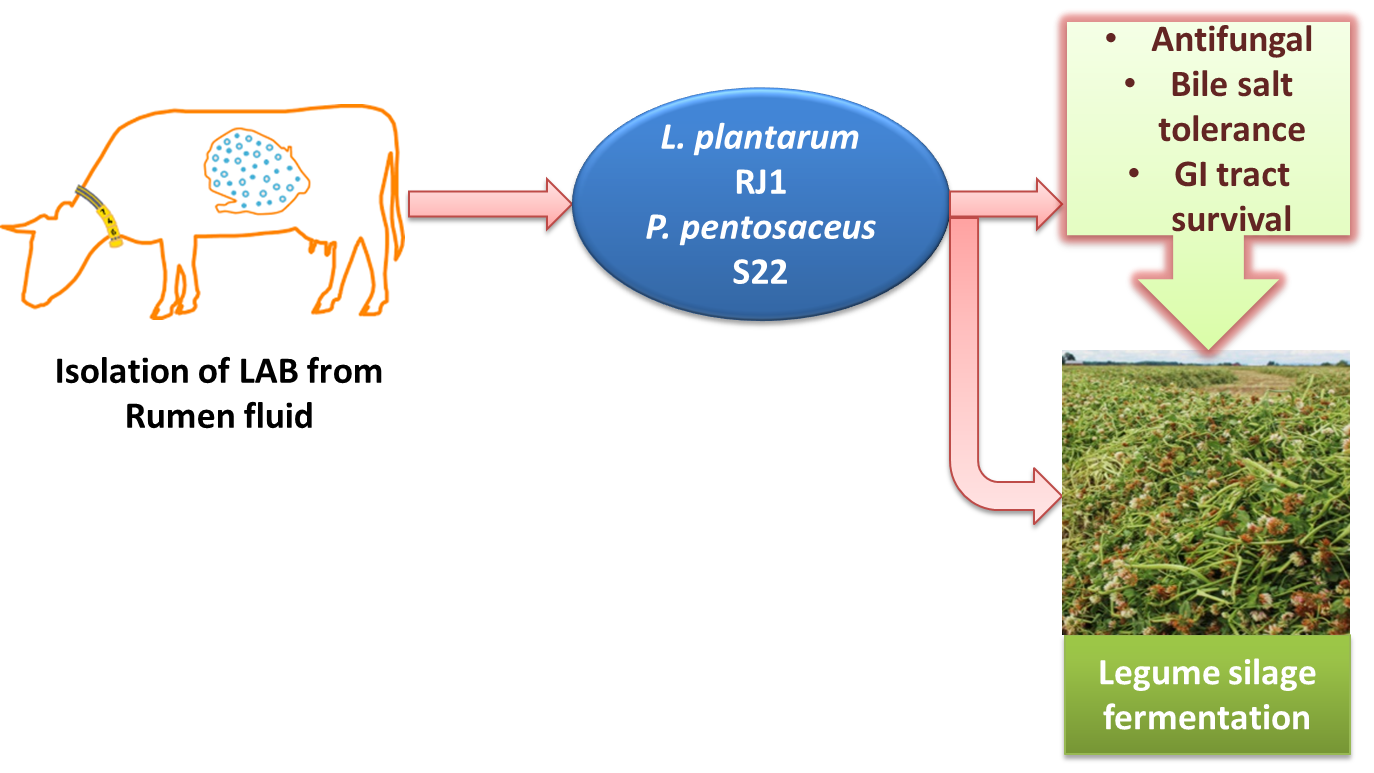
Low-Carbohydrate Tolerant LAB Strains Identified from Rumen Fluid: Investigation of Probiotic Activity and Legume Silage Fermentation



Abstract
:
1. Introduction
2. Materials and Methods
2.1. Rumen Fluid for LAB Isolation
2.2. Isolation of LAB Cultures
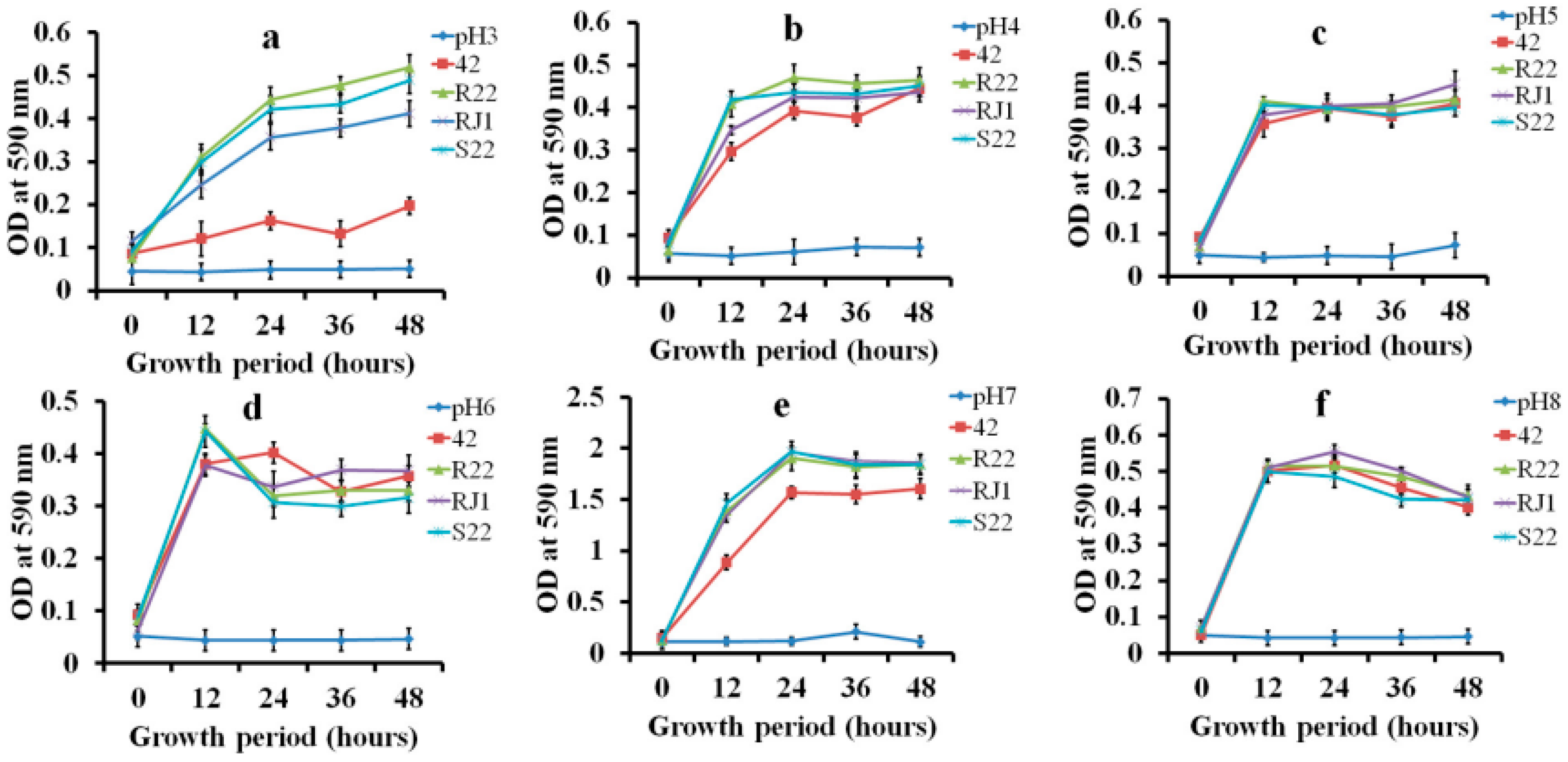
2.3. LAB Growth Pattern Analysis in Minimal Carbohydrate Medium
2.4. Molecular Characterization by 16S-rRNA Gene Sequencing
2.5. Biochemical Characterization of Isoalted LAB
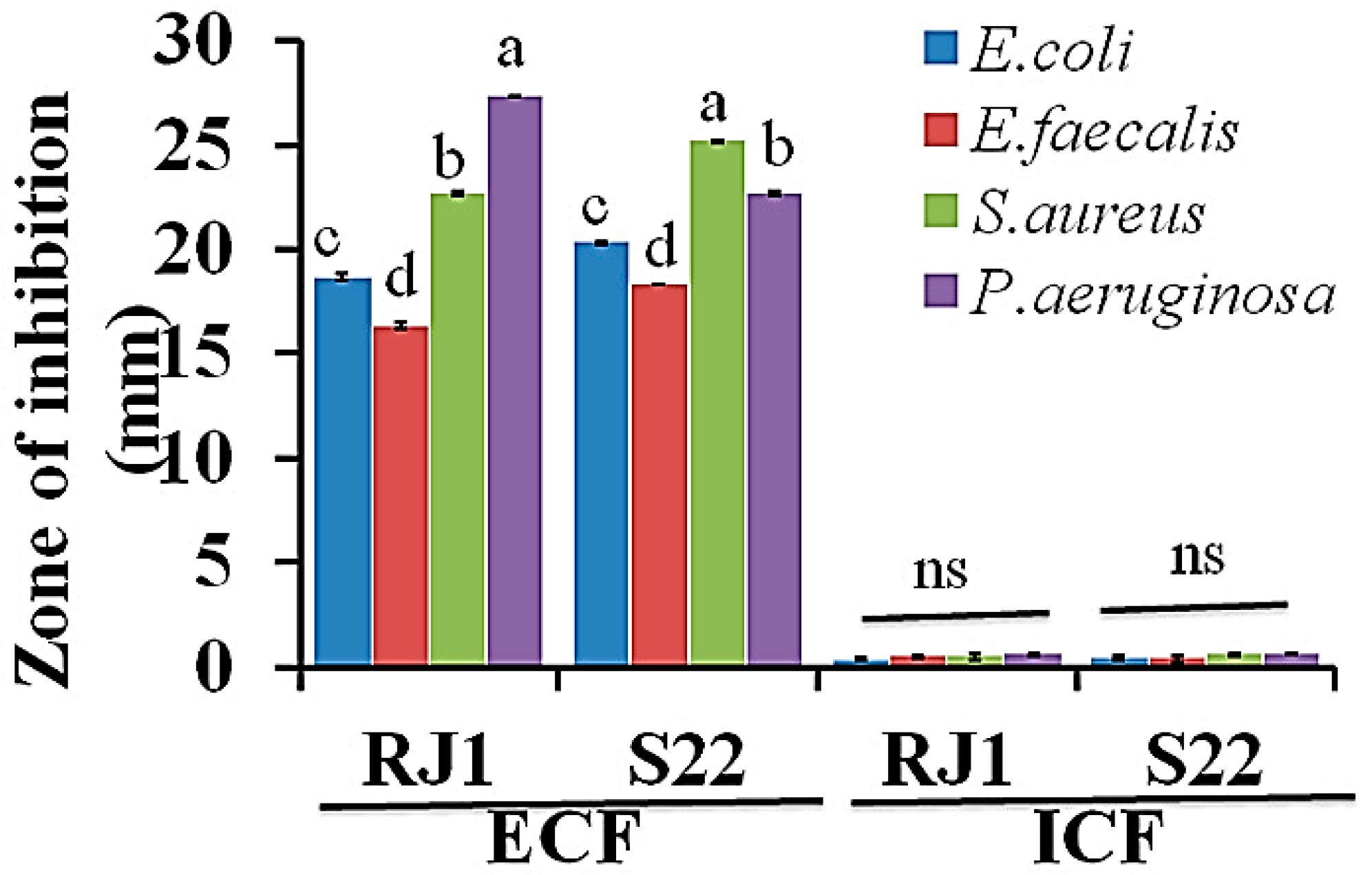
2.6. Antibacterial Assay of Isolates
2.7. Antifingal Activity of LAB Isolates by Agar Spot Method
2.8. Antioxidant Activity of Isolates
2.9. Probiotic Characterization of LAB Isolates
2.9.1. Bile Salt Tolerance Assay
2.9.2. Simulated Gastric and Intestinal Fluid Tolerance Tests
2.9.3. Cell Surface Hydrophobicity, Auto-Aggregation, and Co-Aggregation Properties of LAB Strains
2.10. Proteolytic Activity of LAB Cultures
2.11. In Vitro Legume Silage Production
2.11.1. Legumes Silage Preparation
2.11.2. pH Analysis of Fermented Silage
2.11.3. Total Microbial Population Analysis in Legume Silage
2.11.4. Organic Acid Analysis Using HPLC-RID
2.12. Statistical Analysis
3. Results
3.1. Isolation and Selection of LAB from Rumen Fluid
3.2. Growth Pattern Analysis in Low-Carbohydrate Medium
3.3. Molecular Characterization by 16S-rRNA Gene Sequencing
3.4. Biochemical Properties of Isolates
3.5. Antibacterial Activity of Isolates
3.6. Antifungal Properties of Isolates
3.7. Antioxidant Activity of Isolates
3.8. Probiotic Characterization of LAB Isolates
3.8.1. LAB Tolerance to Simulated Gastric and Intestinal Conditions
3.8.2. Bile Salt Tolerance of Isolates
3.9. Proteolytic Activity
3.10. Cell Surface Hydrophobicity and Aggregation Properties of RJ1 and S22
3.11. Experimental Study of Silage Fermentation by LAB Isolates
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Lee, K.W.; Park, J.Y.; Sa, H.D.; Jeong, J.H.; Jin, D.E.; Heo, H.J.; Kim, J.H. Probiotic properties of Pediococcus strains isolated from jeotgals, salted and fermented Korean sea-food. Anaerobe 2014, 28, 199–206. [Google Scholar] [CrossRef]
- Son, S.-H.; Jeon, H.-L.; Jeon, E.B.; Lee, N.-K.; Park, Y.-S.; Kang, D.-K.; Paik, H.-D. Potential probiotic Lactobacillus plantarum Ln4 from kimchi: Evaluation of β-galactosidase and antioxidant activities. LWT—Food Sci. Technol. 2017, 85, 181–186. [Google Scholar] [CrossRef]
- Kim, J.A.; Bayo, J.; Cha, J.; Choi, Y.J.; Jung, M.Y.; Kim, D.H.; Kim, Y. Investigating the probiotic characteristics of four microbial strains with potential application in feed industry. PLoS ONE 2019, 14, e0218922. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Timmerman, H.M.; Veldman, A.; van den Elsen, E.; Rombouts, F.M.; Beynen, A.C. Mortality and growth performance of broilers given drinking water supplemented with chicken-specific probiotics. Poult. Sci. 2006, 85, 1383–1388. [Google Scholar] [CrossRef] [PubMed]
- Szumacher-Strabel, M.; Stochmal, A.; Cieslak, A.; Kozlowska, M.; Kuznicki, D.; Kowalczyk, M.; Oleszek, W. Structural and quantitative changes of saponins in fresh alfalfa compared to alfalfa silage. J. Sci. Food Agric. 2019, 99, 2243–2250. [Google Scholar] [CrossRef]
- Fijalkowska, M.; Przemieniecki, S.W.; Purwin, C.; Lipinski, K.; Kurowski, T.P.; Karwowska, A. The effect of an additive containing three Lactobacillus species on the fermentation pattern and microbiological status of silage. J. Sci. Food Agric. 2020, 100, 1174–1184. [Google Scholar] [CrossRef]
- Li, D.; Wang, Y.; Zhang, Y.; Lin, Y.; Yang, F. Evaluation of lactic acid bacteria isolated from alfalfa for silage fermentation. Grassl. Sci. 2018, 64, 190–198. [Google Scholar] [CrossRef]
- Zheng, J.; Wittouck, S.; Salvetti, E.; Franz, C.; Harris, H.M.B.; Mattarelli, P.; O’Toole, P.W.; Pot, B.; Vandamme, P.; Walter, J.; et al. A taxonomic note on the genus Lactobacillus: Description of 23 novel genera, emended description of the genus Lactobacillus Beijerinck 1901, and union of Lactobacillaceae and Leuconostocaceae. Int. J. Syst. Evol. Microbiol. 2020, 70, 2782–2858. [Google Scholar] [CrossRef]
- Martin, C.; Copani, G.; Niderkorn, V. Imapcts oof forage legumes on intake, digestion and methane emissions in ruminants. Legume Perspect. 2016, 12, 24–25. [Google Scholar]
- Jatkauskas, J.; Vrotniakiene, V.; Lanckriet, A. The effect of different types of inoculants on the characteristics of alfalfa, ryegrass and red clover/ryegrass/timothy silage. Zemdirb.-Agric. 2015, 102, 95–102. [Google Scholar] [CrossRef] [Green Version]
- Kung, L., Jr.; Shaver, R.D.; Grant, R.J.; Schmidt, R.J. Silage review: Interpretation of chemical, microbial, and organoleptic components of silages. J. Dairy Sci. 2018, 101, 4020–4033. [Google Scholar] [CrossRef] [PubMed]
- Mulaw, G.; Sisay Tessema, T.; Muleta, D.; Tesfaye, A. In Vitro Evaluation of Probiotic Properties of Lactic Acid Bacteria Isolated from Some Traditionally Fermented Ethiopian Food Products. Int. J. Microbiol. 2019, 2019, 7179514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uher, D.; Konjačić, M.; Jareš, D.; Maćešić, D. The effect of bacterial inoculant on chemical composition and fermentation of alfalfa silage. J. Cent. Eur. Agric. 2019, 20, 657–664. [Google Scholar] [CrossRef]
- Wang, J.; Wang, J.Q.; Zhou, H.; Feng, T. Effects of addition of previously fermented juice prepared from alfalfa on fermentation quality and protein degradation of alfalfa silage. Anim. Feed Sci. Technol. 2009, 151, 280–290. [Google Scholar] [CrossRef]
- Broderick, G.A.; Stevenson, M.J.; Patton, R.A.; Lobos, N.E.; Olmos Colmenero, J.J. Effect of supplementing rumen-protected methionine on production and nitrogen excretion in lactating dairy cows. J. Dairy Sci. 2008, 91, 1092–1102. [Google Scholar] [CrossRef] [Green Version]
- Guo, L.; Yao, D.; Li, D.; Lin, Y.; Bureenok, S.; Ni, K.; Yang, F. Effects of Lactic Acid Bacteria Isolated From Rumen Fluid and Feces of Dairy Cows on Fermentation Quality, Microbial Community, and in vitro Digestibility of Alfalfa Silage. Front. Microbiol. 2019, 10, 2998. [Google Scholar] [CrossRef]
- Kim, J.G.; Ham, J.S.; Li, Y.W.; Park, H.S.; Huh, C.S.; Park, B.C. Development of a new lactic acid bacterial inoculant for fresh rice straw silage. Asian-Australas J. Anim. Sci. 2017, 30, 950–956. [Google Scholar] [CrossRef]
- Yi, L.; Dang, J.; Zhang, L.; Wu, Y.; Liu, B.; Lü, X. Purification, characterization and bactericidal mechanism of a broad spectrum bacteriocin with antimicrobial activity against multidrug-resistant strains produced by Lactobacillus coryniformis XN8. Food Control 2016, 67, 53–62. [Google Scholar] [CrossRef] [Green Version]
- Ogunade, I.M.; Kim, D.H.; Jiang, Y.; Weinberg, Z.G.; Jeong, K.C.; Adesogan, A.T. Control of Escherichia coli O157:H7 in contaminated alfalfa silage: Effects of silage additives. J. Dairy Sci. 2016, 99, 4427–4436. [Google Scholar] [CrossRef]
- Arasu, M.V.; Jung, M.W.; Ilavenil, S.; Kim, D.H.; Park, H.S.; Park, J.W.; Al-Dhabi, N.A.; Choi, K.C. Characterization, phylogenetic affiliation and probiotic properties of high cell density Lactobacillus strains recovered from silage. J. Sci. Food Agric. 2014, 94, 2429–2440. [Google Scholar] [CrossRef]
- Das, D.; Goyal, A. Antioxidant activity and γ-aminobutyric acid (GABA) producing ability of probiotic Lactobacillus plantarum DM5 isolated from Marcha of Sikkim. LWT—Food Sci. Technol. 2015, 61, 263–268. [Google Scholar] [CrossRef]
- Ding, W.; Wang, L.; Zhang, J.; Ke, W.; Zhou, J.; Zhu, J.; Guo, X.; Long, R. Characterization of antioxidant properties of lactic acid bacteria isolated from spontaneously fermented yak milk in the Tibetan Plateau. J. Funct. Foods 2017, 35, 481–488. [Google Scholar] [CrossRef]
- Huang, Y.; Adams, M.C. In vitro assessment of the upper gastrointestinal tolerance of potential probiotic dairy propionibacteria. Int. J. Food Microbiol. 2004, 91, 253–260. [Google Scholar] [CrossRef] [PubMed]
- Prabhurajeshwar, C.; Chandrakanth, R.K. Probiotic potential of Lactobacilli with antagonistic activity against pathogenic strains: An in vitro validation for the production of inhibitory substances. Biomed. J. 2017, 40, 270–283. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Wang, Y.; Cui, H.; Li, Y.; Sun, Y.; Qiu, H.J. Characterization of Lactic Acid Bacteria Isolated From the Gastrointestinal Tract of a Wild Boar as Potential Probiotics. Front. Vet. Sci. 2020, 7, 49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ilavenil, S.; Vijayakumar, M.; Kim, D.H.; Valan Arasu, M.; Park, H.S.; Ravikumar, S.; Choi, K.C. Assessment of probiotic, antifungal and cholesterol lowering properties of Pediococcus pentosaceus KCC-23 isolated from Italian ryegrass. J. Sci. Food Agric. 2016, 96, 593–601. [Google Scholar] [CrossRef] [PubMed]
- TY, T. Isolation and Purification of Proteolytic Enzyme Produced by Lactic Acid Bacteria from Budu and Bambangan. Master’s Thesis, Universiti Putra Malaysia, Serdang, Selangor, Malaysia, 2012. [Google Scholar]
- Bao, W.; Mi, Z.; Xu, H.; Zheng, Y.; Kwok, L.Y.; Zhang, H.; Zhang, W. Assessing quality of Medicago sativa silage by monitoring bacterial composition with single molecule, real-time sequencing technology and various physiological parameters. Sci. Rep. 2016, 6, 28358. [Google Scholar] [CrossRef]
- Soundharrajan, I.; Kim, D.; Kuppusamy, P.; Muthusamy, K.; Lee, H.J.; Choi, K.C. Probiotic and Triticale Silage Fermentation Potential of Pediococcus pentosaceus and Lactobacillus brevis and Their Impacts on Pathogenic Bacteria. Microorganisms 2019, 7, 318. [Google Scholar] [CrossRef] [Green Version]
- Zhang, F.; Wang, X.; Lu, W.; Li, F.; Ma, C. Improved Quality of Corn Silage When Combining Cellulose-Decomposing Bacteria and Lactobacillus buchneri during Silage Fermentation. Biomed. Res. Int. 2019, 2019, 4361358. [Google Scholar] [CrossRef] [Green Version]
- Kuley, E.; Ozyurt, G.; Ozogul, I.; Boga, M.; Akyol, I.; Rocha, J.M.; Ozogul, F. The Role of Selected Lactic Acid Bacteria on Organic Acid Accumulation during Wet and Spray-Dried Fish-based Silages. Contributions to the Winning Combination of Microbial Food Safety and Environmental Sustainability. Microorganisms 2020, 8, 172. [Google Scholar] [CrossRef] [Green Version]
- Charalampopoulos, D.; Pandiella, S.S.; Webb, C. Growth studies of potentially probiotic lactic acid bacteria in cereal-based substrates. J. Appl. Microbiol. 2002, 92, 851–859. [Google Scholar] [CrossRef] [PubMed]
- Jin, J.; Zhang, B.; Guo, H.; Cui, J.; Jiang, L.; Song, S.; Sun, M.; Ren, F. Mechanism analysis of acid tolerance response of bifidobacterium longum subsp. longum BBMN 68 by gene expression profile using RNA-sequencing. PLoS ONE 2012, 7, e50777. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gavrilova, E.; Anisimova, E.; Gabdelkhadieva, A.; Nikitina, E.; Vafina, A.; Yarullina, D.; Bogachev, M.; Kayumov, A. Newly isolated lactic acid bacteria from silage targeting biofilms of foodborne pathogens during milk fermentation. BMC Microbiol. 2019, 19, 248. [Google Scholar] [CrossRef] [PubMed]
- Tilahun, B.; Tesfaye, A.; Muleta, D.; Bahiru, A.; Terefework, Z.; Wessel, G. Isolation and Molecular Identification of Lactic Acid Bacteria Using 16s rRNA Genes from Fermented Teff (Eragrostis tef (Zucc.)) Dough. Int. J. Food Sci. 2018, 2018, 8510620. [Google Scholar] [CrossRef] [Green Version]
- Grimoud, J.; Durand, H.; Courtin, C.; Monsan, P.; Ouarne, F.; Theodorou, V.; Roques, C. In vitro screening of probiotic lactic acid bacteria and prebiotic glucooligosaccharides to select effective synbiotics. Anaerobe 2010, 16, 493–500. [Google Scholar] [CrossRef] [Green Version]
- Zielinska, K.; Fabiszewska, A.; Stefanska, I. Different aspects of Lactobacillus inoculants on the improvement of quality and safety of alfalfa silage. Chil. J. Agric. Res. 2015, 75, 298–306. [Google Scholar] [CrossRef] [Green Version]
- Guimaraes, A.; Venancio, A.; Abrunhosa, L. Antifungal effect of organic acids from lactic acid bacteria on Penicillium nordicum. Food Addit. Contam. Part A 2018, 35, 1803–1818. [Google Scholar] [CrossRef] [Green Version]
- Valan Arasu, M.; Jung, M.W.; Kim, D.H.; Park, H.S.; Ilavenil, S.; Al-Dhabi, N.A.; Choon Choi, K. Identification and phylogenetic characterization of novel Lactobacillus plantarum species and their metabolite profiles in grass silage. Ann. Microbiol. 2014, 65, 15–25. [Google Scholar] [CrossRef]
- Liu, C.; Zhang, Z.Y.; Dong, K.; Yuan, J.P.; Guo, X.K. Antibiotic resistance of probiotic strains of lactic acid bacteria isolated from marketed foods and drugs. Biomed. Environ. Sci. 2009, 22, 401–412. [Google Scholar] [CrossRef]
- Shazali, N.; Foo, H.L.; Loh, T.C.; Choe, D.W.; Abdul Rahim, R. Prevalence of antibiotic resistance in lactic acid bacteria isolated from the faeces of broiler chicken in Malaysia. Gut Pathog 2014, 6, 1. [Google Scholar] [CrossRef] [Green Version]
- Ji, K.; Jang, N.Y.; Kim, Y.T. Isolation of Lactic Acid Bacteria Showing Antioxidative and Probiotic Activities from Kimchi and Infant Feces. J. Microbiol. Biotechnol. 2015, 25, 1568–1577. [Google Scholar] [CrossRef] [PubMed]
- Tokatli, M.; Gulgor, G.; Bagder Elmaci, S.; Arslankoz Isleyen, N.; Ozcelik, F. In Vitro Properties of Potential Probiotic Indigenous Lactic Acid Bacteria Originating from Traditional Pickles. Biomed. Res. Int. 2015, 2015, 315819. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, B.; Wang, Y.; Tan, Z.; Li, Z.; Jiao, Z.; Huang, Q. Screening of Probiotic Activities of Lactobacilli Strains Isolated from Traditional Tibetan Qula, A Raw Yak Milk Cheese. Asian-Australas J. Anim. Sci. 2016, 29, 1490–1499. [Google Scholar] [CrossRef] [Green Version]
- Mathipa, M.G.; Thantsha, M.S. Cocktails of probiotics pre-adapted to multiple stress factors are more robust under simulated gastrointestinal conditions than their parental counterparts and exhibit enhanced antagonistic capabilities against Escherichia coli and Staphylococcus aureus. Gut Pathog 2015, 7, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, Z.; Li, J.; Chen, L.; Yuan, X.; Shao, T. Comparison of nitrogen transformation dynamics in non-irradiated and irradiated alfalfa and red clover during ensiling. Asian-Australas J. Anim. Sci. 2019, 32, 1521–1527. [Google Scholar] [CrossRef] [PubMed]
- Junges, D.; Morais, G.; Spoto, M.H.F.; Santos, P.S.; Adesogan, A.T.; Nussio, L.G.; Daniel, J.L.P. Short communication: Influence of various proteolytic sources during fermentation of reconstituted corn grain silages. J. Dairy Sci. 2017, 100, 9048–9051. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lim, Y.H.; Foo, H.L.; Loh, T.C.; Mohamad, R.; Abdullah, N. Comparative studies of versatile extracellular proteolytic activities of lactic acid bacteria and their potential for extracellular amino acid productions as feed supplements. J. Anim. Sci. Biotechnol. 2019, 10, 15. [Google Scholar] [CrossRef] [Green Version]
- Nascimento Agarussi, M.C.; Gomes Pereira, O.; Paula, R.A.; Silva, V.P.D.; Santos Roseira, J.P.; Fonseca, E.S.F. Novel lactic acid bacteria strains as inoculants on alfalfa silage fermentation. Sci. Rep. 2019, 9, 8007. [Google Scholar] [CrossRef] [Green Version]
- Ni, K.; Wang, Y.; Li, D.; Cai, Y.; Pang, H. Characterization, identification and application of lactic Acid bacteria isolated from forage paddy rice silage. PLoS ONE 2015, 10, e0121967. [Google Scholar] [CrossRef] [Green Version]
- Agarussi, M.C.N.; Pereira, O.G.; da Silva, V.P.; Leandro, E.S.; Ribeiro, K.G.; Santos, S.A. Fermentative profile and lactic acid bacterial dynamics in non-wilted and wilted alfalfa silage in tropical conditions. Mol. Biol. Rep. 2019, 46, 451–460. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Li, J.; Zhao, J.; Wu, J.; Shao, T. Enhancement of lignocellulosic degradation in high-moisture alfalfa via anaerobic bioprocess of engineered Lactococcus lactis with the function of secreting cellulase. Biotechnol. Biofuels 2019, 12, 88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oliveira, A.S.; Weinberg, Z.G.; Ogunade, I.M.; Cervantes, A.A.P.; Arriola, K.G.; Jiang, Y.; Kim, D.; Li, X.; Goncalves, M.C.M.; Vyas, D.; et al. Meta-analysis of effects of inoculation with homofermentative and facultative heterofermentative lactic acid bacteria on silage fermentation, aerobic stability, and the performance of dairy cows. J. Dairy Sci. 2017, 100, 4587–4603. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silva, V.P.; Pereira, O.G.; Leandro, E.S.; Da Silva, T.C.; Ribeiro, K.G.; Mantovani, H.C.; Santos, S.A. Effects of lactic acid bacteria with bacteriocinogenic potential on the fermentation profile and chemical composition of alfalfa silage in tropical conditions. J. Dairy Sci. 2016, 99, 1895–1902. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strains | Simulated Gastric Fluid (GF) Tolerance | Simulated Intestinal Fluid (IF) Tolerance | |||||
|---|---|---|---|---|---|---|---|
| Simulated Gastric Fluid at pH3 (Log CFU/mL) | LAB Survival Rate (%) | Simulated Intestinal Fluid at pH8 (log CFU/mL) | LAB Survival Rate (%) | ||||
| 0 h | 4 h | 6 h | 12 h | 24 h | |||
| S22 | 8.52 ± 0.08 b | 8.28 ± 0.81 b | 97.18 ± 0.73 c | 7.85 ± 0.08 NS | 7.59 ± 0.01 b | 6.59 ± 0.06 a | 77.34 ± 1.24 b |
| RJ1 | 8.76 ± 0.03 a | 8.59 ± 0.76 a | 98.05 ± 1.02 a | 7.83 ± 0.03 NS | 7.74 ± 0.02 a | 6.98± 0.06 b | 79.68 ± 0.88 a |
| STD * (F17) | 8.15 ± 0.41 c | 7.65 ± 0.12 c | 93.87 ± 1.43 b | - | - | 4.77± 0.31 c | 58.47 ± 3.86 c |
| Strains | Simulated Bile Salt Tolerance | |||||
|---|---|---|---|---|---|---|
| MRS Broth (log CFU/mL) | MRS Broth + 0.3% Oxygall (log CFU/mL) | |||||
| 0 h | 12 h | 24 h | 0 h | 12 h | 24 h | |
| S22 | 1.19 ± 0.04 NS | 4.66 ± 0.02 b | 5.62 ± 0.02 c | 3.25 ± 0.04 NS | 2.28 ± 0.01 b | 2.08 ± 0.04 c |
| RJ1 | 1.38 ± 0.07 NS | 5.92 ± 0.02 a | 7.88 ± 0.03 b | 3.23 ± 0.05 NS | 2.84 ± 0.07 a | 2.57 ± 0.03 b |
| STD * (F17) | - | - | 3.36 ± 0.01 a | - | - | 3.84 ± 0.01 a |
| Groups/Treatments | pH | Lactate (%) | Acetate (%) | Butyrate (%) | Total Bacteria (×107CFU/g) |
|---|---|---|---|---|---|
| Control | 5.56 b | 0.57 b | 0.16 b | 0.03 NS | 5.08 c |
| Lactobacillus plantarum (RJ1) | 4.53 a | 2.35 a | 0.24 a | 0.01 NS | 15.30 a |
| Pediococcus pentosaceus (S22) | 4.57 a | 2.24 a | 0.11 b | 0 NS | 12.50 b |
| Groups/Treatments | pH | Lactate (%) | Acetate (%) | Butyrate (%) | Total Bacteria (×107CFU/g) |
|---|---|---|---|---|---|
| Control | 5.83 a | 0.89 b | 0.38 a | 0.12 NS | 1.43 c |
| Lactobacillus plantarum (RJ1) | 4.58 b | 2.25 a | 0.30 a | 0 NS | 9.47 b |
| Pediococcus pentosaceus (S22) | 4.64 b | 1.75 a | 0.17 b | 0 NS | 16.71 a |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kuppusamy, P.; Kim, D.; Soundharrajan, I.; Park, H.S.; Jung, J.S.; Yang, S.H.; Choi, K.C. Low-Carbohydrate Tolerant LAB Strains Identified from Rumen Fluid: Investigation of Probiotic Activity and Legume Silage Fermentation. Microorganisms 2020, 8, 1044. https://doi.org/10.3390/microorganisms8071044
Kuppusamy P, Kim D, Soundharrajan I, Park HS, Jung JS, Yang SH, Choi KC. Low-Carbohydrate Tolerant LAB Strains Identified from Rumen Fluid: Investigation of Probiotic Activity and Legume Silage Fermentation. Microorganisms. 2020; 8(7):1044. https://doi.org/10.3390/microorganisms8071044
Chicago/Turabian StyleKuppusamy, Palaniselvam, Dahye Kim, Ilavenil Soundharrajan, Hyung Soo Park, Jeong Sung Jung, Seung Hak Yang, and Ki Choon Choi. 2020. "Low-Carbohydrate Tolerant LAB Strains Identified from Rumen Fluid: Investigation of Probiotic Activity and Legume Silage Fermentation" Microorganisms 8, no. 7: 1044. https://doi.org/10.3390/microorganisms8071044
APA StyleKuppusamy, P., Kim, D., Soundharrajan, I., Park, H. S., Jung, J. S., Yang, S. H., & Choi, K. C. (2020). Low-Carbohydrate Tolerant LAB Strains Identified from Rumen Fluid: Investigation of Probiotic Activity and Legume Silage Fermentation. Microorganisms, 8(7), 1044. https://doi.org/10.3390/microorganisms8071044