Age and Species of Eucalyptus Plantations Affect Soil Microbial Biomass and Enzymatic Activities



Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site and Soil Sample Collection
2.2. Soil Microbial Biomass Analysis
2.3. DNA Extraction and Real-Time Quantitative PCR for Fungal and Bacterial Biomass
2.4. Soil Enzyme Activity Analysis
2.5. Statistical Analysis
3. Results
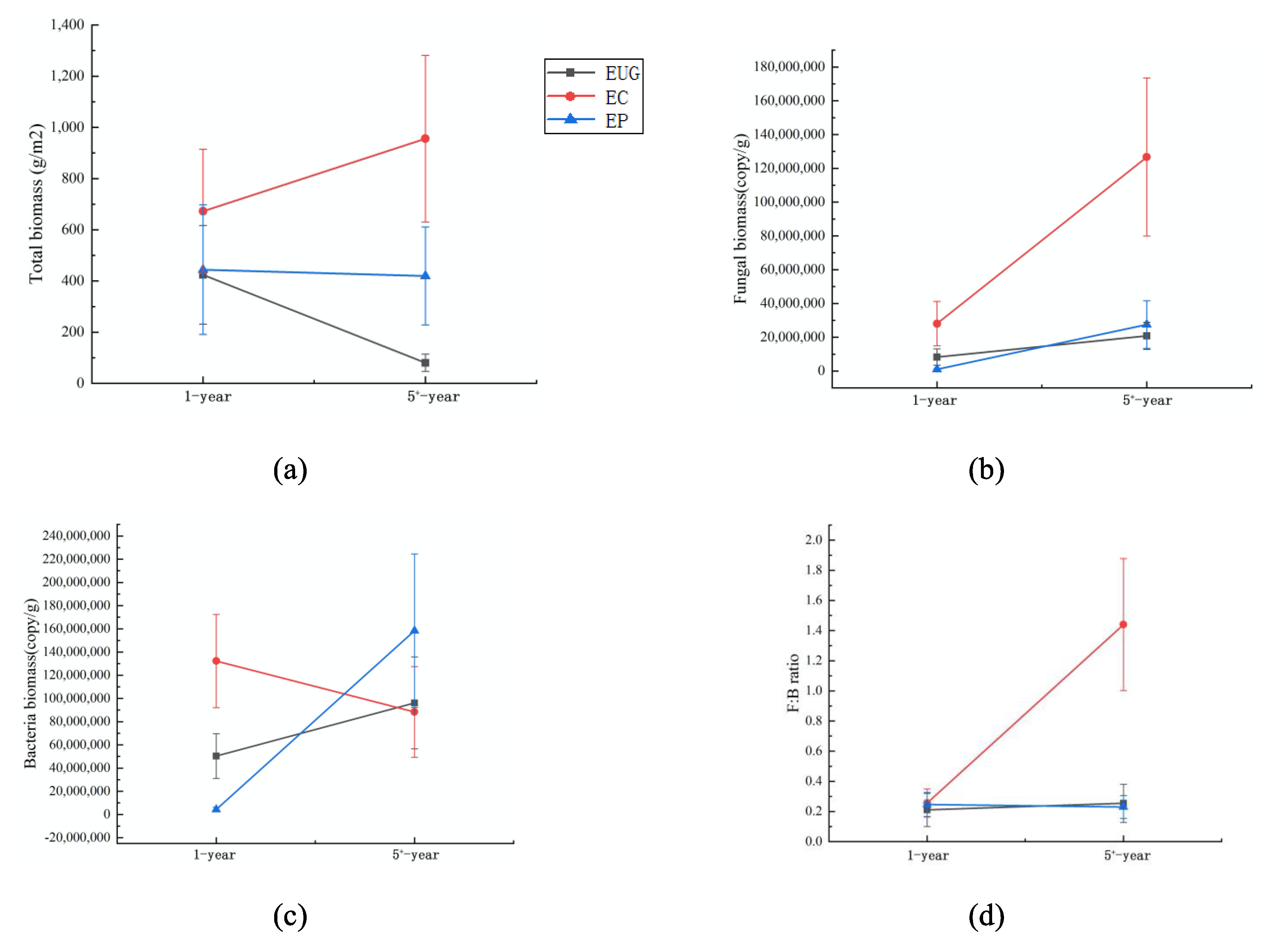
3.1. Soil Total Microbial Biomass and Fungal and Bacterial Biomass in Different Plantations
3.2. Soil Enzyme Activities in Different Plantations
4. Discussion
4.1. Soil Total Microbial Biomass
4.2. Soil Enzyme Activities
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Smith, J.L.; Paul, E.; Lefroy, R.D.B.; Blair, G.J.; Graswell, E.T. The significance of soil microbial biomass estimations. Soil Biochem. 1990, 6, 357–396. [Google Scholar]
- Han, W.; Kemmitt, S.J.; Brookes, P.C. Soil microbial biomass and activity in Chinese tea gardens of varying stand age and productivity. Soil Biol. Biochem. 2007, 39, 1468–1478. [Google Scholar] [CrossRef]
- Wick, B.; Kühne, R.F.; Vlek, P.L.G. Soil microbiological parameters as indicators of soil quality under improved fallow management systems in south-western Nigeria. Plant. Soil 1998, 202, 97–107. [Google Scholar] [CrossRef]
- Aon, M.A.; Colaneri, A.C., II. Temporal and spatial evolution of enzymatic activities and physico-chemical properties in an agricultural soil. Appl. Soil Ecol. 2001, 18, 255–270. [Google Scholar] [CrossRef]
- Baum, C.; Leinweber, P.; Schlichting, A. Effects of chemical conditions in re-wetted peats on temporal variation in microbial biomass and acid phosphatase activity within the growing season. Appl. Soil Ecol. 2003, 22, 167–174. [Google Scholar] [CrossRef]
- Sinsabaugh, R.; Lauber, C.; Weintraub, M.; Ahmed, B.; Allison, S.; Crenshaw, C.; Contosta, A.; Cusack, D.; Frey, S.; Gallo, M.; et al. Stoichiometry of soil enzyme activity at global scale. Ecol. Lett. 2008, 11, 1252–1264. [Google Scholar] [CrossRef]
- Lin, Z.; Li, Y.; Tang, C.; Luo, Y.; Fu, W.; Cai, X.; Li, Y.; Yue, T.; Jiang, P.; Hu, S.; et al. Converting natural evergreen broadleaf forests to intensively managed moso bamboo plantations affects the pool size and stability of soil organic carbon and enzyme activities. Biol. Fert. Soils 2018, 54, 467–480. [Google Scholar] [CrossRef]
- Sardans, J.; Peñuelas, J.; Estiarte, M. Changes in soil enzymes related to C and N cycle and in soil C and N content under prolonged warming and drought in a Mediterranean shrubland. Appl. Soil Ecol. 2008, 39, 223–235. [Google Scholar] [CrossRef]
- Brockett, B.F.T.; Prescott, C.E.; Grayston, S.J. Soil moisture is the major factor influencing microbial community structure and enzyme activities across seven biogeoclimatic zones in western Canada. Soil Biol. Biochem. 2012, 44, 9–20. [Google Scholar] [CrossRef]
- Baldrian, P.; Šnajdr, J.; Merhautová, V.; Dobiášová, P.; Cajthaml, T.; Valášková, V. Responses of the extracellular enzyme activities in hardwood forest to soil temperature and seasonality and the potential effects of climate change. Soil Biol. Biochem. 2013, 56, 60–68. [Google Scholar] [CrossRef]
- Yang, M.; Yang, D.; Yu, X. Soil microbial communities and enzyme activities in sea-buckthorn (Hippophae rhamnoides) plantation at different ages. PLoS ONE 2018, 13, e0190959. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Wang, S.; Chen, C. Modeling the effects of tree species and temperature on soil’s extracellular enzyme activity in 78-year-old tree plantations. Biogeosci. Discuss. 2017, 14, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Nannipieri, P.; Trasar-Cepeda, C.; Dick, R.P. Soil enzyme activity: A brief history and biochemistry as a basis for appropriate interpretations and meta-analysis. Biol. Fert. Soils 2017, 54, 11–19. [Google Scholar] [CrossRef]
- Singh, J.S.; Gupta, V.K. Soil microbial biomass: A key soil driver in management of ecosystem functioning. Sci. Total Environ. 2018, 634, 497–500. [Google Scholar] [CrossRef] [PubMed]
- Bréchet, L.; Ponton, S.; Roy, J.; Freycon, V.; Coûteaux, M.-M.; Bonal, D.; Epron, D. Do tree species characteristics influence soil respiration in tropical forests? A test based on 16 tree species planted in monospecific plots. Plant. Soil 2009, 319, 235–246. [Google Scholar] [CrossRef]
- Huang, Z.; Wan, X.; He, Z.; Yu, Z.; Wang, M.; Hu, Z.; Yang, Y. Soil microbial biomass, community composition and soil nitrogen cycling in relation to tree species in subtropical China. Soil Biol. Biochem. 2013, 62, 68–75. [Google Scholar] [CrossRef]
- Noll, L.; Leonhardt, S.; Arnstadt, T.; Hoppe, B.; Poll, C.; Matzner, E.; Hofrichter, M.; Kellner, H. Fungal biomass and extracellular enzyme activities in coarse woody debris of 13 tree species in the early phase of decomposition. For. Ecol. Manag. 2016, 378, 181–192. [Google Scholar] [CrossRef]
- Chatterjee, A.; Ingram, L.; Vance, G.; Stahl, P.D. Soil processes and microbial community structures in 45- and 135-year-old lodgepole pine stands. Can. J. For. Res. 2009, 39, 2263–2271. [Google Scholar] [CrossRef]
- Li, J.; Jiang, X.-M.; Yin, H.-J.; Yin, C.-Y.; Wei, Y.-H.; Liu, Q. Root exudates and soil microbes in three Picea asperata plantations with different stand ages. Ying Yong Sheng Tai Xue Bao = J. Appl. Ecol. 2014, 25, 325–332. [Google Scholar]
- Bragazza, L.; Bardgett, R.; Mitchell, E.; Buttler, A. Linking soil microbial communities to vascular plant abundance along a climate gradient. New Phytol. 2014, 205. [Google Scholar] [CrossRef]
- Bauhus, J.; Paré, D.; Coté, L. Effects of tree species, stand age and soil type on soil microbial biomass and its activity in a southern boreal forest. Soil Biol. Biochem. 1998, 30, 1077–1089. [Google Scholar] [CrossRef]
- Grattapaglia, D.; Vaillancourt, R.E.; Shepherd, M.; Thumma, B.R.; Foley, W.; Külheim, C.; Potts, B.M.; Myburg, A.A. Progress in Myrtaceae genetics and genomics: Eucalyptus as the pivotal genus. Tree Genet. Genomes 2012, 8, 463–508. [Google Scholar] [CrossRef] [Green Version]
- Arnold, R.J.; Luo, J.Z.; Clarke, B. Trials of Cold-tolerant Eucalypt Species in Cooler Regions of South Central China; Australian Centre for International Agricultural Research: Canberra, Australia, 2004; pp. 11–37.
- Zhang, C.; Li, X.; Chen, Y.; Zhao, J.; Wan, S.; Lin, Y.; Fu, S. Effects of Eucalyptus litter and roots on the establishment of native tree species in Eucalyptus plantations in South China. For. Ecol. Manag. 2016, 375, 76–83. [Google Scholar] [CrossRef]
- Bertin, C.; Yang, X.; Weston, L.; Bertin, C.; Yang, X.H.; Weston, L.A. The role of root exudates and allelochemicals in the rhizosphere. Plant. Soil 2003, 256, 67–83. [Google Scholar] [CrossRef]
- Laclau, J.-P.; Ranger, J.; de Moraes Gonçalves, J.L.; Maquère, V.; Krusche, A.V.; M’Bou, A.T.; Nouvellon, Y.; Saint-André, L.; Bouillet, J.-P.; de Cassia Piccolo, M.; et al. Biogeochemical cycles of nutrients in tropical Eucalyptus plantations. For. Ecol. Manag. 2010, 259, 1771–1785. [Google Scholar] [CrossRef]
- Leite, F.; Silva, I.; Novais, R.; Barros, N.; Neves, J. Alterations of soil chemical properties by Eucalyptus cultivation in five regions in the Rio Doce Valley. Rev. Bras. Cienc. Solo 2010, 34. [Google Scholar] [CrossRef] [Green Version]
- Versini, A.; Mareschal, L.; Matsoumbou, T.; Zeller, B.; Ranger, J.; Laclau, J.-P. Effects of litter manipulation in a tropical Eucalyptus plantation on leaching of mineral nutrients, dissolved organic nitrogen and dissolved organic carbon. Geoderma 2014, 232-234, 426–436. [Google Scholar] [CrossRef]
- Mishra, A.; Sharma, S.D.; Khan, G.H. Improvement in physical and chemical properties of sodic soil by 3, 6 and 9 years old plantation of Eucalyptus tereticornis: Biorejuvenation of sodic soil. For. Ecol. Manag. 2003, 184, 115–124. [Google Scholar] [CrossRef]
- Yu, X.B.; Xu, D.P. Studies on the growth characteristics and the crown structure of Eucalyptus plantation with the different continuous-planting rotation. Sci. Silva. Sin. 2000, 36, 137–142. [Google Scholar]
- Madejón, P.; Marañón, T.; Navarro-Fernández, C.M.; Domínguez, M.T.; Alegre, J.M.; Robinson, B.; Murillo, J.M. Potential of Eucalyptus camaldulensis for phytostabilization and biomonitoring of trace-element contaminated soils. PLoS ONE 2017, 12, e0180240. [Google Scholar] [CrossRef] [Green Version]
- Inail, M.; Hardiyanto, E.; Mendham, D.S. Growth Responses of Eucalyptus pellita F. Muell Plantations in South Sumatra to Macronutrient Fertilisers Following Several Rotations of Acacia mangium Willd. Forests 2019, 10, 1054. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.-J.; Xu, D.-P.; Xie, Z.-S.; Zhang, N.-N. Effects of different culture media on the growth of Indian sandalwood (Santalum album L.) seedlings in Zhanjiang, Guangdong, southern China. For. Stud. China 2009, 11, 132–138. [Google Scholar] [CrossRef]
- Liu, W.; Huang, S.Y.; Li, D.; Wang, C.Y. 2016 Fourth International Workshop on Earth Observation and Remote Sensing Applications. In Temperature Variation and Winter Planted Potato’s NDVI Change during early 2016’s Super Cold Wave in Guangdong Province, South China, Guangzhou, China, 2016.7.4-7.6; Institute of Electrical and Electronics Engineers: Guangzhou, China, 2016. [Google Scholar]
- Vance, E.D.; Brookes, P.C.; Jenkinson, D.S. An extraction method for measuring soil microbial biomass C. Soil Biol. Biochem. 1987, 19, 703–707. [Google Scholar] [CrossRef]
- Wu, J.; Joergensen, R.; Pommerening, B.; Chaussod, R.; Brookes, P.C. Measurement of soil microbial biomass C by fumigation-extraction–An automated procedure. Soil Biol. Biochem. 1990, 22, 1167–1169. [Google Scholar] [CrossRef]
- Vainio, E.J.; Hantula, J. Direct analysis of wood-inhabiting fungi using denaturing gradient gel electrophoresis of amplified ribosomal DNA. Mycol. Res. 2000, 104, 927–936. [Google Scholar] [CrossRef]
- Fierer, N.; Jackson, J.; Vilgalys, R.; Jackson, R. Assessment of Soil Microbial Community Structure by Use of Taxon-Specific Quantitative PCR Assays. Appl. Environ. Microbiol. 2005, 71, 4117–4120. [Google Scholar] [CrossRef] [Green Version]
- Helin, A.; Sietiö, O.-M.; Heinonsalo, J.; Bäck, J.; Riekkola, M.-L.; Parshintsev, J. Characterization of free amino acids, bacteria and fungi in size-segregated atmospheric aerosols in boreal forest: Seasonal patterns, abundances and size distributions. Atmos. Chem. Phys. Discuss. 2017, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Zhou, X.; Sun, H.; Pumpanen, J.; Sietiö, O.-M.; Heinonsalo, J.; Köster, K.; Berninger, F. The impact of wildfire on microbial C:N:P stoichiometry and the fungal-to-bacterial ratio in permafrost soil. Biogeochemistry 2018, 142, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Sinsabaugh, R.L.; Saiya-Cork, K.; Long, T.; Osgood, M.P.; Neher, D.A.; Zak, D.R.; Norby, R.J. Soil microbial activity in a Liquidambar plantation unresponsive to CO2-driven increases in primary production. Appl. Soil Ecol. 2003, 24, 263–271. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.; Wan, X.; Yu, Z.; Hu, Z.; He, Z.; Huang, Z. Effects of tree species transition on soil microbial biomass and community structure in subtropical China. Acta Ecol. Sin. 2016, 36, 417–423. [Google Scholar] [CrossRef]
- Hobbie, S.E. Effects of plant species on nutrient cycling. Trends Ecol. Evol. 1992, 7, 336–339. [Google Scholar] [CrossRef]
- Priha, O.; Smolander, A. Nitrogen transformations in soil under Pinus sylvestris, Picea abies and Betula pendula at two forest sites. Soil Biol. Biochem. 1999, 31, 965–977. [Google Scholar] [CrossRef]
- Murphy, D.V.; Cookson, W.R.; Braimbridge, M.; Marschner, P.; Jones, D.L.; Stockdale, E.A.; Abbott, L.K. Relationships between soil organic matter and the soil microbial biomass (size, functional diversity, and community structure) in crop and pasture systems in a semi-arid environment. Soil Res. 2011, 49, 582–594. [Google Scholar] [CrossRef]
- Schnürer, J.; Clarholm, M.; Rosswall, T. Microbial biomass and activity in an agricultural soil with different organic matter contents. Soil Biol. Biochem. 1985, 17, 611–618. [Google Scholar] [CrossRef]
- Pellissier, F.; Souto, C. Allelopathy in Northern Temperate and Boreal Semi Natural Woodland. Crit. Rev. Plant. Sci. 1999, 18, 637–652. [Google Scholar] [CrossRef]
- Huang, C.B.; Wen, Y.G.; Mo, T.S. Study on the soil hydrology-physical properties of Eucalyptus urophylla × E. grandis continuous plantations. Ecol. Environ. 2007, 16, 538–543. [Google Scholar]
- Ren, C.; Zhang, W.; Zhong, Z.; Han, X.; Yang, G.; Feng, Y.; Ren, G. Differential responses of soil microbial biomass, diversity, and compositions to altitudinal gradients depend on plant and soil characteristics. Sci. Total Environ. 2018, 610-611, 750–758. [Google Scholar] [CrossRef]
- Zhang, Q.; Zak, J. Effects of Water and Nitrogen Amendment on Soil Microbial Biomass and Fine Root Production in a Semi-Arid Environment in West Texas. Soil Biol. Biochem. 1998, 30, 39–45. [Google Scholar] [CrossRef]
- Hart, S.C.; Stark, J.M. Nitrogen limitation of the microbial biomass in an old-growth forest soil. Écoscience 2016, 4, 91–98. [Google Scholar] [CrossRef]
- Wallenstein, M.D.; McNulty, S.; Fernandez, I.J.; Boggs, J.; Schlesinger, W.H. Nitrogen fertilization decreases forest soil fungal and bacterial biomass in three long-term experiments. For. Ecol. Manag. 2006, 222, 459–468. [Google Scholar] [CrossRef]
- Eisenhauer, N.; Lanoue, A.; Strecker, T.; Scheu, S.; Steinauer, K.; Thakur, M.; Mommer, L. Root biomass and exudates link plant diversity with soil bacterial and fungal biomass. Sci. Rep. 2017, 7. [Google Scholar] [CrossRef] [Green Version]
- Papa, S.; Cembrola, E.; Pellegrino, A.; Fuggi, A.; Fioretto, A. Microbial enzyme activities, fungal biomass and quality of the litter and upper soil layer in a beech forest of south Italy. Eur. J. Soil Sci. 2013, 65. [Google Scholar] [CrossRef]
- Priha, O.; Grayston, S.J.; Hiukka, R.; Pennanen, T.; Smolander, A. Microbial community structure and characteristicsof the organic matter in soils under Pinus sylvestris, Picea abies and Betula pendula at two forest sites. Biol. Fert. Soils 2001, 33, 17–24. [Google Scholar] [CrossRef]
- Grayston, S.J.; Prescott, C.E. Microbial communities in forest floors under four tree species in coastal British Columbia. Soil Biol. Biochem. 2005, 37, 1157–1167. [Google Scholar] [CrossRef]
- Sinsabaugh, R.; Benfield, E.; Linkins, A. Cellulase Activity Associated with the Decomposition of Leaf Litter in a Woodland Stream. Oikos 1981, 36, 184. [Google Scholar] [CrossRef]
- Linkins, A.E.; Sinsabaugh, R.L.; McClaugherty, C.A.; Melillo, J.M. Comparison of cellulase activity on decomposing leaves in a hardwood forest and woodland stream. Soil Biol. Biochem. 1990, 22, 423–425. [Google Scholar] [CrossRef]
- Lu, S.; Chen, C.; Zhou, X.; Xu, Z.; Bacon, G.; Rui, Y.; Guo, X. Responses of soil dissolved organic matter to long-term plantations of three coniferous tree species. Geoderma 2012, 170, 136–143. [Google Scholar] [CrossRef]
- Aweto, A.; Moleele, N. Impact of Eucalyptus camaldulensis plantation on an alluvial soil in south eastern Botswana. Int. J. Environ. Stud. 2005, 62, 163–170. [Google Scholar] [CrossRef]
- Chung, H.; Zak, D.R.; Reich, P.B.; Ellsworth, D.S. Plant species richness, elevated CO2, and atmospheric nitrogen deposition alter soil microbial community composition and function. Glob. Chang. Biol. 2007, 13, 980–989. [Google Scholar] [CrossRef] [Green Version]
- Binkley, D.; Valentine, D. Fifty-year biogeochemical effects of green ash, white pine, and Norway spruce in a replicated experiment. For. Ecol. Manag. 1991, 40, 13–25. [Google Scholar] [CrossRef]
- Ushio, M.; Wagai, R.; Balser, T.; Kitayama, K. Variations in the soil microbial community composition of a tropical montane forest ecosystem: Does tree species matter? Soil Biol. Biochem. 2008, 40, 2699–2702. [Google Scholar] [CrossRef]
- Kourtev, P.S.; Ehrenfeld, J.G.; Huang, W.Z. Enzyme activities during litter decomposition of two exotic and two native plant species in hardwood forests of New Jersey. Soil Biol. Biochem. 2002, 34, 1207–1218. [Google Scholar] [CrossRef]
- Hu, Y.L.; Wang, S.L.; Zeng, D.H. Effects of Single Chinese Fir and Mixed Leaf Litters on Soil Chemical, Microbial Properties and Soil Enzyme Activities. Plant. Soil 2006, 282, 379–386. [Google Scholar] [CrossRef]
- Ostonen, I.; Lõhmus, K.; Helmisaari, H.-S.; Truu, J.; Meel, S. Fine root morphological adaptations in Scots pine, Norway spruce and silver birch along a latitudinal gradient in boreal forests. Tree Physiol. 2007, 27, 1627–1634. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, Y.; Zhu, B.; Sun, Z.; Zhao, C.; Yang, Y.; Piao, S. The effects of simulated nitrogen deposition on extracellular enzyme activities of litter and soil among different-aged stands of larch. J. Plant. Ecol. 2013, 7, 240–249. [Google Scholar] [CrossRef] [Green Version]
- McGill, W.B.; Cole, C.V. Comparative aspects of cycling of organic C, N, S and P through soil organic matter. Geoderma 1981, 26, 267–286. [Google Scholar] [CrossRef]
- Xu, Y.X.; Wang, Z.C.; Zhang, L.L.; Zhu, W.K.; Du, A.P. The Stoichiometric Characteristics of C, N and P in Leaf-litter-soil of Different Aged Eucalyptus urophylla×E. grandis Plantations. For. Res. 2018, 31, 168–174. [Google Scholar]
- Klose, S.; Moore, J.M.; Tabatabai, M.A. Arylsulfatase activity of microbial biomass in soils as affected by cropping systems. Biol. Fert. Soils 1999, 29, 46–54. [Google Scholar] [CrossRef]
- Vong, P.-C.; Dedourge, O.; Lasserre-Joulin, F.; Guckert, A. Immobilized-S, microbial biomass-S and soil arylsulfatase activity in the rhizosphere soil of rape and barley as affected by labile substrate C and N additions. Soil Biol. Biochem. 2003, 35, 1651–1661. [Google Scholar] [CrossRef]
- Allison, V.J.; Condron, L.M.; Peltzer, D.A.; Richardson, S.J.; Turner, B.L. Changes in enzyme activities and soil microbial community composition along carbon and nutrient gradients at the Franz Josef chronosequence, New Zealand. Soil Biol. Biochem. 2007, 39, 1770–1781. [Google Scholar] [CrossRef]


{kind=link}
| Age | Species | Age × Species | ||||
|---|---|---|---|---|---|---|
| F-Value | p-Value | F-Value | p-Value | F-Value | p-Value | |
| Fungal biomass (copy/g) | 19.046 | <0.001 | 14.191 | <0.001 | 4.471 | 0.017 |
| Bacteria biomass (copy/g) | 5.981 | 0.018 | 1.906 | 0.160 | 7.332 | 0.002 |
| F:B ratio | 25.818 | <0.001 | 25.312 | <0.001 | 23.972 | <0.001 |
| Total biomass (g/m2) | 0.821 | 0.369 | 3.746 | 0.031 | 6.249 | 0.004 |
| Cycle | Enzyme | 1-Year | 5+-Year | ||||
|---|---|---|---|---|---|---|---|
| (nmol·g−1·h−1) | EUG | EC | EP | EUG | EC | EP | |
| Carbon | β-Xylosidase | 30.46 ± 12.46 | 20.78 ± 8.99 | 12.08 ± 3.74 | 17.35 ± 4.09 | 17.83 ± 6.09 | 19.92 ± 6.68 |
| β-D-glucuronidase | 4.28 ± 1.17 | 7.79 ± 1.40 | 5.01 ± 2.09 | 4.39 ± 1.56 | 8.06 ± 1.63 | 6.46 ± 2.29 | |
| β-Cellobiosidase | 26.99 ± 12.24 | 38.83 ± 17.54 | 10.81 ± 6.75 | 18.61 ± 4.48 | 19.58 ± 4.71 | 15.19 ± 6.78 | |
| β-Glucosidase | 106.45 ± 24.98 | 160.48 ± 72.84 | 106.79 ± 35.45 | 62.83 ± 23.60 | 149.30 ± 49.30 | 88.88 ± 26.13 | |
| Nitrogen | N-Acetyl-glucosamidase | 209.01 ± 98.29 | 284.97 ± 140.50 | 178.79 ± 63.39 | 140.24 ± 77.79 | 364.46 ± 134.46 | 207.62 ± 56.58 |
| Phosphorus | Phosphatase | 439.38 ± 119.63 | 563.46 ± 186.49 | 571.10 ± 313.57 | 355.93 ± 116.23 | 581.10 ± 250.75 | 514.42 ± 196.91 |
| Sulphur | Sulfatase | 74.44 ± 28.10 | 233.75 ± 102.86 | 128.25 ± 32.54 | 180.14 ± 54.46 | 210.37 ± 78.30 | 459.97 ± 133.08 |
| Cycle | Enzyme | Age | Species | Age × Species | |||
|---|---|---|---|---|---|---|---|
| (nmol·g−1·h−1) | F-Value | p-Value | F-Value | p-Value | F-Value | p-Value | |
| Carbon | β-Xylosidase | 1.509 | 0.225 | 4.203 | 0.021 | 7.284 | 0.002 |
| β-D-glucuronidase | 4.149 | 0.047 | 7.930 | 0.001 | 8.837 | 0.001 | |
| β-Cellobiosidase | 19.672 | <0.001 | 6.118 | 0.004 | 3.395 | 0.042 | |
| β-Glucosidase | 0.171 | 0.681 | 5.091 | 0.010 | 9.745 | <0.001 | |
| Nitrogen | N-Acetyl-glucosamidase | 1.792 | 0.187 | 6.676 | 0.003 | 5.353 | 0.008 |
| Phosphorus | Phosphatase | 2.887 | 0.096 | .530 | 0.592 | 1.700 | 0.193 |
| Sulphur | Sulfatase | 15.081 | <0.001 | 19.081 | <0.001 | 12.735 | <0.001 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, J.; Liu, B.; Qu, Z.-l.; Ma, Y.; Sun, H. Age and Species of Eucalyptus Plantations Affect Soil Microbial Biomass and Enzymatic Activities. Microorganisms 2020, 8, 811. https://doi.org/10.3390/microorganisms8060811
Xu J, Liu B, Qu Z-l, Ma Y, Sun H. Age and Species of Eucalyptus Plantations Affect Soil Microbial Biomass and Enzymatic Activities. Microorganisms. 2020; 8(6):811. https://doi.org/10.3390/microorganisms8060811
Chicago/Turabian StyleXu, Jie, Bing Liu, Zhao-lei Qu, Yang Ma, and Hui Sun. 2020. "Age and Species of Eucalyptus Plantations Affect Soil Microbial Biomass and Enzymatic Activities" Microorganisms 8, no. 6: 811. https://doi.org/10.3390/microorganisms8060811
APA StyleXu, J., Liu, B., Qu, Z. -l., Ma, Y., & Sun, H. (2020). Age and Species of Eucalyptus Plantations Affect Soil Microbial Biomass and Enzymatic Activities. Microorganisms, 8(6), 811. https://doi.org/10.3390/microorganisms8060811