Genetic Diversity of Pseudomonas syringae pv. actinidiae: Seasonal and Spatial Population Dynamics

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Field Surveys
2.2. Putative Psa Isolation and Total DNA Extraction
2.3. Psa Molecular Identification and Typing
2.4. Molecular Characterization Psa Strains
2.5. Multi-Locus Sequencing Analysis(MLSA) of Psa Strains
2.6. Statistical Analysis
2.7. Nucleotide Sequence Accession Numbers
3. Results and Discussion
3.1. Psa Isolation from Portuguese Kiwifruit Orchards
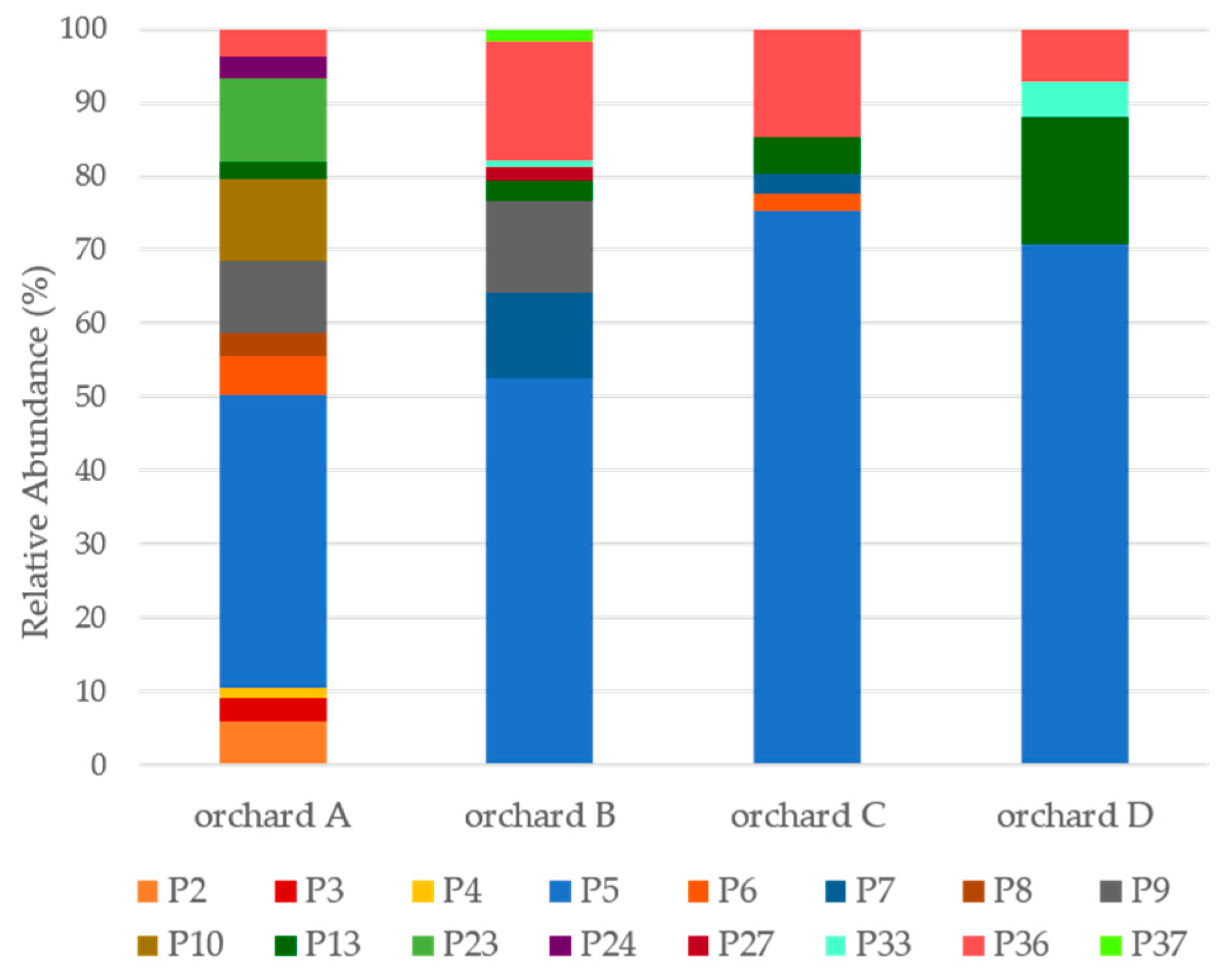
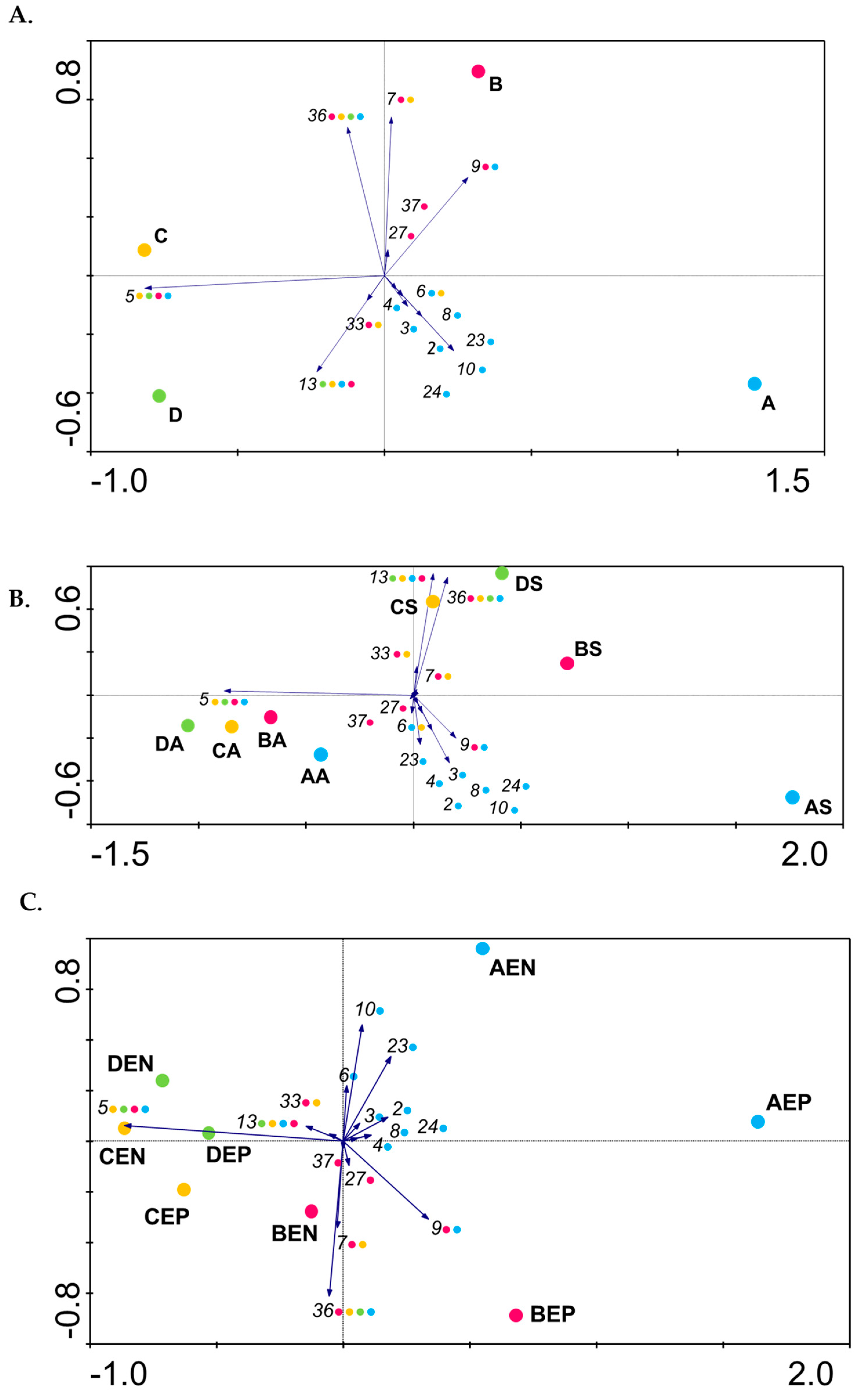
3.2. Diversity of Psa Populations in Portuguese Kiwifruit Orchards
3.3. The Diversity of Psa Populations Varies between Seasons in Each Orchard
3.4. The Diversity of Psa Populations Varies in the Phyllosphere
3.5. Psa Population Structure in Individual Plants
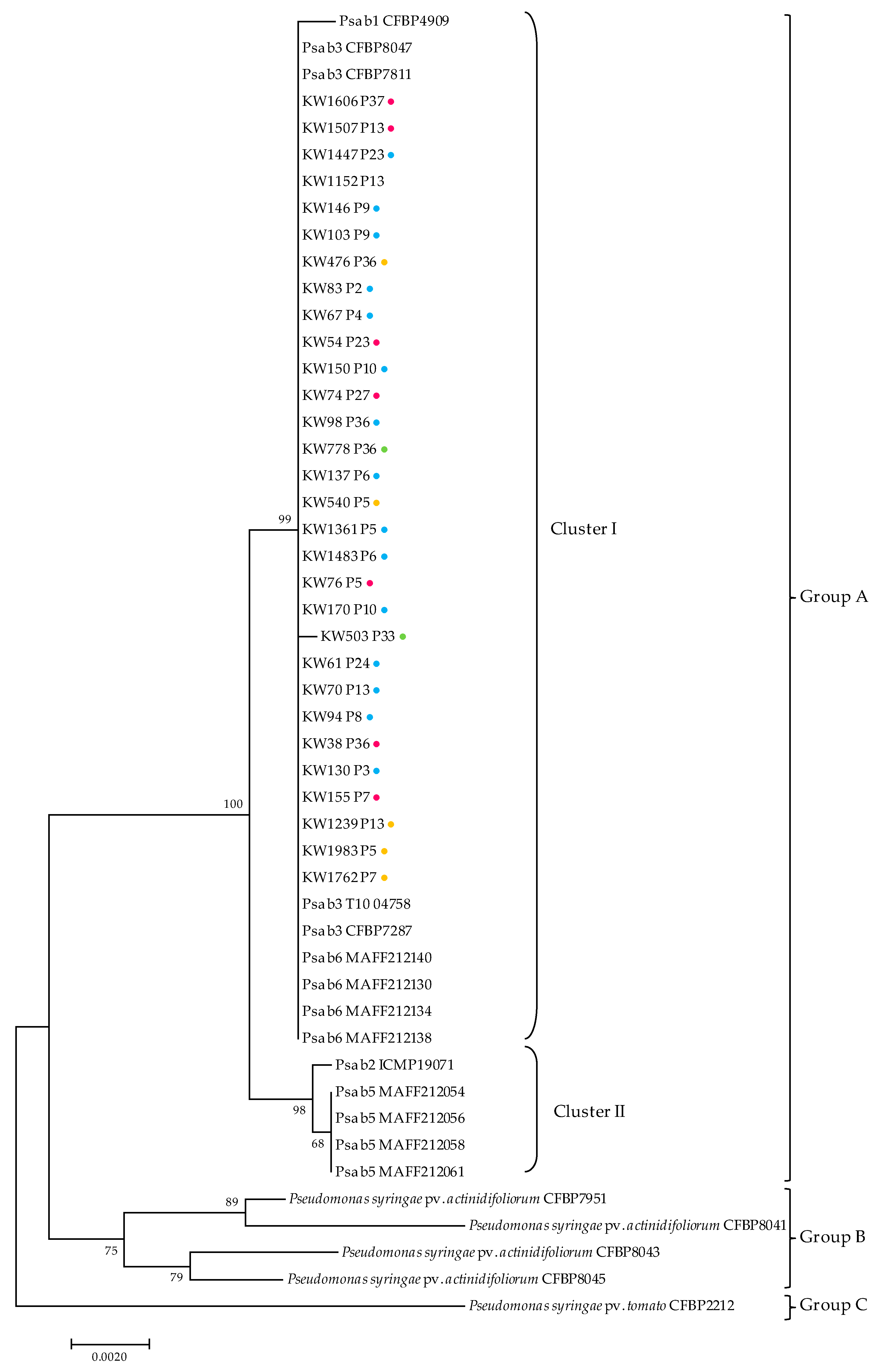
3.6. Molecular Characterization and Phylogenetic Analysis of Selected Psa Strains
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Balestra, G.M.; Mazzaglia, A.; Quattrucci, A.; Renzi, M.; Rossetti, A. Current status of bacterial canker spread on kiwifruit in Italy. Australas. Plant Dis. Notes 2009, 4, 34–36. [Google Scholar] [CrossRef]
- Scortichini, M.; Marcelletti, S.; Ferrante, P.; Petriccione, M.; Firrao, G. Pseudomonas syringae pv. actinidiae: A re-emerging, multi-faced, pandemic pathogen. Mol. Plant Pathol. 2012, 13, 1–10. [Google Scholar] [CrossRef]
- Vanneste, J. The scientific, economic, and social impacts of the New Zealand outbreak of bacterial canker of kiwifruit (Pseudomonas syringae pv. actinidiae). Annu. Rev. Phytopathol. 2017, 55, 377–399. [Google Scholar] [CrossRef] [PubMed]
- Donati, I.; Cellini, A.; Sangiorgio, D.; Vanneste, J.L.; Scortichini, M.; Balestra, G.M.; Spinelli, F. Pseudomonas syringae pv. Actinidiae: Ecology, infection dynamics and disease epidemiology. Microb. Ecol. 2020, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Takikawa, Y.; Serizawa, S.; Ichikawa, T.; Goto, M.; Takanashi, M. Pseudomonas syringae pv. actinidiae pv. nov.: The causal bacterium of canker of Kiwifruit in Japan. Ann. Phytopathol. Soc. Jpn. 1989, 55, 585–595. [Google Scholar] [CrossRef] [Green Version]
- Ferrante, P.; Scortichini, M. Molecular and phenotypic features of Pseudomonas syringae pv. actinidiae isolated during recent epidemics of bacterial canker on yellow kiwifruit (Actinidia chinensis) in central Italy. Plant Pathol. 2010, 59, 954–962. [Google Scholar] [CrossRef]
- European and Mediterranean Plant Protection Organization: EPPO A1 and A2 Lists of Pests Recommended for Regulation as Quarantine Pests. 2012. Available online: https://www.eppo.int/ACTIVITIES/plant_quarantine/A2_list (accessed on 7 August 2018).
- Donati, I.; Buriani, G.; Cellini, A.; Mauri, S.; Costa, G.; Spinelli, F. New insights on the bacterial canker of kiwifruit (Pseudomonas syringae pv. actinidiae). J. Berry Res. 2014, 4, 53–67. [Google Scholar] [CrossRef] [Green Version]
- Vanneste, J.L.; Yu, J.; Cornish, D.A.; Max, S.; Kay, C.; Onorato, R.; Spinelli, F. Recent advances in the characterization and control of Pseudomonas syringae pv. actinidiae, the causal agent of bacterial canker on Kiwifruit. Acta Hortic. 2011, 913, 443–455. [Google Scholar] [CrossRef]
- Chapman, J.R.; Taylor, R.K.; Weir, B.S.; Romberg, M.K.; Vanneste, J.L.; Luck, J.; Alexander, B.J. Phylogenetic relationships among global populations of Pseudomonas syringae pv. actinidiae. Phytopathology 2012, 102, 1034–1044. [Google Scholar] [CrossRef] [Green Version]
- Fujikawa, T.; Sawada, H. Genome analysis of the kiwifruit canker pathogen Pseudomonas syringae pv. actinidiae biovar 5. Sci. Rep. 2016, 6, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Sawada, H.; Kondo, K.; Nakaune, R. Novel biovar (biovar 6) of Pseudomonas syringae pv. actinidiae causing bacterial canker of kiwifruit (Actinidia deliciosa) in Japan. Jpn. J. Phytopathol. 2016, 82, 101–115. [Google Scholar] [CrossRef] [Green Version]
- Marceletti, S.; Ferrante, P.; Petriccione, M.; Firrao, G.; Scotichini, M. Pseudomonas syringae pv. actinidiae draft genomes comparison reveal strain-specific features involved in adaptation and virulence to actinidia species. PLoS ONE 2011, 6, e27297. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Imamura, K.; Yoneyama, K.; Kohno, Y.; Takikawa, Y.; Yamaguchi, I.; Takahashi, H. Role of phaseolotoxin by Pseudomonas syringae pv. actinidiae in the formation of halo lesions of kiwifruit canker disease. Physiol. Mol. Plant Pathol. 2002, 60, 207–214. [Google Scholar] [CrossRef]
- Koh, Y.J.; Chung, H.J.; Cha, B.J.; Lee, D.H. Outbreak and spread of bacterial canker in kiwifruit. Korean J. Plant Pathol. 1994, 10, 68–72. [Google Scholar]
- Han, H.S.; Koh, Y.J.; Hur, J.S.; Jung, J.S. Identification and characterization of coronatine-producing Pseudomonas syringae pv. actinidiae. J. Microbiol. Biotechnol. 2003, 13, 110–118. [Google Scholar]
- Vanneste, J.L. Recent progress on detecting, understanding and controlling Pseudomonas syringae pv. actinidiae: A short review. N. Z. Plant Prot. 2013, 66, 170–177. [Google Scholar] [CrossRef] [Green Version]
- McCann, H.C.; Li, L.; Liu, Y.; Li, D.; Pan, H.; Zhong, C.; Rikkerink, E.H.A.; Templeton, M.D.; Straub, C.; Colombi, E.; et al. Origin and evolution of the kiwifruit canker pandemic. Genome Boil. Evol. 2017, 9, 932–944. [Google Scholar] [CrossRef]
- Firrao, G.; Torelli, E.; Polano, C.; Ferrante, P.; Ferrini, F.; Martini, M.; Marcelletti, S.; Scortichini, M.; Ermacora, P. Genomic structural variations affecting virulence during clonal expansion of Pseudomonas syringae pv. actinidiae biovar 3 in Europe. Front. Microbiol. 2018, 9, 1–13. [Google Scholar] [CrossRef]
- Vanneste, J.L.; Yu, J.; Cornish, D.A.; Tanner, D.J.; Winder, R.; Chapman, J.R.; Taylor, R.K.; Mackay, J.F.; Dowlut, S. Identification, virulence, and distribution of two biovars of Pseudomonas syringae pv. actinidiae in New Zealand. Plant Dis. 2013, 97, 708–719. [Google Scholar] [CrossRef] [Green Version]
- Cunty, A.; Poliakoff, F.; Rivoal, C.; Cesbron, S.; Fisher-Le Saux, M.; Lemaire, C.; Jacques, M.A.; Manceau, C.; Vanneste, J.L. Characterization of Pseudomonas syringae pv. actinidiae (Psa) isolated from France and assignment of Psa biovar 4 to a de novo pathovar: Pseudomonas syringae pv. actinidifoliorum pv. nov. Plant Pathol. 2015, 64, 582–596. [Google Scholar] [CrossRef]
- Abelleira, A.; Ares, A.; Aguin, O.; Peñalver, J.; Morente, M.C.; López, M.M.; Sainz, M.J.; Mansilla, J.P. Detection and characterization of Pseudomonas syringae pv. actinidifoliorum in kiwifruit in Spain. J. Appl. Microbiol. 2015, 119, 1659–1671. [Google Scholar] [CrossRef] [PubMed]
- McCann, H.C.; Rikkerink, E.H.A.; Bertels, F.; Fiers, M.; Lu, A.; Rees-George, J.; Andersen, M.T.; Gleave, A.P.; Haubold, B.; Wohlers, M.W.; et al. Genomic analysis of the kiwifruit pathogen Pseudomonas syringae pv. actinidiae provides insight into the origins of an emergent plant disease. PLoS Pathog. 2013, 9, e1003503. [Google Scholar] [CrossRef]
- Gao, X.; Huang, Q.; Zhao, Z.; Han, Q.; Ke, X.; Qin, H.; Huang, L. Studies on the infection, colonization, and movement of Pseudomonas syringae pv. actinidiae in kiwifruit tissues using a GFPuv-Labeled strain. PLoS ONE 2016, 11, e0151169. [Google Scholar] [CrossRef]
- Xin, X.; Kvitko, B.; He, S.Y. Pseudomonas syringae: What it takes to be a pathogen. Nat. Rev. Microbiol. 2018, 16, 316–328. [Google Scholar] [CrossRef] [PubMed]
- Serizawa, S.; Ichikawa, T. Epidemiology of bacterial canker of kiwifruit. Ann. Phytopathol. Soc. Jpn. 1993, 59, 452–459. [Google Scholar] [CrossRef]
- Ferrante, P.; Fiorillo, P.; Marcelletti, S.; Marocchi, F.; Mastroleo, M.; Simeoni, S.; Scortichini, M. The importance of the main colonization and penetration sites of Pseudomonas syringae pv. actinidiae and prevailing weather conditions in the development of epidemics in yellow kiwifruit, recently observed in central Italy. J. Plant Pathol. 2012, 94, 455–461. [Google Scholar] [CrossRef]
- Hacquard, S.; Spnaepen, S.; Garrido-Oter, R.; Schulze-Lefert, P. Interplay between innate immunity and the plant microbiota. Annu. Rev. Phytopathol. 2017, 55, 565–589. [Google Scholar] [CrossRef]
- Stefani, E.; Giovanardi, D. Dissemination of Pseudomonas syringae pv. actinidiae through pollen and its epiphytic life on leaves and fruits. Phytopathol. Mediterr. 2011, 50, 489–496. [Google Scholar] [CrossRef]
- Spinelli, F.; Donati, I.; Vanneste, J.L.; Costa, M.; Costa, G. Real time monitoring of the interactions between Pseudomonas syringae pv. actinidiae and Actinidia species. Acta Hortic. 2011, 913, 461–465. [Google Scholar] [CrossRef]
- Louws, F.J.; Fulbright, D.W.; Stephens, C.T.; Bruijn, F.J. Specific genomic fingerprints of Phytopathogenic Xanthomonas and Pseudomonas pathovars and strains generated with repetitive sequences and PCR. Appl. Environ. Microbiol. 1994, 60, 2286–2295. [Google Scholar] [CrossRef] [Green Version]
- Mazzaglia, A.; Studholme, D.J.; Taratufolo, M.C.; Cai, R.; Almeida, N.F.; Goodman, T.; Guttman, D.S.; Vinatzer, B.A.; Balestra, G.M. Pseudomonas syringae pv. actinidiae (Psa) isolates from recent bacterial canker of kiwifruit outbreaks belong to the same genetic lineage. PLoS ONE 2012, 7, e36518. [Google Scholar] [CrossRef] [PubMed]
- Petriccione, M.; Zampella, L.; Mastrobuoni, F.; Scortichini, M. Occurrence of copper-resistant Pseudomonas syringae pv. syringae strains isolated from rain and kiwifruit orchards also infected by P. s. pv. actinidiae. Eur. J. Plant Pathol. 2017, 149, 953–968. [Google Scholar] [CrossRef]
- Prencipe, S.; Gullino, M.L.; Spadaro, D. Pseudomonas syringae pv. actinidiae isolated from Actinidia chinensis Var. deliciosa in Northern Italy: Genetic diversity and virulence. Eur. J. Plant Pathol. 2017, 150, 191–204. [Google Scholar] [CrossRef]
- Zhao, Z.; Chen, J.; Gao, X.; Zhang, D.; Zhang, J.; Wen, J.; Qin, H.; Guo, M.; Huang, L. Comparative genomics reveal pathogenicity-related loci in Pseudomonas syringae pv. actinidiae biovar 3. Mol. Plant Pathol. 2019, 20, 923–942. [Google Scholar] [CrossRef]
- Balestra, G.M.; Renzi, M.; Mazzaglia, A. First report of bacterial canker of Actinidia deliciosa caused by Pseudomonas syringae pv. actinidiae in Portugal. New Dis. Rep. 2010, 22, 10. [Google Scholar] [CrossRef] [Green Version]
- Renzi, M.; Mazzaglia, A.; Balestra, G.M. Widespread distribution of kiwifruit bacterial canker caused by the European Pseudomonas syringae pv. actinidiae genotype in the main production areas of Portugal. Phytopathol. Mediterr. 2012, 51, 402–409. [Google Scholar] [CrossRef]
- Moura, L.; Garcia, E.; Aguín, O.; Ares, A.; Abelleira, A.; Mansilla, P. Identification and characterization of Pseudomonas syringae pv. actinidiae (Psa) in Entre Douro e Minho region (Portugal). Rev. Ciênc. Agrár. 2015, 38, 196–205. [Google Scholar]
- Garcia, E.; Moura, L.; Abelleira, A.; Aguín, O.; Ares, A.; Mansilla, P. Characterization of Pseudomonas syringae pv. actinidiae biovar 3 on kiwifruit in north-west Portugal. J. Appl. Microbiol. 2018, 125, 1147–1161. [Google Scholar] [CrossRef]
- Eevers, N.; Gielen, M.; Sánchez-López, A.; Jaspers, S.; White, J.C.; Vangronsveld, J.; Weyens, N. Optimization of isolation and cultivation of bacterial endophytes through addition of plant extract to nutrient media. Microb. Biotechnol. 2015, 8, 707–715. [Google Scholar] [CrossRef]
- European and Mediterranean Plant Protection Organization. PM 7/120 Pseudomonas syringae pv. actinidiae. Bull 2014, 44, 360–375. [Google Scholar] [CrossRef] [Green Version]
- Koh, Y.J.; Nou, I.S. DNA markers for identification of Pseudomonas syringae pv. actinidiae. Mol. Cells. 2002, 33, 68–72. [Google Scholar]
- Gallelli, A.; L’Aurora, A.; Loreti, S. Gene sequence analysis for the molecular detection of Pseudomonas syringae pv. actinidiae: Developing diagnostic protocols. J. Plant Pathol. 2011, 93, 425–435. [Google Scholar] [CrossRef]
- Balestra, G.M.; Taratufolo, M.C.; Vinatzer, B.A.; Mazzaglia, A. A multiplex PCR assay for detection of Pseudomonas syringae pv. actinidiae and differentiation of populations with different geographic origin. Plant Dis. 2013, 97, 472–478. [Google Scholar] [CrossRef] [PubMed]
- Bereswill, S.; Bugert, P.; Völksch, B.; Ullrich, M.; Bender, C.L.; Geider, K. Identification and relatedness of coronatine-producing Pseudomonas syringae pathovars by PCR analysis and sequence determination of the amplification products. Appl. Environ. Microbiol. 1993, 60, 2921–2930. [Google Scholar] [CrossRef] [Green Version]
- Templeton, M.D.; Reinhardt, L.A.; Collyer, C.A.; Mitchell, R.E.; Cleland, W.W. Kinetic analysis of the L-Ornithine transcarbamoylase from Pseudomonas savastanoi pv. phaseolicola that is resistant to the transition state analogue (R)-Nδ-(N’-Sulfodiaminophosphinyl)-L-ornithine. Biochemistry 2005, 44, 4408–4415. [Google Scholar] [CrossRef]
- Sarkar, S.F.; Guttman, D.S. Evolution of the core genome of Pseudomonas syringae, a highly clonal, endemic plant pathogen. Jpn. J. Phytopathol. 2016, 82, 101–115. [Google Scholar] [CrossRef] [Green Version]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Higgins, D.G. CLUSTAL V: Multiple alignment of DNA and protein sequences. Methods Mol. Biol. 1994, 25, 307–318. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Boil. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Gardan, L.; Shafik, H.; Belouin, S.; Broch, R.; Grimont, F.; Grimont, P.A.D. DNA relatedness among the pathovars of Pseudomonas syringae and description of Pseudomonas tremae sp. nov. and Pseudomonas cannabina sp. nov. (ex Sutic and Dowson 1959). Int. J. Syst. Evol. Microbiol. 1999, 49, 469–478. [Google Scholar] [CrossRef] [Green Version]
- Gironde, S.; Manceau, C. Housekeeping gene sequencing and Multilocus Variable-Number Tandem-Repeat analysis to identify subpopulations within Pseudomonas syringae pv. maculicola and Pseudomonas syringae pv. tomato that correlate with host specificity. Appl. Environ. Microbiol. 2012, 78, 3266–3279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- R Core Team. R: A language and environment for statistical computing. R Found. Stat. Comput. 2020. Available online: https://www.R-project.org/ (accessed on 28 May 2020).
- Whittaker, R.H. Evolution of species diversity in land communities [Birds and vascular plants]. Evol. Biol. 1977, 10, 1–67. [Google Scholar]
- Hill, M.O. Diversity and evenness: A unifying notation and its consequences. Ecol. Soc. Am. 1973, 54, 427–432. [Google Scholar] [CrossRef] [Green Version]
- Hill, T.C.J.; Walsh, K.A.; Harris, J.A.; Moffett, B.F. Using ecological diversity measures with bacterial communities. FEMS Microbiol. Ecol. 2003, 43, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlim, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; et al. Vegan: Community Ecology Package. R Package Version 2.5-6. Available online: https://CRAN.R-project.org/package=vegan (accessed on 16 October 2019).
- Šmilauer, P.; Lepš, J. Multivariate Analysis of Ecological Data Using CANOCO 5, 2nd ed.; Cambridge University Press: Cambridge, UK, 2014. [Google Scholar] [CrossRef] [Green Version]
- Mazzaglia, A.; Renzi, M.; Balestra, M. Comparison and utilization of different PCR-based approaches for molecular typing of Pseudomonas syringae pv. actinidiae strains Italy. Can. J. Plant Pathol. 2011, 33, 8–18. [Google Scholar] [CrossRef]
- Straub, C.; Colombi, E.; Li, L.; Huang, H.; Templeton, M.D.; McCann, H.C.; Rainey, P.B. The ecological genetics of Pseudomonas syringae from kiwifruit leaves. Environ. Microbiol. 2018, 9, 2066–2084. [Google Scholar] [CrossRef]
- Sawada, H.; Miyoshi, T.; Ide, Y. Novel MLSA group (Psa5) of Pseudomonas syringae pv. actinidiae causing bacterial canker of kiwifruit (Actinidia chinensis) in Japan. Jpn. J. Phytopathol. 2014, 80, 171–184. [Google Scholar] [CrossRef] [Green Version]
- Butler, M.I.; Stockwell, P.A.; Black, M.A.; Day, R.C.; Lamont, I.L.; Poulter, R.T.M. Pseudomonas syringae pv. actinidiae from recent outbreaks of kiwifruit bacterial canker belong to different clones that originated in China. PLoS ONE 2013, 8, e57464. [Google Scholar] [CrossRef]
- Lindow, S.E. Horizontal gene transfer gone wild: Promiscuity in a kiwifruit pathogen leads to resistance to chemical control. Environ. Microbiol. 2017, 19, 1363–1365. [Google Scholar] [CrossRef]
- Monteil, C.L.; Yahara, K.; Studholme, D.J.; Mageiros, L.; Méric, G.; Swingle, B.; Morris, C.E.; Vinatzer, B.A.; Sheppard, S.K. Population-genomic insights into emergence, crop adaptation and dissemination of Pseudomonas syringae pathogens. Microb. Genom. 2016, 2, e000089. [Google Scholar] [CrossRef] [Green Version]
- Monteil, C.L.; Cai, R.; Liu, H.; Llontop, M.E.; Leman, S.; Studholme, D.J.; Morris, C.E.; Vinatzer, B.A. Nonagricultural reservoirs contribute to emergence and evolution of Pseudomonas syringae crop pathogens. New Phytol. 2013, 199, 800–811. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Orchard | Localization | Cultivar | Age (Years) | Psa Detection (Year) | Psa Disease Severity Degree 1 | Average Annual Temperature (T°) 2 | Annual Cumulative Rainfall (mm) 3 | Number of Cold Hours (h) 4 |
|---|---|---|---|---|---|---|---|---|
| A | North | Bo.Erika® | 7 | 2010 | 1 | 13.5 | 1800 | 478 |
| B | North | Hayward | 5 | 2015 | 2 | 12.5 | 1800 | 1031 |
| C | Centre | Hayward | 4 | 2015 | 3 | 14.5 | 1100 | 541 |
| D | Centre | Hayward | 30 | 2016 | 2 | 14.5 | 1100 | 440 |
| A. | ||||||
| Orchard | Index | |||||
| Dmg 1 | H’ 2 | J’ 3 | 1-D 4 | |||
| A | 2.1 | 2.0 | 0.8 | 0.8 | ||
| B | 1.5 | 1.4 | 0.7 | 0.7 | ||
| C | 0.8 | 0.8 | 0.5 | 0.4 | ||
| D | 0.6 | 0.9 | 0.6 | 0.5 | ||
| B. | ||||||
| Orchard | Season | Leaf Location | Index | |||
| Dmg 1 | H’ 2 | J’ 3 | 1-D 4 | |||
| A | spring | EP 5 | 2.4 | 2.1 | 0.9 | 0.9 |
| EN 6 | 1.5 | 1.5 | 0.8 | 0.7 | ||
| autumn | EP | 0 | 0 | 0 | 0 | |
| EN | 0.5 | 0.8 | 0.7 | 0.4 | ||
| B | spring | EP | 1.3 | 1.5 | 0.8 | 0.7 |
| EN | 1.1 | 1.4 | 0.9 | 0.7 | ||
| autumn | EP | 1.1 | 0.9 | 0.8 | 0.5 | |
| EN | 0.6 | 0.4 | 0.4 | 0.2 | ||
| C | spring | EP | 0.6 | 0.8 | 0.8 | 0.5 |
| EN | 0.7 | 1 | 0.7 | 0.6 | ||
| autumn | EP | 0.3 | 0.4 | 0.6 | 0.3 | |
| EN | 0.5 | 0.2 | 0.2 | 0.1 | ||
| D | spring | EP | 0.6 | 1 | 0.7 | 0.5 |
| EN | 0.7 | 1.2 | 0.9 | 0.7 | ||
| autumn | EP | 0 | 0 | 0 | 0 | |
| EN | 0 | 0 | 0 | 0 | ||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Figueira, D.; Garcia, E.; Ares, A.; Tiago, I.; Veríssimo, A.; Costa, J. Genetic Diversity of Pseudomonas syringae pv. actinidiae: Seasonal and Spatial Population Dynamics. Microorganisms 2020, 8, 931. https://doi.org/10.3390/microorganisms8060931
Figueira D, Garcia E, Ares A, Tiago I, Veríssimo A, Costa J. Genetic Diversity of Pseudomonas syringae pv. actinidiae: Seasonal and Spatial Population Dynamics. Microorganisms. 2020; 8(6):931. https://doi.org/10.3390/microorganisms8060931
Chicago/Turabian StyleFigueira, Daniela, Eva Garcia, Aitana Ares, Igor Tiago, António Veríssimo, and Joana Costa. 2020. "Genetic Diversity of Pseudomonas syringae pv. actinidiae: Seasonal and Spatial Population Dynamics" Microorganisms 8, no. 6: 931. https://doi.org/10.3390/microorganisms8060931
APA StyleFigueira, D., Garcia, E., Ares, A., Tiago, I., Veríssimo, A., & Costa, J. (2020). Genetic Diversity of Pseudomonas syringae pv. actinidiae: Seasonal and Spatial Population Dynamics. Microorganisms, 8(6), 931. https://doi.org/10.3390/microorganisms8060931