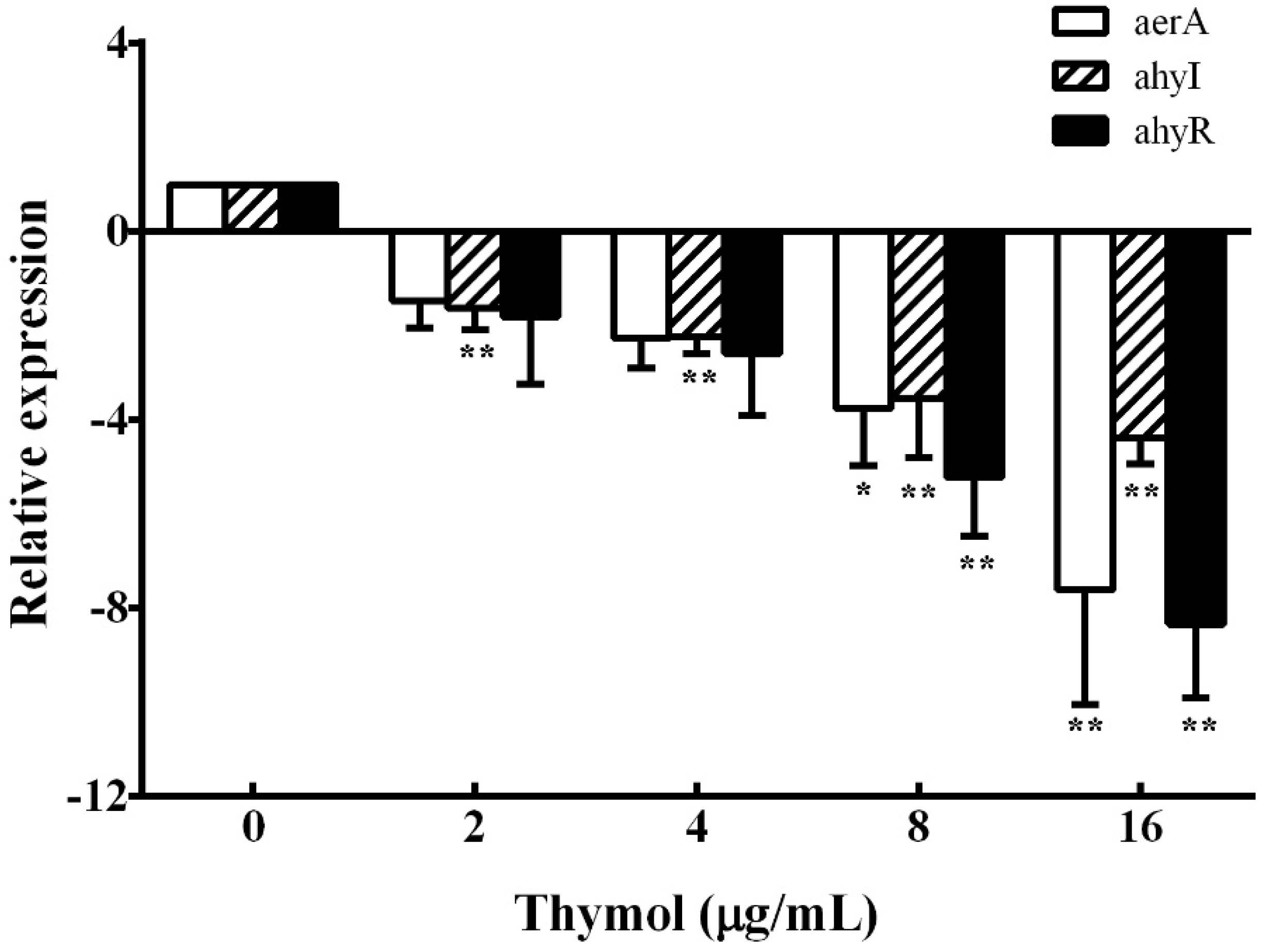
3.4. Real-Time PCR
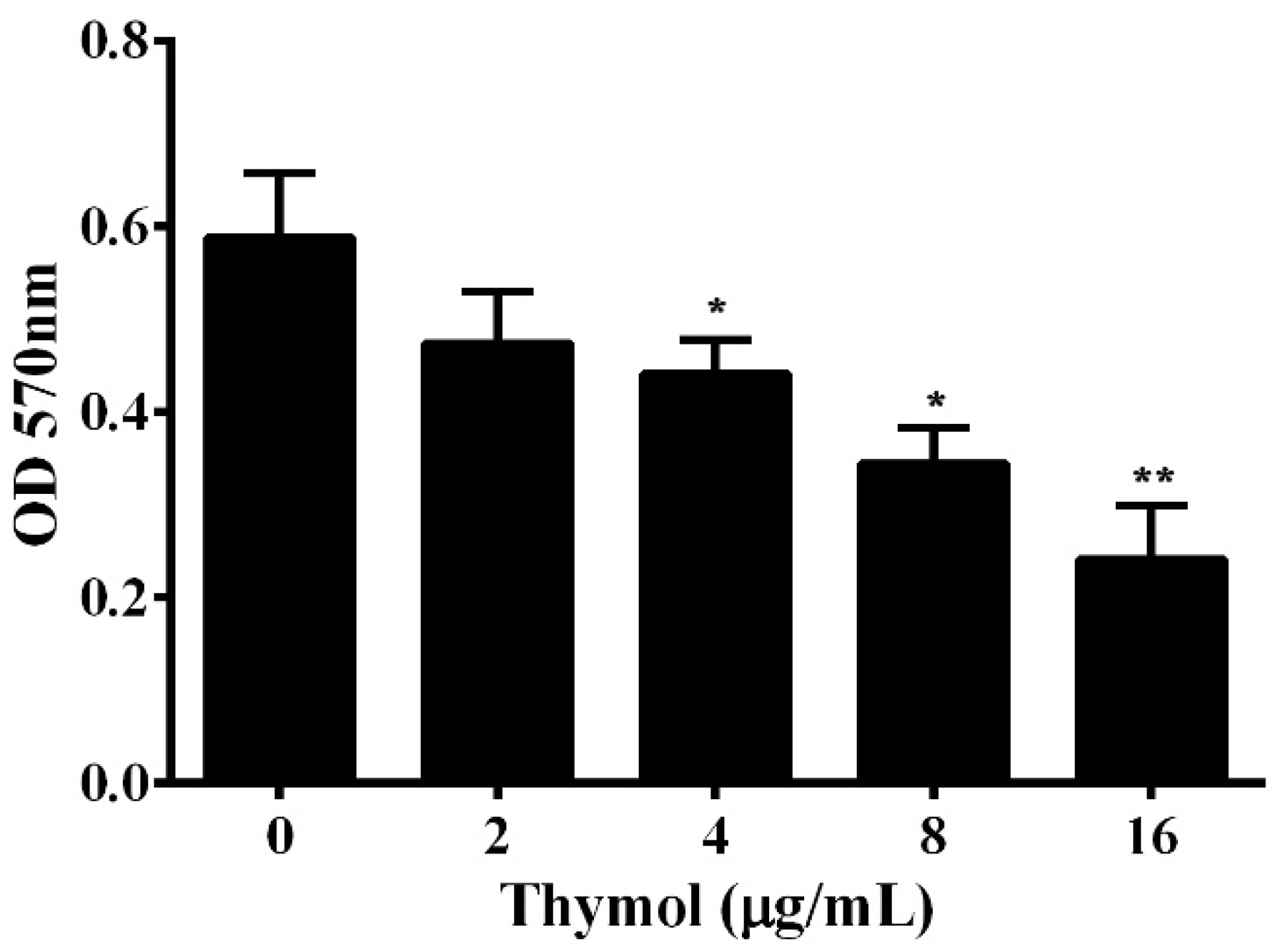
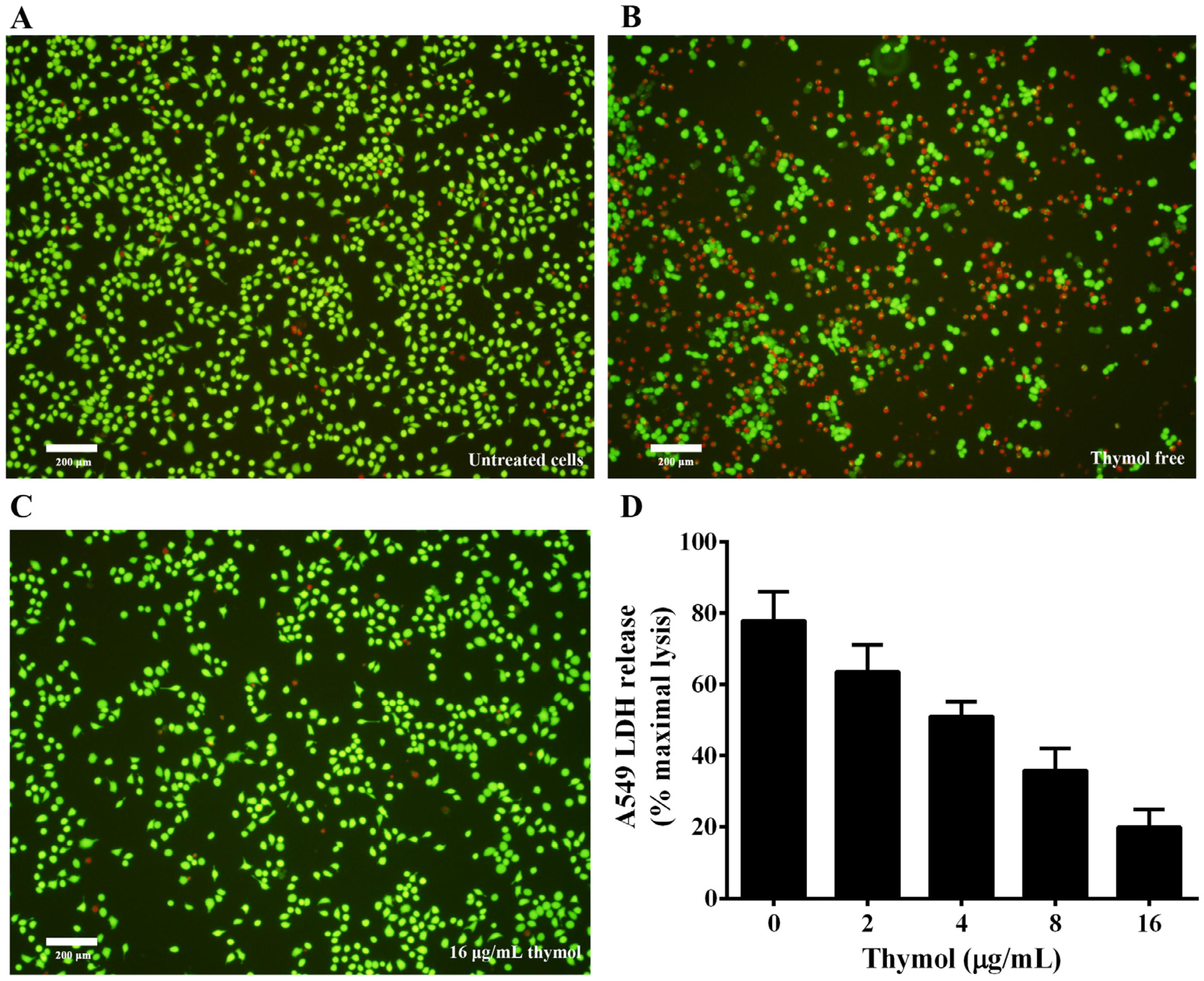
3.5. Thymol Protects A549 Cells from Cell Injury Induced by A. hydrophila
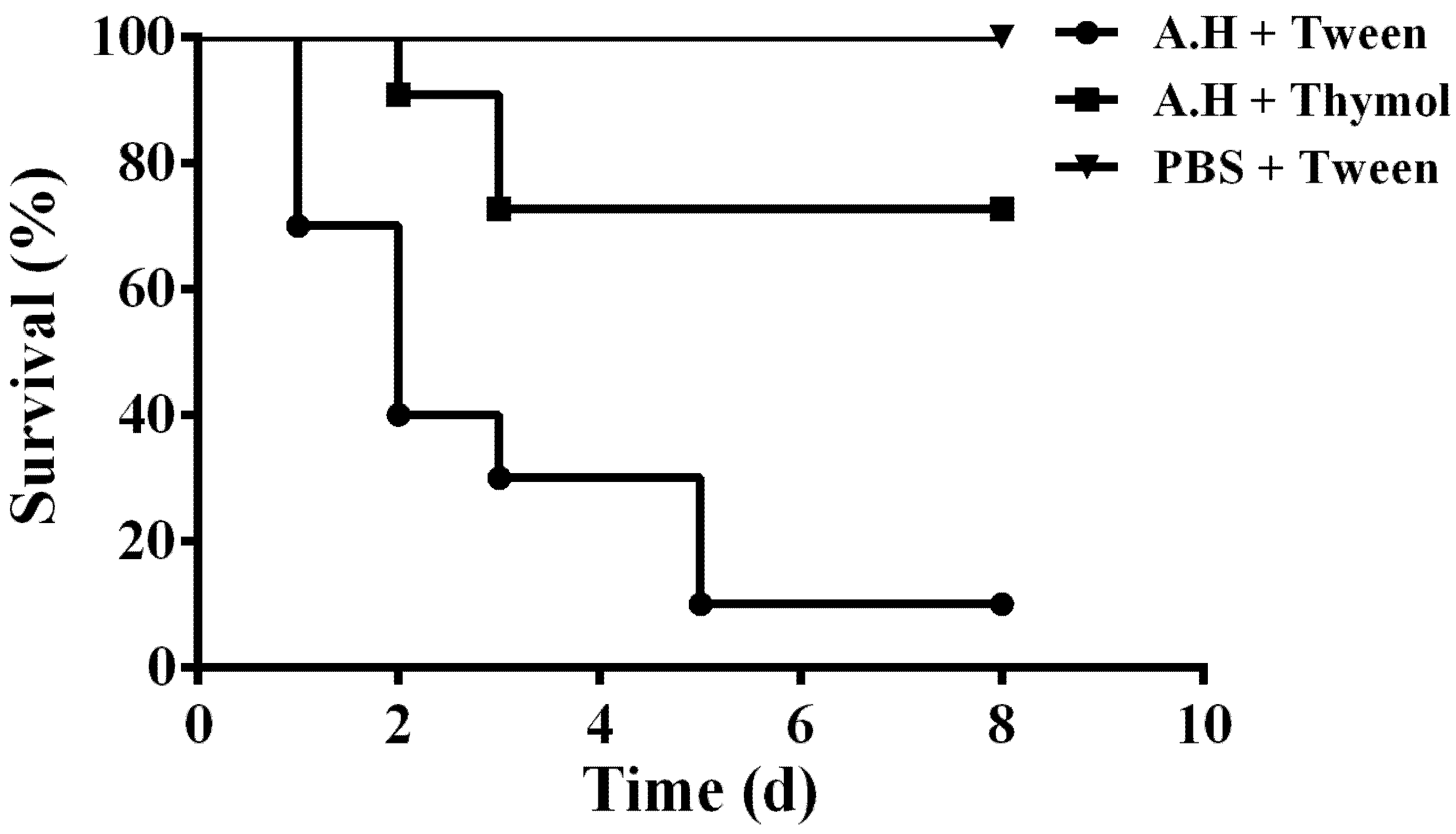
3.6. Thymol Decreases the Mortality of Channel Catfish Infected with A. hydrophila
4. Discussion
Author Contributions
Funding
Conflicts of Interest
References
- El-Bahar, H.M.; Ali, N.G.; Aboyadak, I.M.; Khalil, S.; Ibrahim, M.S. Virulence genes contributing to Aeromonas hydrophila pathogenicity in Oreochromis niloticus. Int. Microbiol. Off. J. Span. Soc. Microbiol. 2019, 22, 479–490. [Google Scholar] [CrossRef] [PubMed]
- Singh, V.; Chaudhary, D.K.; Mani, I.; Jain, R.; Mishra, B.N. Development of diagnostic and vaccine markers through cloning, expression, and regulation of putative virulence-protein-encoding genes of Aeromonas hydrophila. J. Microbiol. 2013, 51, 275–282. [Google Scholar] [CrossRef] [PubMed]
- Dong, J.; Ding, H.; Liu, Y.; Yang, Q.; Xu, N.; Yang, Y.; Ai, X. Magnolol protects channel catfish from Aeromonas hydrophila infection via inhibiting the expression of aerolysin. Vet. Microbiol. 2017, 211, 119–123. [Google Scholar] [CrossRef] [PubMed]
- Yousr, A.H.; Napis, S.; Rahmat Ali, G.R.; Radu, S. Detection of Aerolysin and Hemolysin Genes in Aeromonas spp.Isolated from Environmental and Shellfish Sources byPolymerase Chain Reaction. ASEAN Food J. 2007, 14, 115–122. [Google Scholar]
- Isonhood, J.H.; Drake, M. Aeromonas species in foods. J. Food Prot. 2002, 65, 575–582. [Google Scholar] [CrossRef]
- Wang, Q.; Ji, W.; Xu, Z. Current use and development of fish vaccines in China. Fish Shellfish Immunol. 2020, 96, 223–234. [Google Scholar] [CrossRef]
- Maura, D.; Ballok, A.E.; Rahme, L.G. Considerations and caveats in anti-virulence drug development. Curr. Opin. Microbiol. 2016, 33, 41–46. [Google Scholar] [CrossRef] [Green Version]
- Bucker, R.; Krug, S.M.; Rosenthal, R.; Gunzel, D.; Fromm, A.; Zeitz, M.; Chakraborty, T.; Fromm, M.; Epple, H.J.; Schulzke, J.D. Aerolysin from Aeromonas hydrophila perturbs tight junction integrity and cell lesion repair in intestinal epithelial HT-29/B6 cells. J. Infect. Dis. 2011, 204, 1283–1292. [Google Scholar] [CrossRef] [Green Version]
- Abrami, L.; Fivaz, M.; Decroly, E.; Seidah, N.G.; Jean, F.; Thomas, G.; Leppla, S.H.; Buckley, J.T.; van der Goot, F.G. The pore-forming toxin proaerolysin is activated by furin. J. Biol. Chem. 1998, 273, 32656–32661. [Google Scholar] [CrossRef] [Green Version]
- Howard, S.P.; Buckley, J.T. Membrane glycoprotein receptor and hole-forming properties of a cytolytic protein toxin. Biochemistry 1982, 21, 1662–1667. [Google Scholar] [CrossRef]
- Zhang, D.; Pridgeon, J.W.; Klesius, P.H. Expression and activity of recombinant proaerolysin derived from Aeromonas hydrophila cultured from diseased channel catfish. Vet. Microbiol. 2013, 165, 478–482. [Google Scholar] [CrossRef] [PubMed]
- Chakraborty, T.; Huhle, B.; Hof, H.; Bergbauer, H.; Goebel, W. Marker exchange mutagenesis of the aerolysin determinant in Aeromonas hydrophila demonstrates the role of aerolysin in A. hydrophila-associated systemic infections. Infect. Immun. 1987, 55, 2274–2280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marchese, A.; Orhan, I.E.; Daglia, M.; Barbieri, R.; Di Lorenzo, A.; Nabavi, S.F.; Gortzi, O.; Izadi, M.; Nabavi, S.M. Antibacterial and antifungal activities of thymol: A brief review of the literature. Food Chem. 2016, 210, 402–414. [Google Scholar] [CrossRef] [PubMed]
- Kwon, H.I.; Jeong, N.H.; Kim, S.Y.; Kim, M.H.; Son, J.H.; Jun, S.H.; Kim, S.; Jeon, H.; Kang, S.C.; Kim, S.H.; et al. Inhibitory effects of thymol on the cytotoxicity and inflammatory responses induced by Staphylococcus aureus extracellular vesicles in cultured keratinocytes. Microb. Pathog. 2019, 134, 103603. [Google Scholar] [CrossRef] [PubMed]
- CLSI. Methods for DilutionAntimicrobial Susceptibility Tests for Bacteria That Grow Aerobically-Eighth Edition: Approved Standard M07-A8.; CLSI: Wayne, PA, USA, 2009. [Google Scholar]
- Iacovache, I.; Degiacomi, M.T.; Pernot, L.; Ho, S.; Schiltz, M.; Dal Peraro, M.; van der Goot, F.G. Dual chaperone role of the C-terminal propeptide in folding and oligomerization of the pore-forming toxin aerolysin. PLoS Pathog. 2011, 7, e1002135. [Google Scholar] [CrossRef] [Green Version]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef]
- Patel, B.; Kumari, S.; Banerjee, R.; Samanta, M.; Das, S. Disruption of the quorum sensing regulated pathogenic traits of the biofilm-forming fish pathogen Aeromonas hydrophila by tannic acid, a potent quorum quencher. Biofouling 2017, 33, 580–590. [Google Scholar] [CrossRef]
- Ham, Y.; Kim, T.J. Inhibitory activity of monoacylglycerols on biofilm formation in Aeromonas hydrophila, Streptococcus mutans, Xanthomonas oryzae, and Yersinia enterocolitica. SpringerPlus 2016, 5, 1526. [Google Scholar] [CrossRef] [Green Version]
- Chambers, J.R.; Sauer, K. Small RNAs and their role in biofilm formation. Trends Microbiol. 2013, 21, 39–49. [Google Scholar] [CrossRef] [Green Version]
- Li, W.; Yao, Z.; Sun, L.; Hu, W.; Cao, J.; Lin, W.; Lin, X. Proteomics Analysis Reveals a Potential Antibiotic Cocktail Therapy Strategy for Aeromonas hydrophila Infection in Biofilm. J. Proteome Res. 2016, 15, 1810–1820. [Google Scholar] [CrossRef]
- Du, H.; Pang, M.; Dong, Y.; Wu, Y.; Wang, N.; Liu, J.; Awan, F.; Lu, C.; Liu, Y. Identification and Characterization of an Aeromonas hydrophila Oligopeptidase Gene pepF Negatively Related to Biofilm Formation. Front. Microbiol. 2016, 7, 1497. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abrami, L.; Fivaz, M.; Glauser, P.E.; Sugimoto, N.; Zurzolo, C.; van der Goot, F.G. Sensitivity of polarized epithelial cells to the pore-forming toxin aerolysin. Infection Immun. 2003, 71, 739–746. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Annunziato, G. Strategies to Overcome Antimicrobial Resistance (AMR) Making Use of Non-Essential Target Inhibitors: A Review. Int. J. Mol. Sci. 2019, 20, 5844. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hotinger, J.A.; May, A.E. Animal Models of Type III Secretion System-Mediated Pathogenesis. Pathogens 2019, 8, 257. [Google Scholar] [CrossRef] [Green Version]
- Wu, S.C.; Liu, F.; Zhu, K.; Shen, J.Z. Natural Products That Target Virulence Factors in Antibiotic-Resistant Staphylococcus aureus. J. Agric. Food Chem. 2019, 67, 13195–13211. [Google Scholar] [CrossRef]
- Allen, R.C.; Popat, R.; Diggle, S.P.; Brown, S.P. Targeting virulence: Can we make evolution-proof drugs? Nat. Rev. Microbiol. 2014, 12, 300–308. [Google Scholar] [CrossRef]
- Fernebro, J. Fighting bacterial infections-future treatment options. Drug Resist. Updates Rev. Comment. Antimicrob. Anticancer. Chemother. 2011, 14, 125–139. [Google Scholar] [CrossRef]
- Clatworthy, A.E.; Pierson, E.; Hung, D.T. Targeting virulence: A new paradigm for antimicrobial therapy. Nat. Chem. Biol. 2007, 3, 541–548. [Google Scholar] [CrossRef]
- Taylor, N.G.; Verner-Jeffreys, D.W.; Baker-Austin, C. Aquatic systems: Maintaining, mixing and mobilising antimicrobial resistance? Trends Ecol. Evol. 2011, 26, 278–284. [Google Scholar] [CrossRef]
- Los, F.C.; Randis, T.M.; Aroian, R.V.; Ratner, A.J. Role of pore-forming toxins in bacterial infectious diseases. Microbiol. Mol. Biol. Rev. MMBR 2013, 77, 173–207. [Google Scholar] [CrossRef] [Green Version]
- Degiacomi, M.T.; Iacovache, I.; Pernot, L.; Chami, M.; Kudryashev, M.; Stahlberg, H.; van der Goot, F.G.; Dal Peraro, M. Molecular assembly of the aerolysin pore reveals a swirling membrane-insertion mechanism. Nat. Chem. Biol. 2013, 9, 623–629. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rama Devi, K.; Srinivasan, R.; Kannappan, A.; Santhakumari, S.; Bhuvaneswari, M.; Rajasekar, P.; Prabhu, N.M.; Veera Ravi, A. In vitro and in vivo efficacy of rosmarinic acid on quorum sensing mediated biofilm formation and virulence factor production in Aeromonas hydrophila. Biofouling 2016, 32, 1171–1183. [Google Scholar] [CrossRef] [PubMed]
- Morselli, M.B.; Baldissera, M.D.; Souza, C.F.; Reis, J.H.; Baldisserotto, B.; Sousa, A.A.; Zimmer, F.; Lopes, D.L.A.; Petrolli, T.G.; Da Silva, A.S. Effects of thymol supplementation on performance, mortality and branchial energetic metabolism in grass carp experimentally infected by Aeromonas hydrophila. Microb. Pathog. 2019, 139, 103915. [Google Scholar] [CrossRef] [PubMed]
- da Cunha, J.A.; Bandeira Junior, G.; da Silva, E.G.; de Avila Scheeren, C.; Fausto, V.P.; Salbego, J.; de Almeida Vaucher, R.; de Vargas, A.C.; Baldisserotto, B. The survival and hepatic and muscle glucose and lactate levels of Rhamdia quelen inoculated with Aeromonas hydrophila and treated with terpinen-4-ol, carvacrol or thymol. Microb. Pathog. 2019, 127, 220–224. [Google Scholar] [CrossRef]
- Bandeira Junior, G.; Sutili, F.J.; Gressler, L.T.; Ely, V.L.; Silveira, B.P.; Tasca, C.; Reghelin, M.; Matter, L.B.; Vargas, A.P.C.; Baldisserotto, B. Antibacterial potential of phytochemicals alone or in combination with antimicrobials against fish pathogenic bacteria. J. Appl. Microbiol. 2018, 125, 655–665. [Google Scholar] [CrossRef]
- Yuan, W.; Yuk, H.G. Effects of Sublethal Thymol, Carvacrol, and trans-Cinnamaldehyde Adaptation on Virulence Properties of Escherichia coli O157:H7. Appl. Environ. Microbiol. 2019, 85. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Liu, Y.; Qiu, J.; Luo, Z.Q.; Deng, X. The Herbal Compound Thymol Protects Mice From Lethal Infection by Salmonella Typhimurium. Front. Microbiol. 2018, 9, 1022. [Google Scholar] [CrossRef] [Green Version]
- Yin, D.; Du, E.; Yuan, J.; Gao, J.; Wang, Y.; Aggrey, S.E.; Guo, Y. Supplemental thymol and carvacrol increases ileum Lactobacillus population and reduces effect of necrotic enteritis caused by Clostridium perfringes in chickens. Sci. Rep. 2017, 7, 7334. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Sequence (5′-3′) | PCR Amplicon (bp) | Reference |
|---|---|---|---|
| aerA-F | TCTACCACCACCTCCCTGTC | 218 | [3] |
| aerA-R | GACGAAGGTGTGGTTCCAGT | ||
| ahyR-F | TTTACGGGTGACCTGATTGAG | 206 | [18] |
| ahyR-R | CCTGGATGTCCAACTACATCTT | ||
| ahyI-F | GTCAGCTCCCACACGTCGTT | 202 | this study |
| ahyI-R | GGGATGTGGAATCCCACCGT | ||
| 16S rRNA-F | TAATACCGCATACGCCCTAC | 164 | [3] |
| 16S rRNA-R | ACCGTGTCTCAGTTCCAGTG |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dong, J.; Zhang, L.; Liu, Y.; Xu, N.; Zhou, S.; Yang, Q.; Yang, Y.; Ai, X. Thymol Protects Channel Catfish from Aeromonas hydrophila Infection by Inhibiting Aerolysin Expression and Biofilm Formation. Microorganisms 2020, 8, 636. https://doi.org/10.3390/microorganisms8050636
Dong J, Zhang L, Liu Y, Xu N, Zhou S, Yang Q, Yang Y, Ai X. Thymol Protects Channel Catfish from Aeromonas hydrophila Infection by Inhibiting Aerolysin Expression and Biofilm Formation. Microorganisms. 2020; 8(5):636. https://doi.org/10.3390/microorganisms8050636
Chicago/Turabian StyleDong, Jing, Lushan Zhang, Yongtao Liu, Ning Xu, Shun Zhou, Qiuhong Yang, Yibin Yang, and Xiaohui Ai. 2020. "Thymol Protects Channel Catfish from Aeromonas hydrophila Infection by Inhibiting Aerolysin Expression and Biofilm Formation" Microorganisms 8, no. 5: 636. https://doi.org/10.3390/microorganisms8050636
APA StyleDong, J., Zhang, L., Liu, Y., Xu, N., Zhou, S., Yang, Q., Yang, Y., & Ai, X. (2020). Thymol Protects Channel Catfish from Aeromonas hydrophila Infection by Inhibiting Aerolysin Expression and Biofilm Formation. Microorganisms, 8(5), 636. https://doi.org/10.3390/microorganisms8050636