Anti-Inflammatory Effects of Vitamin E in Response to Candida albicans


Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. Agonists
2.3. Antagonists
2.4. Experimental Conditions
2.5. Gene Expression Analysis
2.5.1. Affymetrix Microarrays
2.5.2. Pathway-Focused Gene Expression: RT2 Profiler PCR Arrays
2.6. ELISA
3. Results
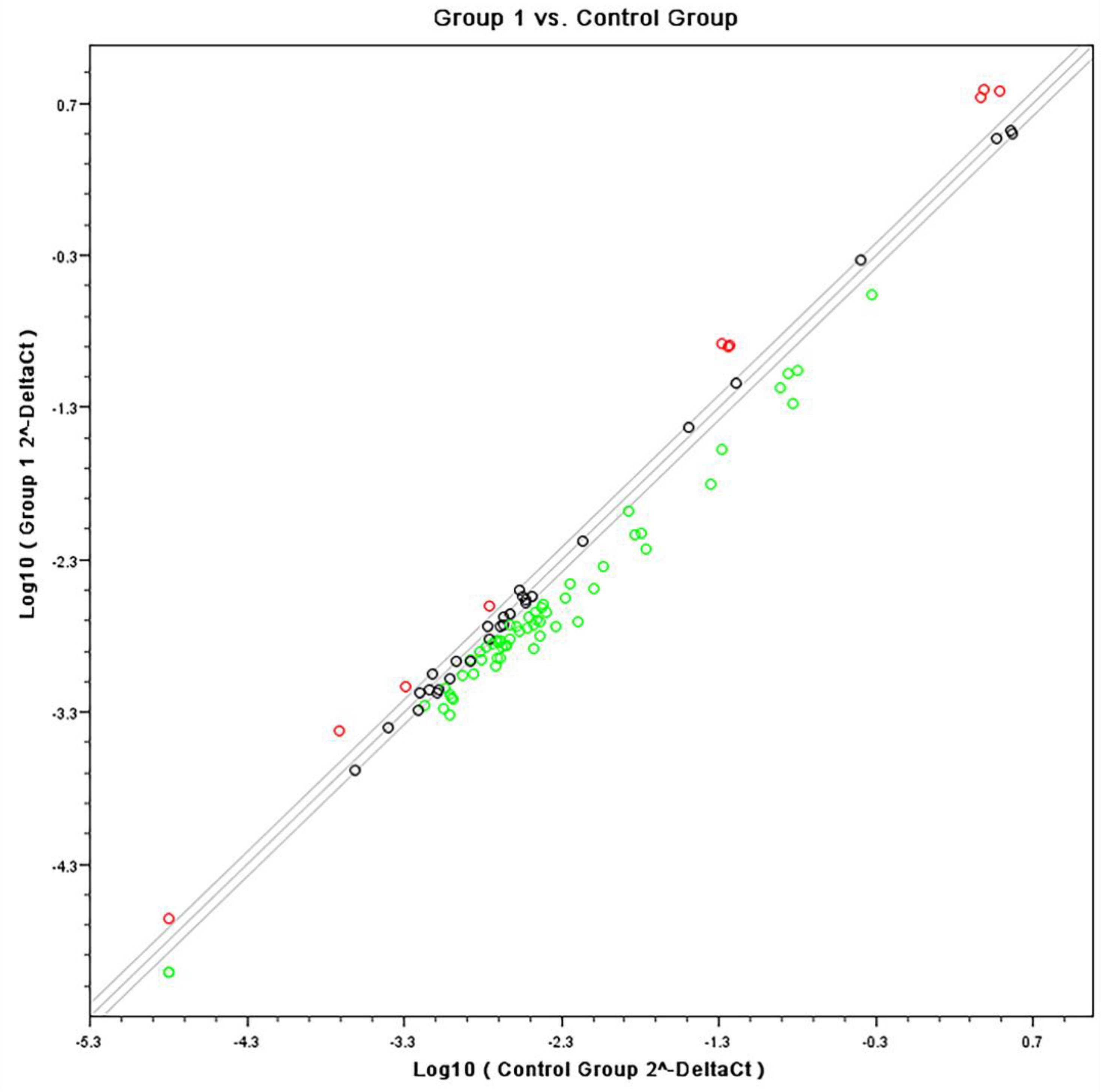
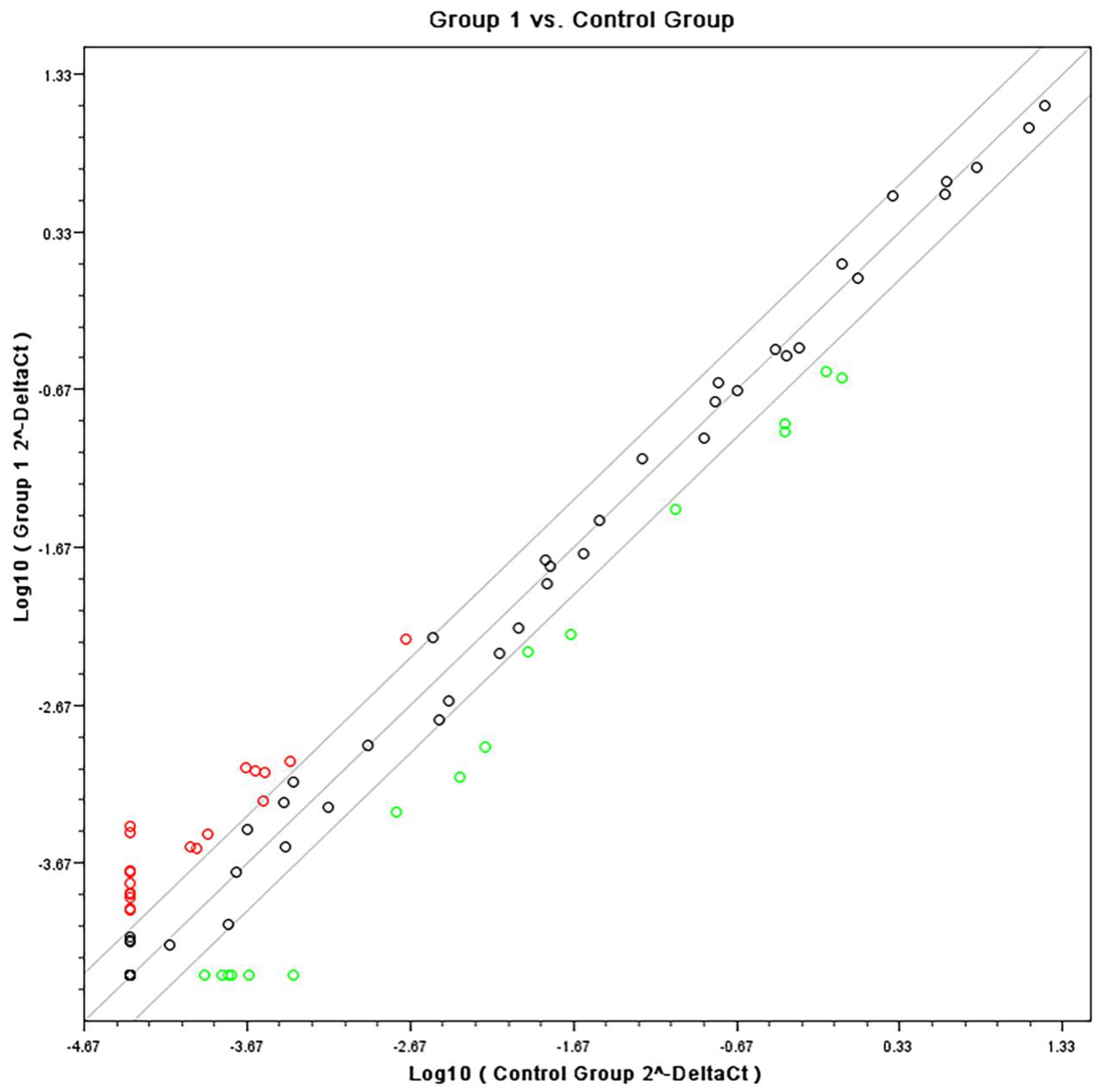
3.1. Gene Expression Array by Affymetrix
3.2. Gene Expression-RT2 Profiler PCR Arrays
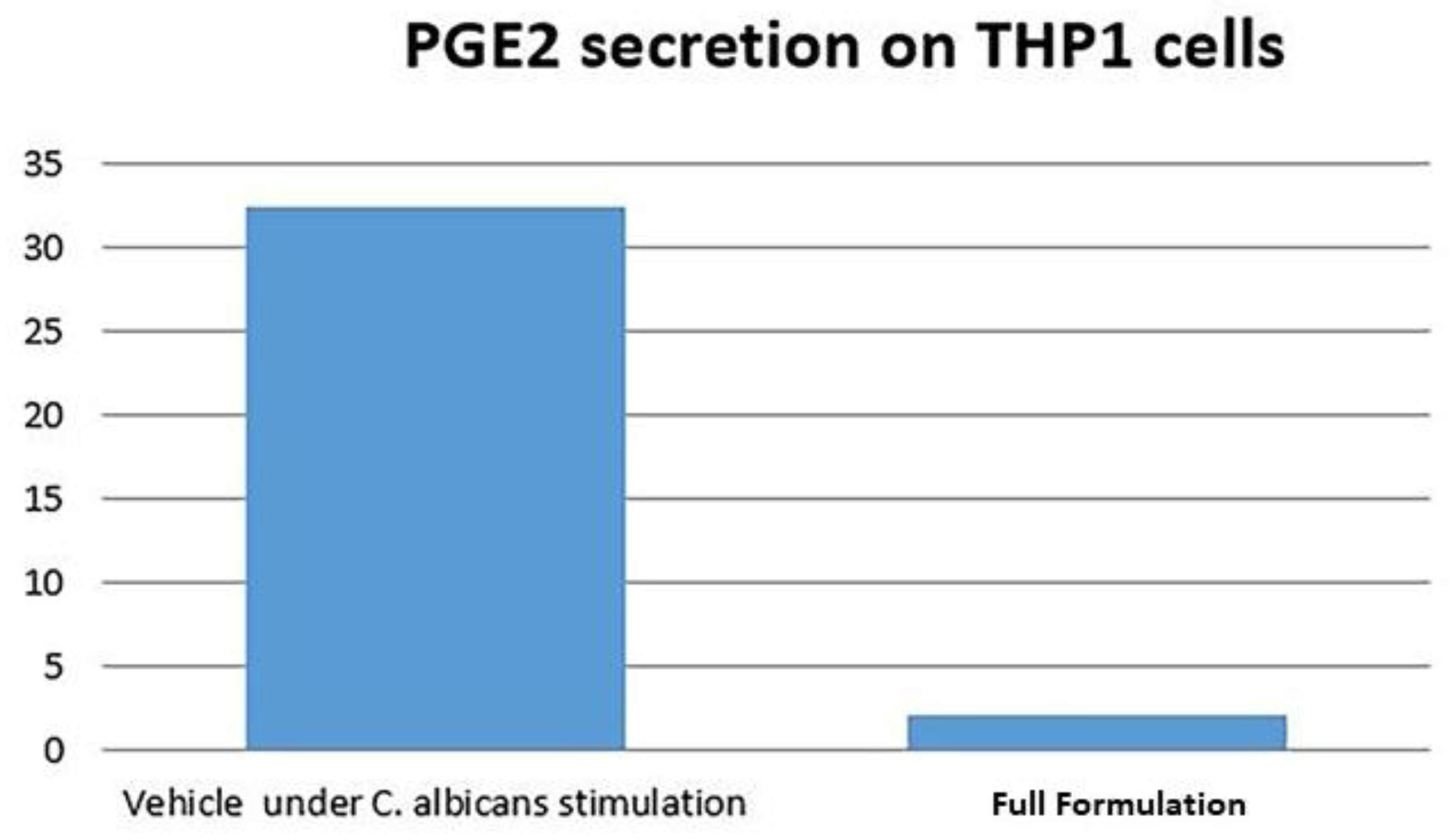
3.3. ELISA Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Samaranayake, L.P.; Keung Leung, W.; Jin, L. Oral mucosal fungal infections. Periodontology 2000 2009, 49, 39–59. [Google Scholar] [CrossRef]
- Gendreau, L.; Loewy, Z.G. Epidemiology and etiology of denture stomatitis. J. Prosthodont. 2011, 20, 251–260. [Google Scholar] [CrossRef] [PubMed]
- Friedman, E.; Alizadeh, N.; Loewy, Z. Oral health: The need for both conventional microbial and molecular characterization. High Throughput 2017, 6, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calderone, R.A.; Fonzi, W.A. Virulence factors of Candida albicans. Trends Microbiol. 2001, 9, 327–335. [Google Scholar] [CrossRef]
- Wu, T.; Shi, W.; Loewy, Z.; He, X. Managing denture biofilm related diseases. Dent. Open J. 2015, 2, 80–86. [Google Scholar] [CrossRef]
- Loewy, Z.G.; Galbut, S.; Loewy, E.; Felton, D. Influence of the Oral Microbiome on General Health; Intechopen: London, UK, 2018. [Google Scholar] [CrossRef]
- Verran, J.; Jackson, S.; Coulthwaite, L.; Scallan, A.; Loewy, Z.; Whitehead, K. The effect of dentifrice abrasion on denture topography and the subsequent retention of microorganisms on abraded surfaces. J. Prosthet. Dent. 2014, 112, 1513–1522. [Google Scholar] [CrossRef]
- Ashman, R.B.; Papadimitriou, J.M. Production and function of cytokines in natural and acquired immunity to Candida albicans infection. Microbiol. Rev. 1995, 59, 646–672. [Google Scholar] [CrossRef]
- Akpan, A.; Morgan, R. Oral candidiasis. Postgrad. Med. J. 2002, 78, 455–459. [Google Scholar] [CrossRef]
- Vanden Abbeele, A.; de Meel, H.; Ahariz, M.; Perraudin, J.P.; Beyer, I.; Courtois, P. Denture contamination by yeasts in the elderly. Gerodontology 2008, 25, 222–228. [Google Scholar] [CrossRef] [Green Version]
- Zhu, W.; Filler, S.G. Interactions of Candida albicans with epithelial cells. Cell Microbiol. 2010, 12, 273–282. [Google Scholar] [CrossRef] [Green Version]
- Offenbacher, S.; Barros, S.; Bencharit, S.; Yu, N.; Preisser, J.; Moss, K.; Loewy, Z.G. Differential mucosal gene expression patterns in Candidiasis-associated, chronic oral denture stomatitis. J. Prosthodont. 2019, 28, 2020–2026. [Google Scholar]
- Dongari-Bagtzoglou, A.; Fidel, P.L., Jr. The host cytokine responses and protective immunity in oropharyngeal candidiasis. J. Dent. Res. 2005, 84, 966–977. [Google Scholar] [CrossRef] [PubMed]
- Ohta, K.; Nishi, H.; Fukui, A.; Shigeishi, H.; Takechi, M.; Kamata, N. CX3CL1 expression induced by Candida albicans in oral fibroblasts. FEMS Immunol. Med. Microbiol. 2010, 60, 179–185. [Google Scholar] [CrossRef] [PubMed]
- Weindl, G.; Wagener, J.; Schaller, M. Epithelial cells and innate antifungal defense. J. Dent. Res. 2010, 89, 666–675. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suzuki, T.; Tsuzuki, A.; Ohno, N.; Ohshima, Y.; Yadomae, T. Enhancement of IL-8 production from human monocytic and granulocytic cell lines, THP-1 and HL-60, stimulated with Malassezia furfur. FEMS Immunol. Med. Microbiol. 2000, 28, 157–162. [Google Scholar] [CrossRef]
- Li, M.; Liu, Z.H.; Chen, Q.; Zhou, W.Q.; Yu, M.W.; Lü, G.X.; Lü, X.L.; Shen, Y.N.; da Liu, W.; Wu, S.X. Insoluble beta-glucan from the cell wall of Candida albicans induces immune responses of human THP-1 monocytes through Dectin-1. Chin. Med. J. 2009, 122, 496–501. [Google Scholar]
- Jeng, K.C.; Yang, C.S.; Siu, W.Y.; Tsai, Y.S.; Liao, W.J.; Kuo, J.S. Supplementation with vitamins C and E enhances cytokine production by peripheral blood mononuclear cells in healthy adults. Am. J. Clin. Nutr. 1996, 64, 960–965. [Google Scholar] [CrossRef] [Green Version]
- Pallast, E.G.; Schouten, E.G.; de Waart, F.G.; Fonk, H.C.; Doekes, G.; von Blomberg, B.M.; Kok, F.J. Effect of 50- and 100-mg vitamin E supplements on cellular immune function in noninstitutionalized elderly persons. Am. J. Clin. Nutr. 1999, 69, 1273–1281. [Google Scholar] [CrossRef]
- Wintergerst, E.S.; Maggini, S.; Hornig, D.H. Contribution of selected vitamins and trace elements to immune function. Ann. Nutr. Metab. 2007, 51, 301–323. [Google Scholar] [CrossRef] [Green Version]
- Härtel, C.; Strunk, T.; Bucsky, P.; Schultz, C. Effects of vitamin C on intracytoplasmic cytokine production in human whole blood monocytes and lymphocytes. Cytokine 2004, 27, 101–106. [Google Scholar] [CrossRef]
- Wu, S.J.; Liu, P.L.; Ng, L.T. Tocotrienol-rich fraction of palm oil exhibits anti-inflammatory property by suppressing the expression of inflammatory mediators in human monocytic cells. Mol. Nutr. Food Res. 2008, 52, 921–929. [Google Scholar] [CrossRef] [PubMed]
- Rowe, R.C.; Sheskey, P.J.; Owen, S.C. Handbook of Pharmaceutical Excipients, 5th ed.; Pharmaceutical Press: London, UK; American Pharmaceutical Association: Chicago, IL, USA, 2005. [Google Scholar]
- Chung, M.Y.; Yeung, S.F.; Park, H.J.; Volek, J.S.; Bruno, R.S. Dietary α- and γ-tocopherol supplementation attenuates lipopolysaccharide-induced oxidative stress and inflammatory-related responses in an obese mouse model of nonalcoholic steatohepatitis. J. Nutr. Biochem. 2010, 21, 1200–1206. [Google Scholar] [CrossRef] [PubMed]
- Cook-Mills, J.M.; McCary, C.A. Isoforms of vitamin E differentially regulate inflammation. Endocr. Metab. Immune Disord. Drug Targets 2010, 10, 348–366. [Google Scholar] [CrossRef] [PubMed]
- Jackson, S.; Coulthwaite, L.; Loewy, Z.; Scallan, A.; Verran, J. Biofilm development by blastospores and hyphae of Candida albicans on abraded denture acrylic surfaces. J. Prosthet. Dent. 2014, 112, 988–993. [Google Scholar] [CrossRef] [PubMed]
- Berdnikovs, S.; Abdala-Valencia, H.; McCary, C.; Somand, M.; Cole, R.; Garcia, A.; Bryce, P.; Cook-Mills, J.M. Isoforms of vitamin E have opposing immunoregulatory functions during inflammation by regulating leukocyte recruitment. J. Immunol. 2009, 182, 4395–4405. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barker, K.S.; Liu, T.; Rogers, P.D. Coculture of THP-1 human mononuclear cells with Candida albicans results in pronounced changes in host gene expression. J. Infect. Dis. 2005, 192, 901–912. [Google Scholar] [CrossRef] [Green Version]
- Schaller, M.; Borelli, C.; Korting, H.C.; Hube, B. Hydrolytic enzymes as virulence factors of Candida albicans. Mycoses 2005, 48, 365–377. [Google Scholar] [CrossRef]
- Egusa, H.; Nikawa, H.; Makihira, S.; Yatani, H.; Hamada, T. In vitro mechanisms of interleukin-8-mediated responses of human gingival epithelial cells to Candida albicans infection. Int. J. Med. Microbiol. 2006, 296, 301–311. [Google Scholar] [CrossRef]
- Jouault, T.; Bernigaud, A.; Lepage, G.; Trinel, P.A.; Poulain, D. The Candida albicans phospholipomannan induces in vitro production of tumour necrosis factor-alpha from human and murine macrophages. Immunology 1994, 83, 268–273. [Google Scholar]
- Pivarcsi, A.; Nagy, I.; Koreck, A.; Kis, K.; Kenderessy-Szabo, A.; Szell, M.; Dobozy, A.; Kemeny, L. Microbial compounds induce the expression of pro-inflammatory cytokines, chemokines and human beta-defensin-2 in vaginal epithelial cells. Microbes Infect. 2005, 7, 1117–1127. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, Y.; Klein, T.W.; Friedman, H. Involvement of mannose receptor in cytokine interleukin-1beta (IL-1beta), IL-6, and granulocyte-macrophage colony-stimulating factor responses, but not in chemokine macrophage inflammatory protein 1beta (MIP-1beta), MIP-2, and KC responses, caused by attachment of Candida albicans to macrophages. Infect. Immun. 1997, 65, 1077–1082. [Google Scholar]
- Netea, M.G.; Gow, N.A.; Munro, C.A.; Bates, S.; Collins, C.; Ferwerda, G.; Hobson, R.P.; Bertram, G.; Hughes, H.B.; Jansen, T.; et al. Immune sensing of Candida albicans requires cooperative recognition of mannans and glucans by lectin and Toll-like receptors. J. Clin. Investig. 2006, 116, 1642–1650. [Google Scholar] [CrossRef]
- Netea, M.G.; Brown, G.D.; Kullberg, B.J.; Gow, N.A. An integrated model of the recognition of Candida albicans by the innate immune system. Nat. Rev. Microbiol. 2008, 6, 67–78. [Google Scholar] [CrossRef] [PubMed]
- Moyes, D.L.; Runglall, M.; Murciano, C.; Shen, C.; Nayar, D.; Thavaraj, S.; Kohli, A.; Islam, A.; Mora-Montes, H.; Challacombe, S.J.; et al. A biphasic innate immune MAPK response discriminates between the yeast and hyphal forms of Candida albicans in epithelial cells. Cell Host Microbe 2010, 8, 225–235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rouabhia, M.; Mukherjee, P.K.; Lattif, A.A.; Curt, S.; Chandra, J.; Ghannoum, M.A. Disruption of sphingolipid biosynthetic gene IPT1 reduces Candida albicans adhesion and prevents activation of human gingival epithelial cell innate immune defense. Med. Mycol. 2011, 49, 458–466. [Google Scholar]
- Akira, S.; Uematsu, S.; Takeuchi, O. Pathogen recognition and innate immunity. Cell 2006, 124, 783–801. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takeuchi, O.; Kawai, T.; Mühlradt, P.F.; Morr, M.; Radolf, J.D.; Zychlinsky, A.; Takeda, K.; Akira, S. Discrimination of bacterial lipoproteins by Toll-like receptor 6. Int. Immunol. 2001, 13, 933–940. [Google Scholar] [CrossRef]
- Triantafilou, M.; Gamper, F.G.; Haston, R.M.; Mouratis, M.A.; Morath, S.; Hartung, T.; Triantafilou, K. Membrane sorting of toll-like receptor (TLR)-2/6 and TLR2/1 heterodimers at the cell surface determines heterotypic associations with CD36 and intracellular targeting. J. Biol. Chem. 2006, 281, 31002–31011. [Google Scholar] [CrossRef] [Green Version]
- Ahn, K.S.; Sethi, G.; Krishnan, K.; Aggarwal, B.B. Gammatocotrienol inhibits nuclear factor-kappaB signaling pathway through inhibition of receptor-interacting protein and TAK1 leading to suppression of antiapoptotic gene products and potentiation of apoptosis. J. Biol. Chem. 2007, 282, 809–820. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Test Formulation | Control (Vehicle) | |
|---|---|---|
| Carboxymethyl cellulose (CMC) | yes | yes |
| Mineral Oil, Light | yes | yes |
| Petrolatum Blend | yes | yes |
| Double Salt | yes | yes |
| Vitamin E Acetate | yes | 0.00% |
| Ethyl Paraben | yes | 0.00% |
| Methyl Paraben | yes | 0.00% |
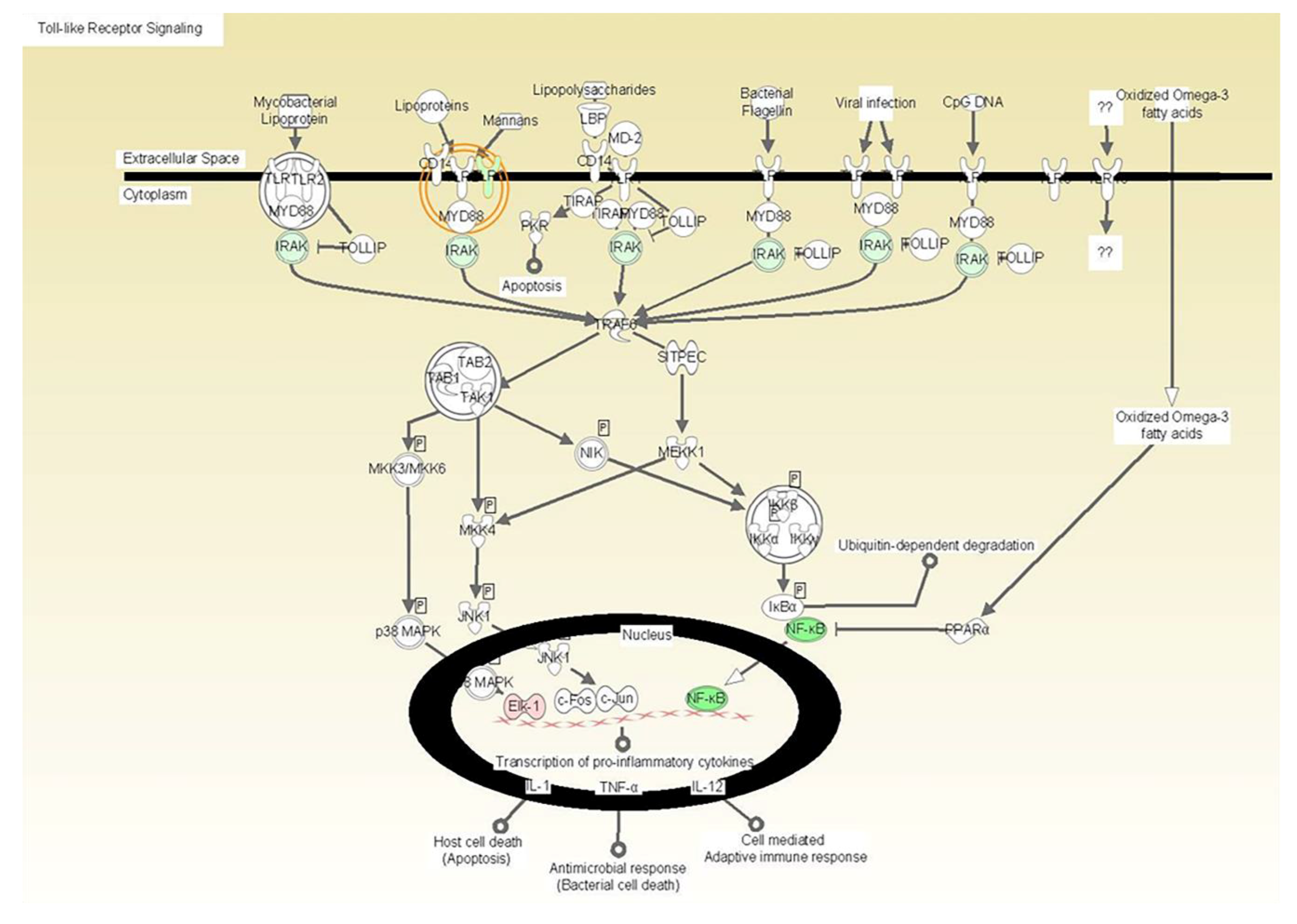
| -TLR 6 synthesis and signaling [processing carbohydrates and mannans associated with fungi] |
| -IL-1 synthesis and signaling [IRAK3, A20] |
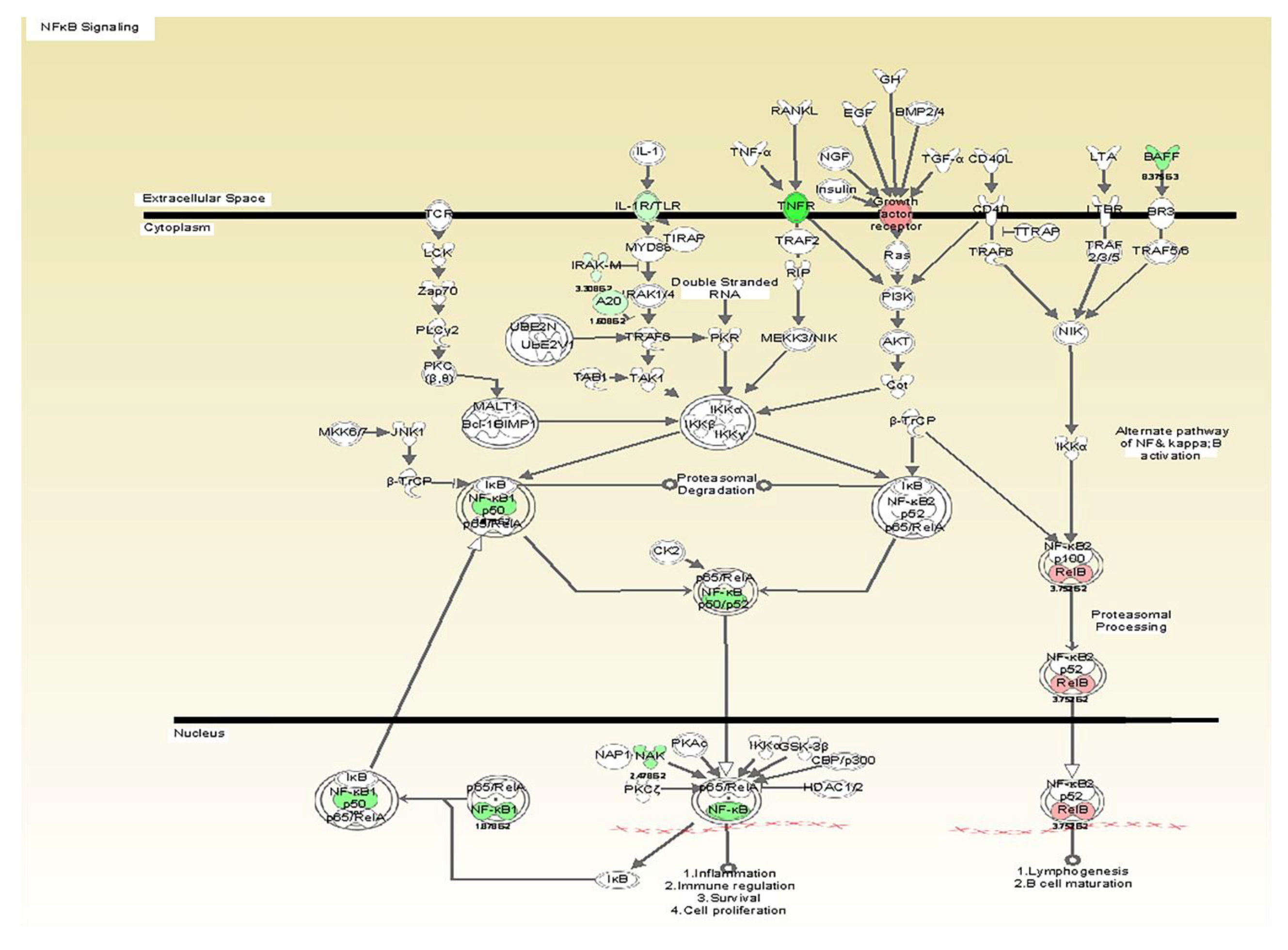
| -NF-kappaB: A master transcriptional regulator that enhances the expression of multiple inflammatory mediators. |
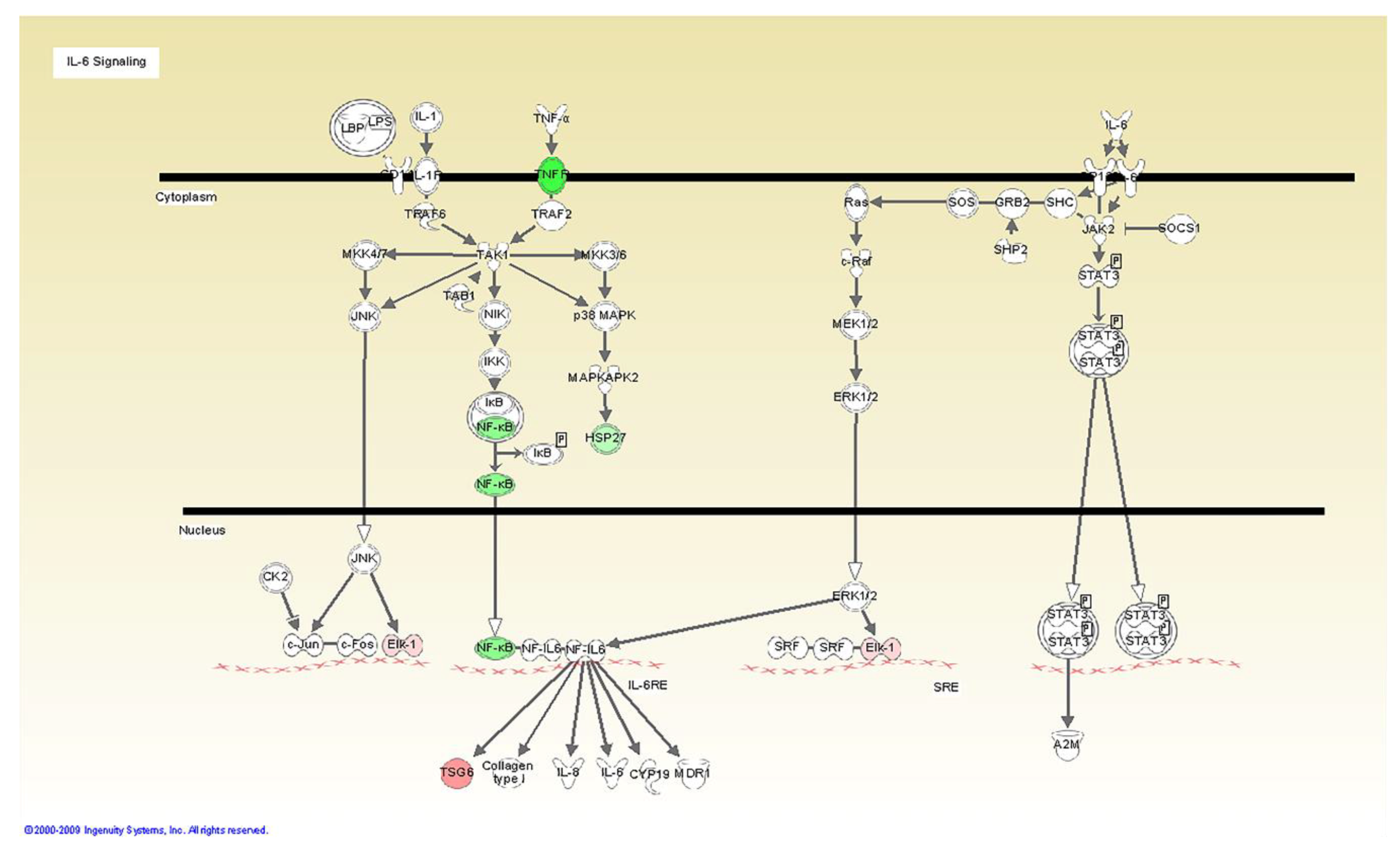
| -IL-6 synthesis and signaling [gp130, JK2 and GRB2] |
| -TNF synthesis and signaling [TNF receptor] |
| Gene Symbol | Adhesive + C. albicans THP1 |
|---|---|
| IL1A | −1.5751 |
| IL1B | −1.0392 |
| IL6 | −1.3753 |
| IL8 | −2.8064 |
| IL10 | −1.5208 |
| IL12A | −1.7109 |
| Gene Symbol | THP1 Full Formulation + C. albicans | Fibroblasts Full Formulation + C. albicans |
|---|---|---|
| IFNA1 | 1.3822 | |
| IFNA2 | −1.1669 | |
| IFNA4 | −1.8824 | |
| IFNA5 | −1.619 | |
| IFNA8 | −1.619 | |
| IFNB1 | 1.1665 | 2.6224 |
| IFNG | −1.2247 | 3.9441 |
| Gene Symbol | Fibroblasts P&P C. albicans |
|---|---|
| TNFSF11 | −4.1403 |
| TNFSF12 | −1.1864 |
| TNFSF13 | 1.7268 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Barros, S.; D. Ribeiro, A.P.; Offenbacher, S.; Loewy, Z.G. Anti-Inflammatory Effects of Vitamin E in Response to Candida albicans. Microorganisms 2020, 8, 804. https://doi.org/10.3390/microorganisms8060804
Barros S, D. Ribeiro AP, Offenbacher S, Loewy ZG. Anti-Inflammatory Effects of Vitamin E in Response to Candida albicans. Microorganisms. 2020; 8(6):804. https://doi.org/10.3390/microorganisms8060804
Chicago/Turabian StyleBarros, Silvana, Ana Paula D. Ribeiro, Steven Offenbacher, and Zvi G. Loewy. 2020. "Anti-Inflammatory Effects of Vitamin E in Response to Candida albicans" Microorganisms 8, no. 6: 804. https://doi.org/10.3390/microorganisms8060804
APA StyleBarros, S., D. Ribeiro, A. P., Offenbacher, S., & Loewy, Z. G. (2020). Anti-Inflammatory Effects of Vitamin E in Response to Candida albicans. Microorganisms, 8(6), 804. https://doi.org/10.3390/microorganisms8060804