Removal Ability and Resistance to Cinnamic and Vanillic Acids by Fungi



 ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Fungi
2.2. Growth on Minimal Media Containing Cinnamic or Vanillic Acid
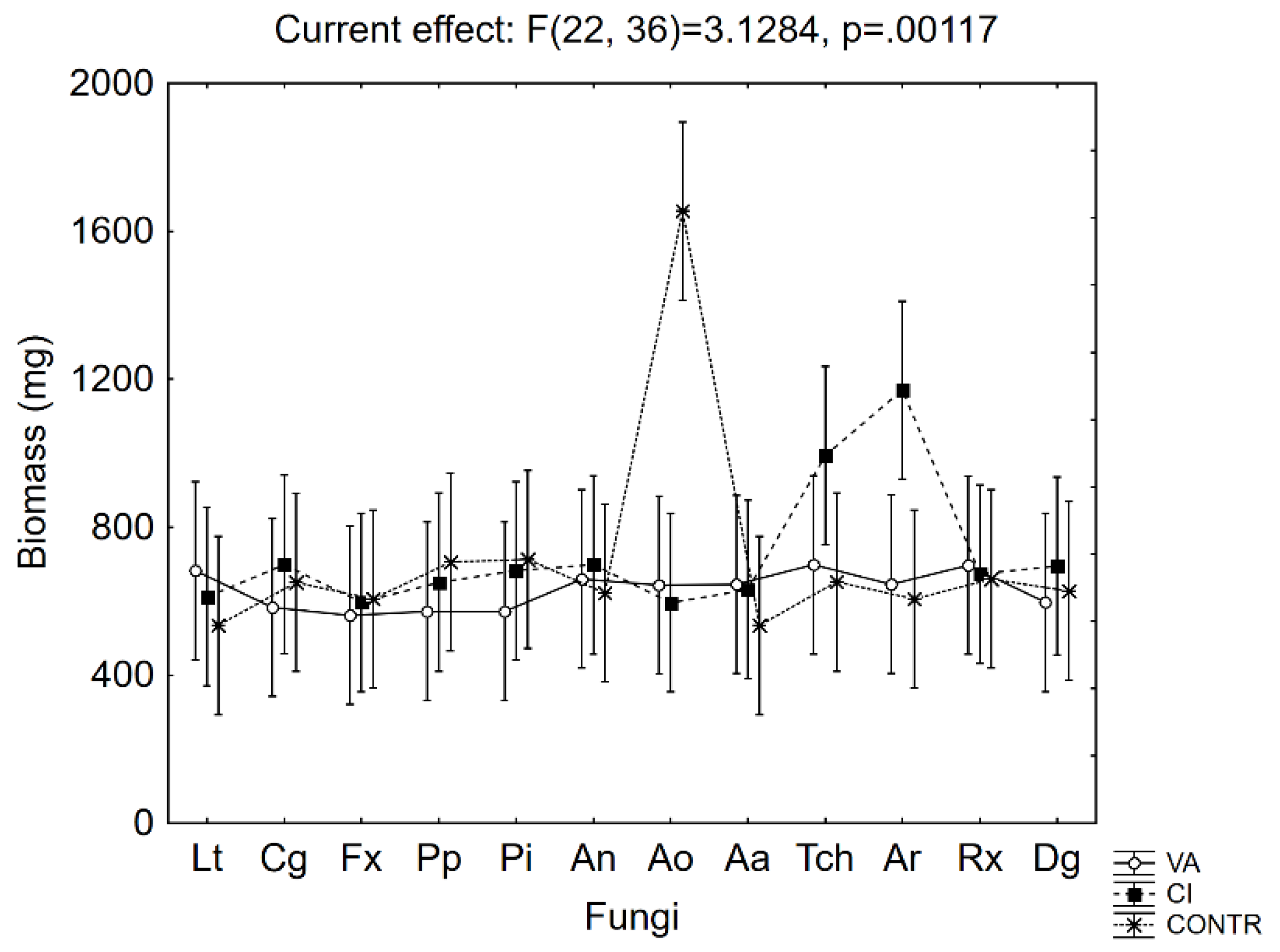
2.3. Evaluation of Fungal Biomass on Cinnamic and Vanillic Acids
2.4. Analysis of Vanillic and Cinnamic Acids
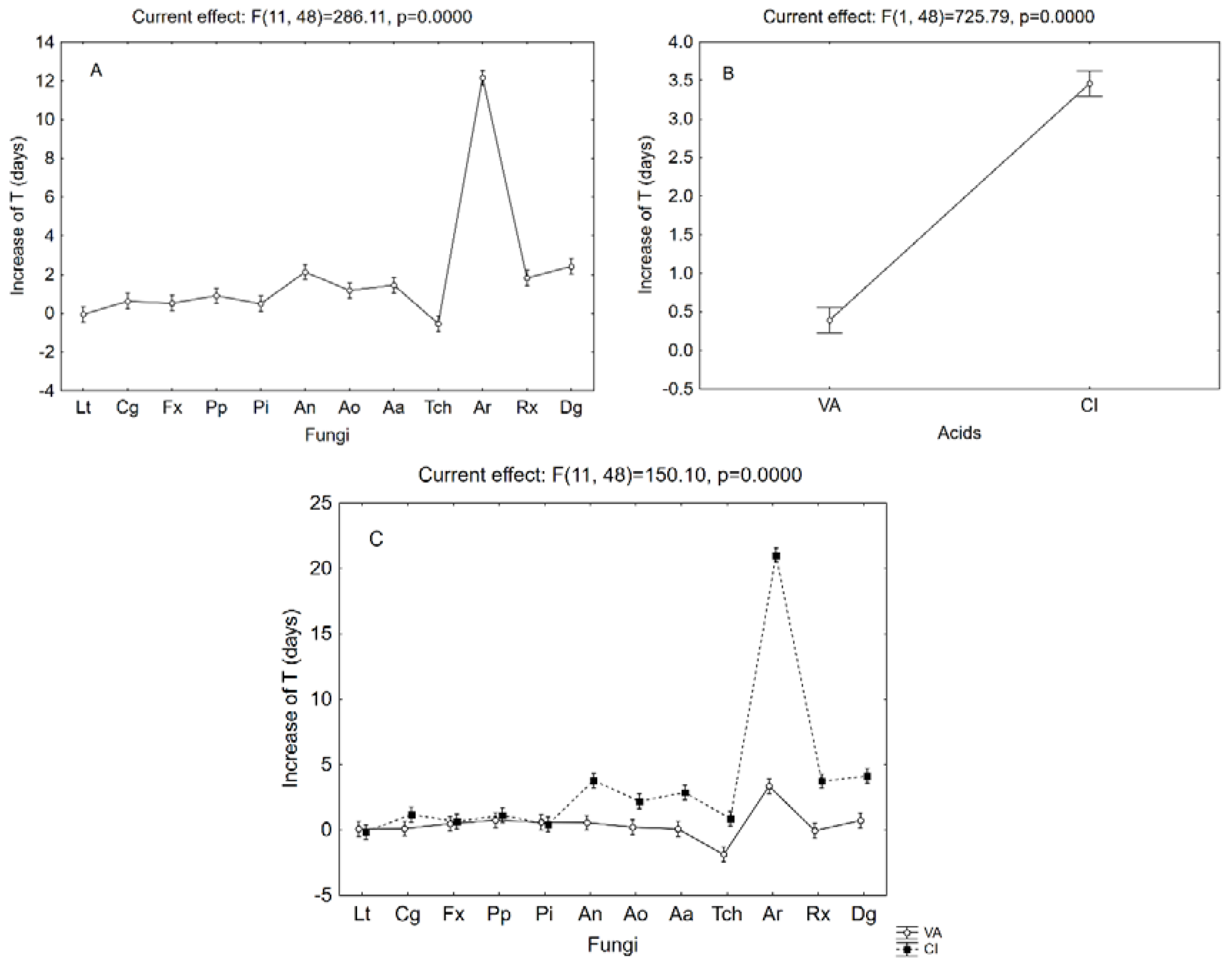
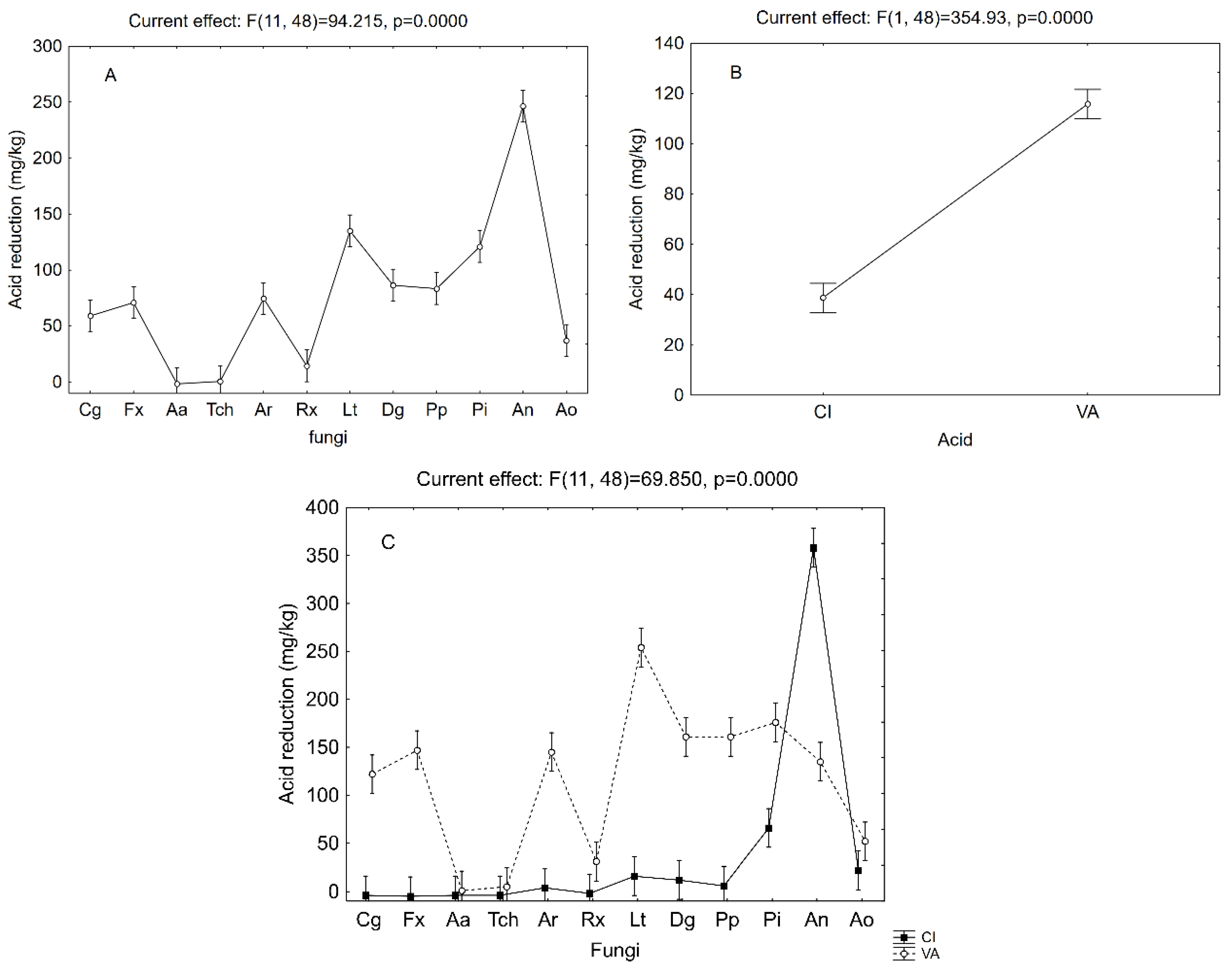
3. Results and Discussion
3.1. Growth Kinetic on Solid Media Containing Cinnamic or Vanillic Acid
3.2. Removal of Cinnamic or Vanillic Acid in Liquid Media
Author Contributions
Funding
Conflicts of Interest
References
- Morillo, J.A.; Antizar-Ladislao, B.; Monteoliva-Sánchez, M.; Ramos-Cormenzana, A.; Russell, N.J. Bioremediation and biovalorisation of olive-mill wastes. Appl. Microbiol. Biotechnol. 2009, 82, 25–39. [Google Scholar] [CrossRef] [PubMed]
- Asses, N.; Ayed, L.; Bouallagui, H.; Sayadi, S.; Hamdi, M. Biodegradation of different molecular-mass polyphenols derived from olive mill wastewaters by Geotrichum candidum. Int. Biodeterior. Biodegrad. 2009, 63, 407–413. [Google Scholar] [CrossRef]
- Mantzavinos, D.; Kalogerakis, N. Treatment of olive mill effluents. Part I. Organic matter degradation by chemical and biological processes—An overview. Environ. Int. 2005, 31, 289–295. [Google Scholar] [CrossRef] [PubMed]
- Miranda, M.A.; Galindo, F.; Amat, A.M.; Arques, A. Pyrylium salt-photosensitized degradation of phenolic contaminants derived from cinnamic acid with solar light. Correlation of the observed reactivities with fluorescence quenching. Appl. Catal. B-Environ. 2000, 28, 127–133. [Google Scholar] [CrossRef]
- Hai, F.I.; Yamamoto, K.; Nakajima, F.; Fukushi, K. Factors governing performance of continuous fungal reactor during non-sterile operation-The case of a membrane bioreactor treating textile wastewater. Chemosphere 2009, 74, 810–817. [Google Scholar] [CrossRef] [Green Version]
- Bevilacqua, A.; Cibelli, F.; Raimondo, M.L.; Carlucci, A.; Lops, F.; Sinigaglia, M.; Corbo, M.R. Fungal bioremediation of olive mill wastewater: Using a multi-step approach to model inhibition or stimulation. J. Sci. Food Agric. 2017, 97, 461–468. [Google Scholar] [CrossRef]
- Cibelli, F.; Bevilacqua, A.; Raimondo, M.L.; Campaniello, D.; Carlucci, A.; Ciccarone, C.; Sinigaglia, M.; Corbo, M.R. Evaluation of fungal growth on olive-mill wastewaters treated at high temperature and by high-pressure homogenization. Front. Microbiol. 2017, 8, 2515. [Google Scholar] [CrossRef]
- Guiraud, P.; Steiman, R.; Seigle-Murandi, F.; Benoit-Guyod, J.L. Metabolism of vanillic acid by micromycetes. World J. Microbiol. Biotechnol. 1992, 9, 270–275. [Google Scholar] [CrossRef]
- Lubbers, R.J.M.; Dilokpimol, A.; Visser, J.; Mäkelä, M.R.; Hildén, K.S.; de Vries, R.P. A comparison between the homocyclic aromatic metabolic pathways from plant-derived compounds by bacteria and fungi. Biotechnol. Adv. 2019, 37, 107396. [Google Scholar] [CrossRef]
- Lubbers, R.J.M.; Dilokpimol, A.; Navarro, J.; Peng, M.; Wang, M.; Lipzen, A.; Ng, V.; Grigoriev, I.V.; Visser, J.; Hildén, K.S.; et al. Cinnamic acid and sorbic acid conversion are mediated by the same transcriptional regulator in Aspergillus niger. Front. Bioeng. Biotechnol. 2019, 7, 249. [Google Scholar] [CrossRef]
- Dantigny, P.; Nanguy, S.P.M.; Judet-Correia, D.; Bensoussan, M. A new model for germination of fungi. Int. J. Food Microbiol. 2011, 146, 176–181. [Google Scholar] [CrossRef] [PubMed]
- Gambacorta, G.; Previtali, M.A.; Pati, S.; Baiano, A.; La Notte, E. Characterization of the phenolic profiles of some monovarietal extravirgin olive oils of Southern Italy. In Proceedings of the XXIII International Conference on Polyphenols, Winnipeg, MB, Canada, 22–25 August 2006. [Google Scholar]
- Carlucci, A.; Cibelli, F.; Lops, F.; Raimondo, M.L. Characterization of Botryosphaeriaceae species as causal agents of trunk diseases on grapevines. Plant Dis. 2015, 99, 1678–1688. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carlucci, A.; Lops, F.; Cibelli, F.; Raimondo, M.L. Phaeoacremonium species associated with olive wilt and decline in southern Italy. Eur. J. Plant Pathog. 2015, 141, 717–729. [Google Scholar] [CrossRef] [Green Version]
- Diogo, E.L.F.; Santos, J.M.; Phillips, A.J.L. Phylogeny, morphology and pathogenicity of Diaporthe and Phomopsis species on almond in Portugal. Fungal Divers. 2010, 44, 107–115. [Google Scholar] [CrossRef]
- Mostert, L.; Groenewald, J.Z.; Summerbell, R.C.; Gams, W.; Crous, P.W. Taxonomy and pathology of Togninia (Diaporthales) and its Phaeoacremonium anamorphs. Stud. Mycol. 2006, 54, 1–115. [Google Scholar] [CrossRef] [Green Version]
- Moral, J.; De La Rosa, R.; León, L.; Barranco, D.; Michailides, T.J.; Trapero, A. High susceptibility of olive cultivar FS-17 to Alternaria alternata in southern Spain. Plant Dis. 2008, 92, 1252. [Google Scholar] [CrossRef]
- Talhinhas, P.; Gonçalves, E.; Sreenivasaprasad, S.; Oliveira, H. Virulence diversity of anthracnose pathogens (Colletotrichum acutatum and C. gloeosporioides species complexes) on eight olive cultivars commonly grown in Portugal. Eur. J. Plant Pathol. 2015, 142, 73–83. [Google Scholar] [CrossRef]
- Armstrong, G.M.; Armstrong, J.K. Formae speciales and races of Fusarium oxysporum causing wilt disease. In Fusarium: Disease, Biology and Taxonomy; Nelson, P.E., Toussoun, T.A., Cook, R.J., Eds.; The Pennsylvania State University: University Park, PA, USA, 1981; pp. 392–399, 457. [Google Scholar]
- Raimondo, M.L.; Lops, F.; Carlucci, A. Phaeoacremonium italicum sp. nov., associated with esca of grapevine in southern Italy. Mycologia 2014, 106, 1119–1126. [Google Scholar] [CrossRef] [PubMed]
- Schuster, A.; Schmoll, M. Biology and biotechnology of Trichoderma. Appl. Microbiol. Biotechnol. 2010, 87, 787–799. [Google Scholar] [CrossRef] [Green Version]
- Lops, F.; Carlucci, A.; Colatruglio, L.; Frisullo, S. Uso di scarti solidi e liquidi della molitura delle olive nella lotta contro Sclerotium rolfsii. Proc. Giorn. Phytopathol. 2008, 2, 541–546. [Google Scholar]
- Carlucci, A.; Manici, L.M.; Colatruglio, L.; Caputo, A.; Frisullo, S. Rosellinia necatrix attack according to soil features in the mediterranean environment. For. Pathol. 2013, 43, 12–18. [Google Scholar] [CrossRef]
- Sharma, P. Cinnamic acid derivatives: A new chapter of various pharmacological activities. J. Chem. Pharm. Res. 2011, 3, 403–423. [Google Scholar]
- Sova, M. Antioxidant and antimicrobial activities of cinnamic acid derivatives. Mini Rev. Med. Chem. 2012, 12, 749–767. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Campbell, B.C.; Mahoney, N.E.; Chan, K.L.; Molyneux, R.J. Identification of phenolics for control of Aspergillus flavus using Saccharomyces cerevisiae in a model target-gene bioassay. J. Agric. Food Chem. 2004, 52, 7814–7821. [Google Scholar] [CrossRef]
- Said, S.; Neves, F.; Griffiths, A.J. Cinnamic acid inhibits the growth of the fungus Neurospora crassa, but is eliminated as acetophenone. Int. Biodeterior. Biodegrad. 2004, 54, 1–6. [Google Scholar] [CrossRef]
- Brennan, N.J.; Larsen, L.; Lorimer, S.D.; Perry, N.B.; Chapin, E.L.; Werk, T.L.; Henry, M.J.; Hahn, D.R. Fungicidal sesquiterpene dialdehyde cinnamates from Pseudowintera axillaris. J. Agric. Food Chem. 2006, 54, 468–473. [Google Scholar] [CrossRef]
- Lattanzio, V.; Lattanzio, V.M.T.; Cardinali, A.; Amendola, V. Role of phenolics in the resistance mechanisms of plants against fungal pathogens and insects. In Phytochemistry: Advances in Research; Imperato, F., Ed.; Research Signpost: Kerala, India, 2006; pp. 23–67. [Google Scholar]
- Bisogno, F.; Mascoti, L.; Sanchez, C.; Garibotto, F.; Giannini, F.; Kurina-Sanz, M.; Enriz, R. Structure antifungal activity relationship of cinnamic acid derivatives. J. Agric. Food Chem. 2007, 55, 10635–10640. [Google Scholar] [CrossRef]
- Korošec, N.; Soca, A.; Turk, N.; Novak, M.; Lah, L.; Stojan, J.; Podobnik, B.; Berne, S.; Zupanec, N.; Bunc, M.; et al. Antifungal activity of cinnamic acid derivatives involves inhibition of benzoate 4-hydrosylase (CYP53). J. Appl. Microbiol. 2014, 116, 955–966. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.Q.G.; Li, B.; Tian, S. Effect of cinnamic acid for controlling gray mold on table grape and its possible mechanisms of action. Curr. Microbiol. 2015, 71, 396–402. [Google Scholar] [CrossRef]
- Roy, S.; Nuckles, E.; Archbold, D.D. Effects of phenolic compounds on growth of Colletotrichum spp. in vitro. Curr. Microbiol. 2018, 75, 550–556. [Google Scholar] [CrossRef]
- Bushwell, J.A.; Eiksson, K.E.; Gupta, J.K.; Hamp, S.G.; Nordh, I. Vanillic acid metabolism by selected soft-rot, brown-rot, and white rote fungi. Arch. Microbiol. 1982, 131, 366–374. [Google Scholar] [CrossRef]
- Salgado, J.M.; Abrunhosa, L.; Venãncio, A.; Domíniguez, J.M.; Belo, I. Combined bioremediation and enzyme production by Aspergillus sp. in olive mill and winery wastewaters. Int. J. Biodeterior. Biodegrad. 2016, 110, 16–23. [Google Scholar] [CrossRef] [Green Version]
- McNamara, C.J.; Anastasiou, C.C.; O’Flaherty, V.; Mitchell, R. Bioremediation of olive mill wastewater. Int. J. Biodeterior. Biodegrad. 2008, 61, 127–134. [Google Scholar] [CrossRef]
- Aissam, H.; Penninckx, M.J.; Benlemlih, M. Reduction of phenolics content and COD in olive oil mill wastewaters by indigenous yeasts and fungi. World J. Microb. Biotechnol. 2007, 23, 1203–1208. [Google Scholar] [CrossRef]
- Vieira, J.C.B.; Câmara, M.P.S.; Bezerra, J.D.P.; Motta, C.M.S.; Machado, A.R. First report of Lasiodiplodia theobromae causing rot in eggplant fruit in Brazil. Dis. Notes 2018, 102, 10. [Google Scholar] [CrossRef] [PubMed]
- Venkatesagowda, B.; Ponugupaty, E.; Barbosa-Dekker, A.M.; Dekker, R.F.H. The purification and characterization of lipases from Lasiodiplodia theobromae, and their immobilization and use for biodiesel production from coconut oil. Appl. Biochem. Biotecnol. 2018, 185, 619–640. [Google Scholar] [CrossRef]
- Clifford, D.R.; Faulkner, J.K.; Walker, J.R.L.; Woodcock, D. Metabolism of cinnamic acid by Aspergillus niger. Phytochemistry 1969, 8, 549–552. [Google Scholar] [CrossRef]
- Pinches, S.E.; Apps, P. Production in food of 1,3-pentadiene and styrene by Trichoderma species. Int. J. Food Microbiol. 2007, 116, 182–185. [Google Scholar] [CrossRef]
- Lafeuille, J.L.; Buniak, M.L.; Vioujas, M.C.; Lefevre, S. Natural formation of styrene by cinnamon mold flora. J. Food Sci. 2009, 74, M276–M283. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Fungal Species | Host | Locality | Identification Tools | Reference or Collection | Abbreviation |
|---|---|---|---|---|---|
| Lasiodiplodia theobromae | Vitis vinifera | Cerignola, Foggia, Italy | MC | [13] | Lt |
| Colletotrichum gloeosporioides | Olea europea | Matino, Lecce, Italy | MOR | Dept.SAFE | Cg |
| Fusarium oxysporum | Olea europea | Presicce, Lecce, Italy | MC | Dept.SAFE | Fx |
| Phaeoacremonium parasiticum | Olea europea | Cerignola, Foggia, Italy | MC | [14] | Pp |
| Penicillium italicum | Soil | Stornarella, Italy | MOR | Dept.SAFE | Pi |
| Aspergillus niger | Oil-mill wastewater | Cerignola, Foggia, Italy | MOR | Dept.SAFE | An |
| Aspergillus ochraceus | Soil | Stornarella, Foggia, Italy | MOR | Dept.SAFE | Ao |
| Alternaria alternata | Olea europea | Cerignola, Foggia, Italy | MOR | [14] | Aa |
| Trichoderma sp. | Soil | Stornarella, Foggia, Italy | MOR | Dept.SAFE | Tch |
| Athelia rolfsii | Solanum lycopersicum | Stornarella, Foggia, Italy | MOR | Dept.SAFE | Ar |
| Rosellinia necatrix | Olea europea | Cerignola, Foggia, Italy | MOR | Dept.SAFE | Rx |
| Diaporthe amygdali | Prunus dulcis | Matino, Lecce, Italy | MC | Dept.SAFE | Dg |
| Code | Species | Control | Vanillic Acid | Cinnamic Acid |
|---|---|---|---|---|
| Lt | Lasiodiplodia theobromae | 3.31 ± 0.07A | 3.38 ± 0.15A | 3.14 ± 0.08A |
| Cg | Colletotrichum gleosporioides | 3.85 ± 0.09A | 3.93 ± 0.08A | 5.01 ± 0.07B |
| Fx | Fusarium oxysporum | 2.70 ± 0.03A | 3.15 ± 0.10A | 3.32 ± 0.08A |
| Pp | Phaeoacremonium parasiticum | 8.80 ± 0.25A | 9.52 ± 0.28A | 9.88 ± 0.48A |
| Pi | Penicillium italicum | 6.22 ± 0.26A | 6.79 ± 0.82A | 6.63 ± 0.89A |
| An | Aspergillus niger | 4.38 ± 0.08A | 4.90 ± 0.12A | 8.11 ± 1.11B |
| Ao | Aspergillus ochraceus | 7.34 ± 0.35A | 7.54 ± 0.59A | 9.50 ± 1.11B |
| Aa | Alternaria alternata | 3.05 ± 0.01A | 3.12 ± 0.02A | 5.88 ± 0.17B |
| Tch | Trichoderma sp. | 3.00 ± 0.01B | 1.10 ± 1.00A | 3.94 ± 0.10C |
| Ar | Athelia rolfsii | 3.00 ± 0.01A | 6.31 ± 0.16B | - * |
| Rx | Rosellinia necatrix | 4.17 ± 0.09A | 4.09 ± 0.08A | 7.89 ± 0.27B |
| Dg | Diaporthe amygdali | 4.10 ± 0.10A | 4.80 ± 0.12A | 6.60 ± 0.49B |
| Ctr | Ctr-21 | Cg | Fx | Aa | Tch | Ar | Rx | Lt | Dg | Pp | Pi | An | Ao |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Cinnamic acid | |||||||||||||
| 351 ± 7 A | 359 ± 10 A, d | 363 ± 11 d | 364 ± 2 d | 363 ± 1 d | 363 ± 1 d | 355 d | 361 ± 1 d | 343 ± 7 c | 347 ± 1 c | 353 ± 2 d | 293 ± 23 b | 1 a | 337 ± 1 c |
| Vanillic acid | |||||||||||||
| 359 ± 31 A | 362 ± 5 A, e | 240 ± 4 b | 215 ± 30 b | 361 ± 1 e | 357 ± 5 e | 217 ± 9 b | 331 ± 13 d | 108 ± 16 a | 201 ± 59 b | 201 ± 11 b | 186 ± 25 b | 227 ± 51 b | 310 ± 8 c |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Speranza, B.; Cibelli, F.; Baiano, A.; Carlucci, A.; Raimondo, M.L.; Campaniello, D.; Viggiani, I.; Bevilacqua, A.; Rosaria Corbo, M. Removal Ability and Resistance to Cinnamic and Vanillic Acids by Fungi. Microorganisms 2020, 8, 930. https://doi.org/10.3390/microorganisms8060930
Speranza B, Cibelli F, Baiano A, Carlucci A, Raimondo ML, Campaniello D, Viggiani I, Bevilacqua A, Rosaria Corbo M. Removal Ability and Resistance to Cinnamic and Vanillic Acids by Fungi. Microorganisms. 2020; 8(6):930. https://doi.org/10.3390/microorganisms8060930
Chicago/Turabian StyleSperanza, Barbara, Francesca Cibelli, Antonietta Baiano, Antonia Carlucci, Maria Luisa Raimondo, Daniela Campaniello, Ilaria Viggiani, Antonio Bevilacqua, and Maria Rosaria Corbo. 2020. "Removal Ability and Resistance to Cinnamic and Vanillic Acids by Fungi" Microorganisms 8, no. 6: 930. https://doi.org/10.3390/microorganisms8060930
APA StyleSperanza, B., Cibelli, F., Baiano, A., Carlucci, A., Raimondo, M. L., Campaniello, D., Viggiani, I., Bevilacqua, A., & Rosaria Corbo, M. (2020). Removal Ability and Resistance to Cinnamic and Vanillic Acids by Fungi. Microorganisms, 8(6), 930. https://doi.org/10.3390/microorganisms8060930