An Overview on Industrial and Medical Applications of Bio-Pigments Synthesized by Marine Bacteria

 ,
,  ,
,  ,
,  ,
,  ,
,  and
and
Abstract
:1. Introduction
1.1. Microbial Pigments
1.2. Bacterial Pigments as Natural Colorants
1.3. Marine Ecosystem as a Source of Pigment Producing Bacterial Species
2. Marine Bacterial Species as Sources of Bio-Pigments
3. Biosynthesis of Bacterial Pigments
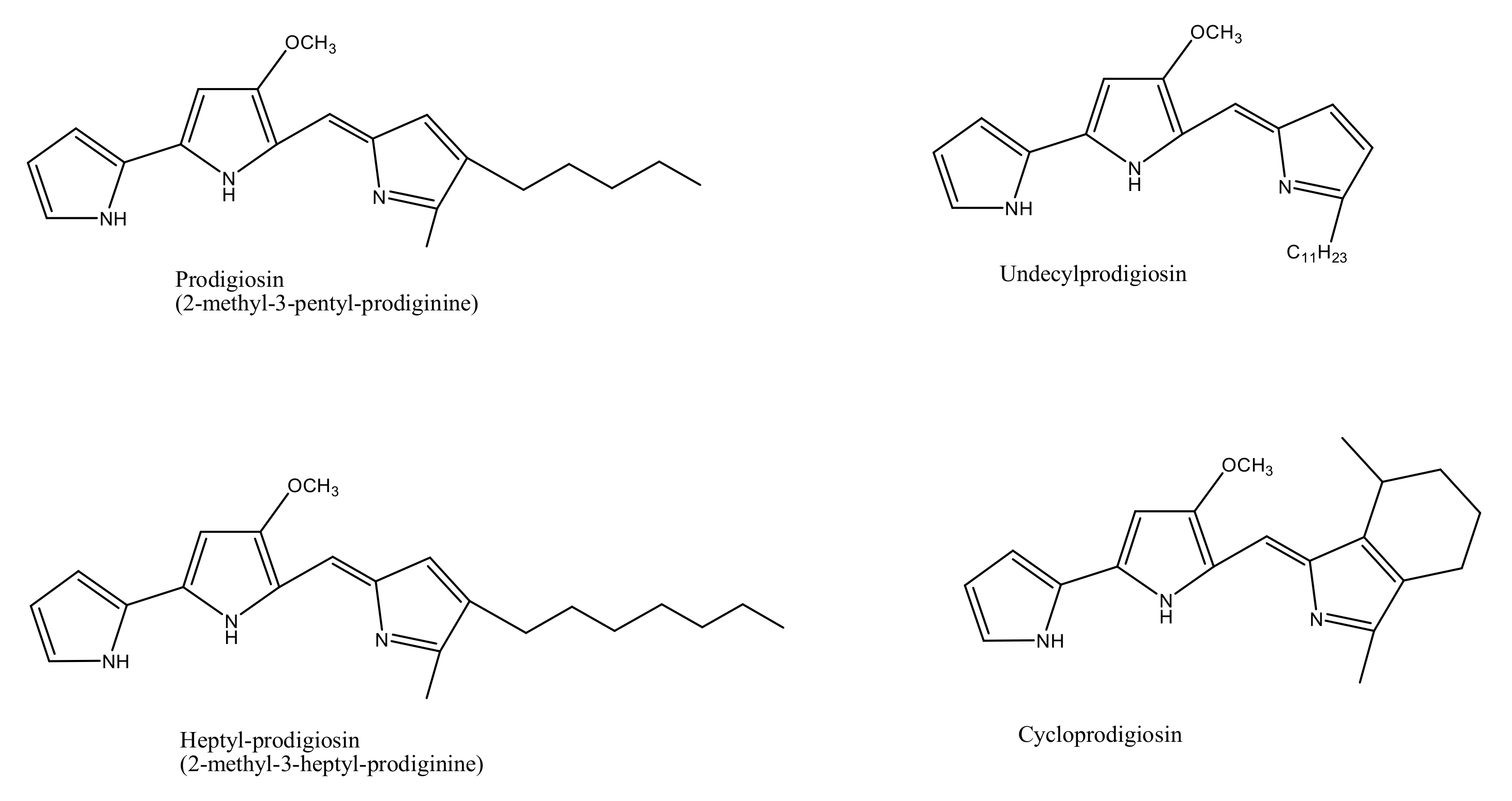
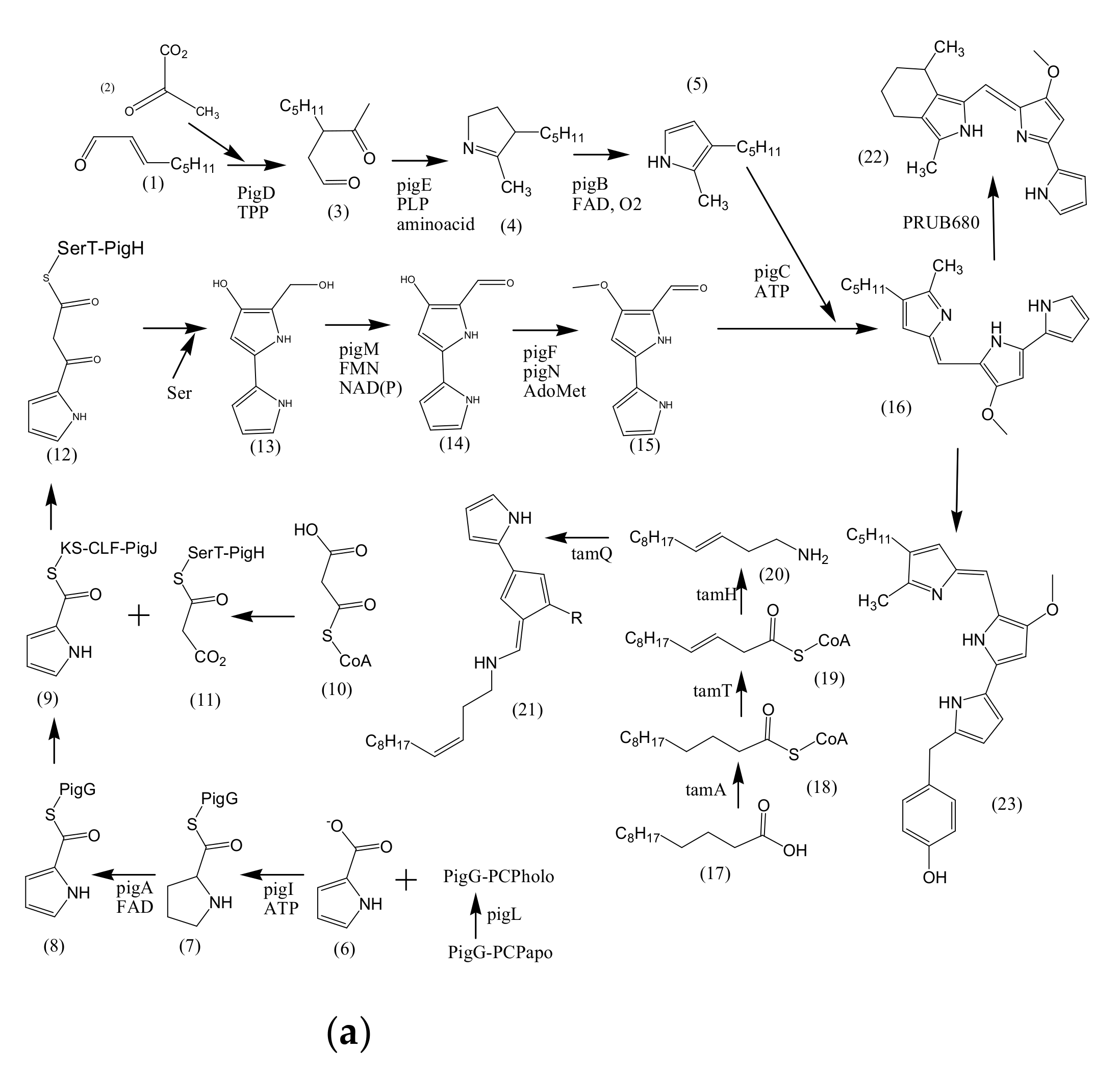
3.1. Biosynthesis of Prodiginine Analogs
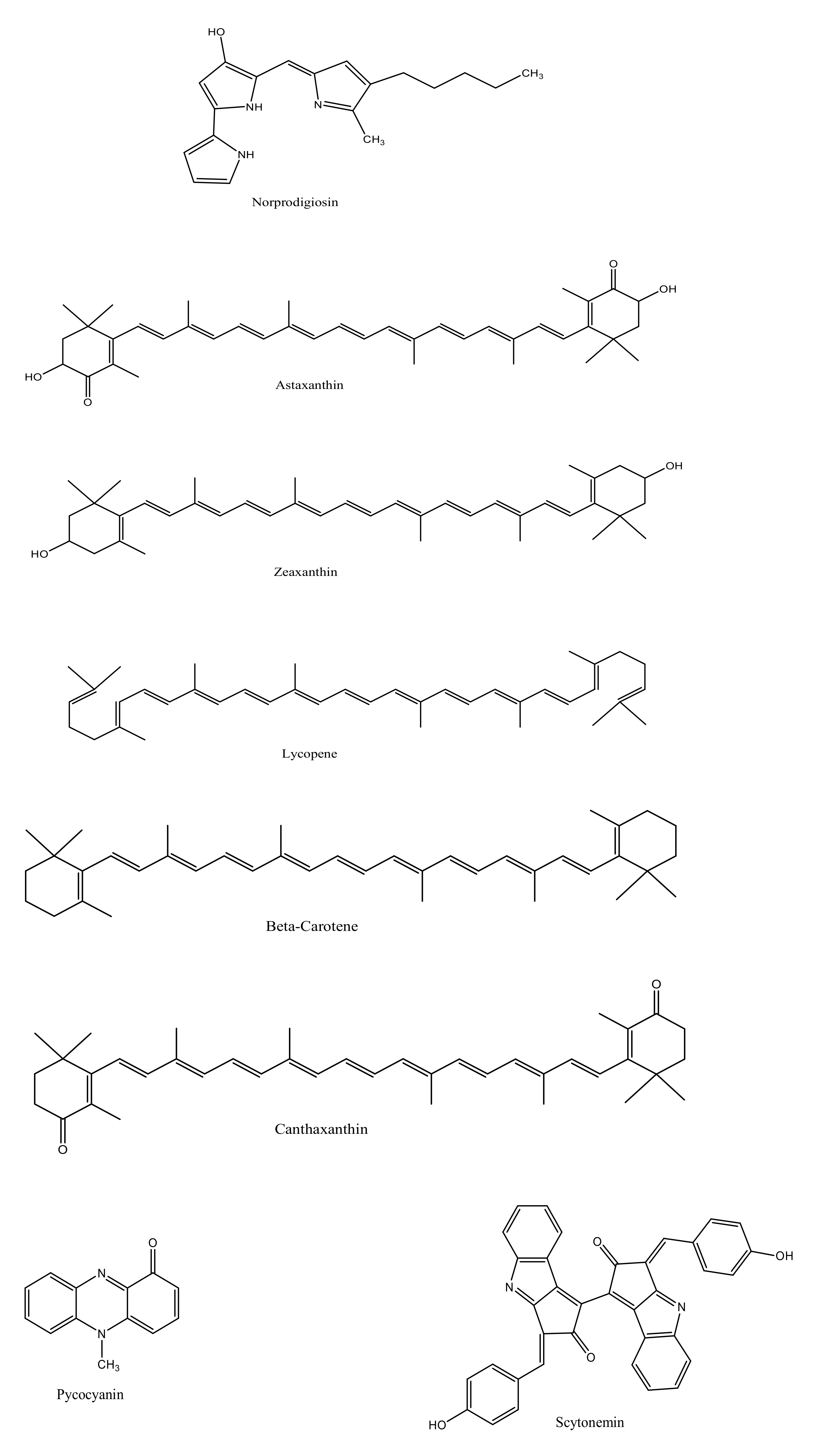
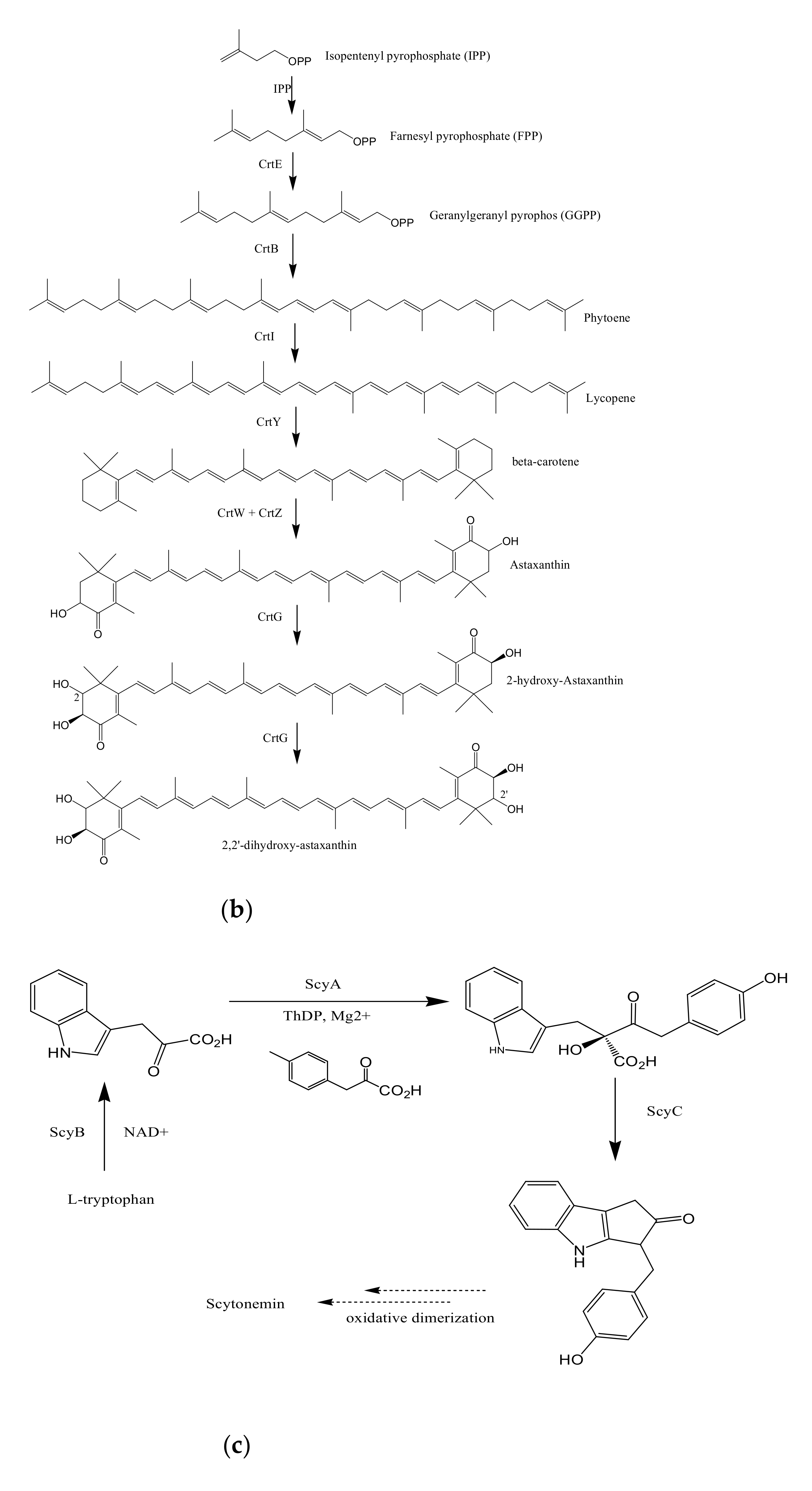
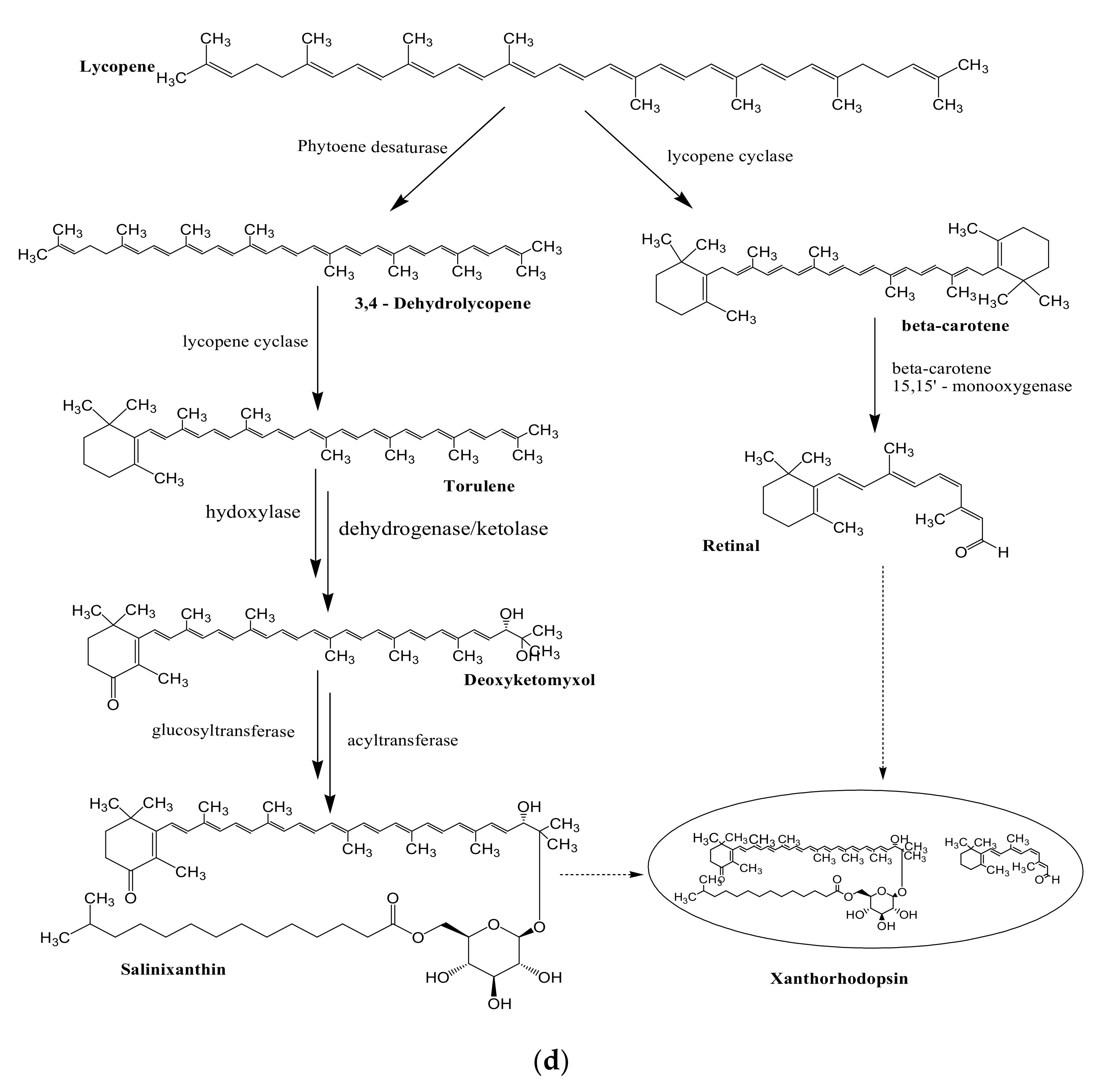
3.2. Biosynthesis of Carotenoids
3.3. Biosynthesis of Scytonemin
3.4. Biosynthesis of Salinixanthin and Retinal
4. Industrial and Therapeutic Applications
4.1. Therapeutic Applications
4.1.1. Antibacterial Activity
4.1.2. Antifungal Activity
4.1.3. Anticancer Activity
4.1.4. Antioxidant Activity
4.1.5. Antiviral Activity
4.2. Industrial Applications
4.2.1. Bio-Pigments as Food Colorants
4.2.2. Bio-Pigments as Dyeing Agents
4.2.3. Use in Cosmetics
4.2.4. Antifouling Agent
4.2.5. Photosensitizers
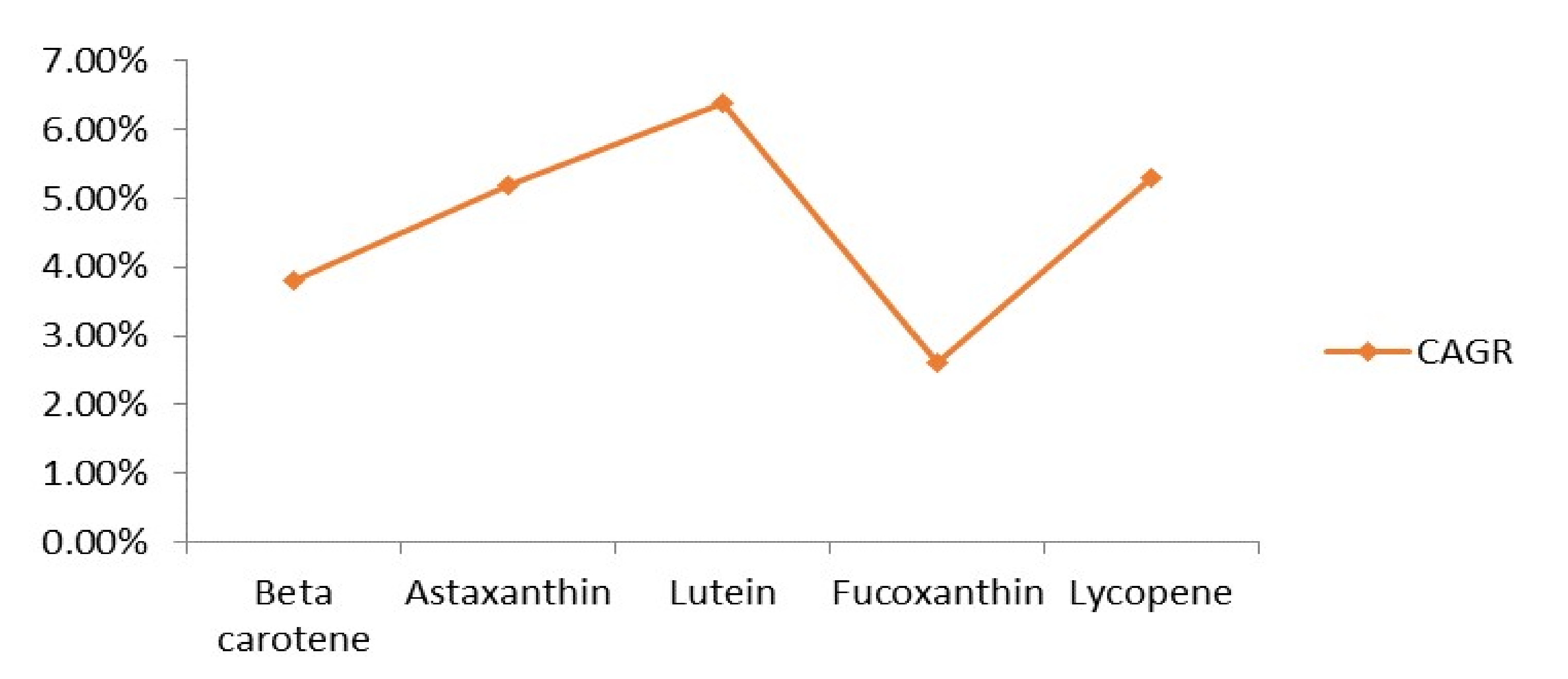
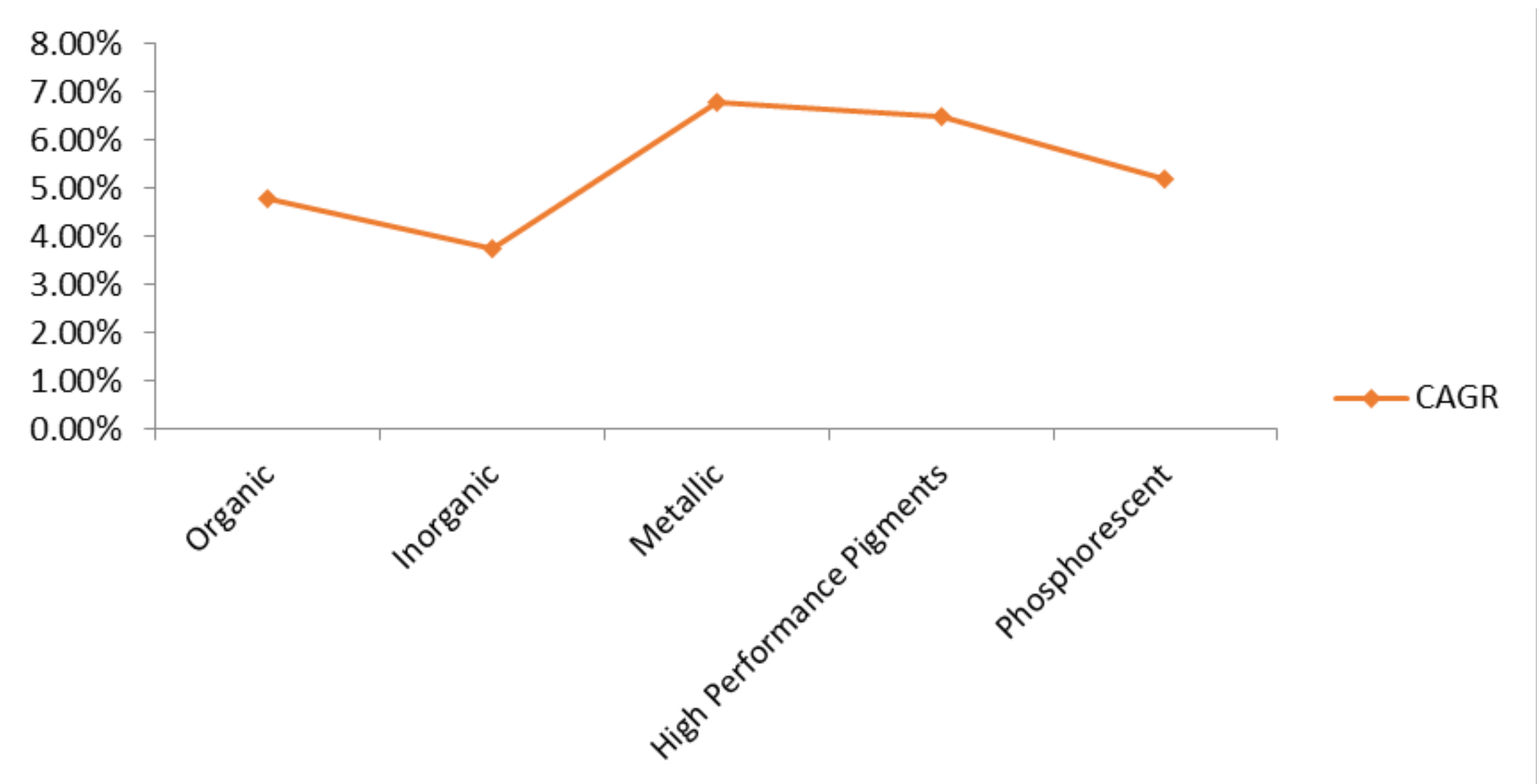
5. Industrial Importance and Global Market Trends of Pigmented Compounds
6. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Ramesh, C.; Vinithkumar, N.V.; Kirubagaran, R. Marine Pigmented Bacteria: A Prospective Source of Antibacterial Compounds. J. Nat. Sci. Biol. Med. 2019, 10, 104–113. [Google Scholar] [CrossRef]
- Venil, C.K.; Zakaria, Z.A.; Ahmad, W.A. Bacterial Pigments and Their Applications. Process Biochem. 2013, 48, 1065–1079. [Google Scholar] [CrossRef]
- Saviola, B. Pigments and Pathogenesis. J. Mycobact. Dis. 2014, 4, 5. [Google Scholar] [CrossRef] [Green Version]
- Shindo, K.; Misawa, N. New and Rare Carotenoids Isolated from Marine Bacteria and Their Antioxidant Activities. Mar. Drugs 2014, 12, 1690–1698. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Azman, A.-S.; Mawang, C.-I.; Abubakar, S. Bacterial Pigments: The Bioactivities and as an Alternative for Therapeutic Applications. Nat. Prod. Commun. 2018, 13, 1747–1754. [Google Scholar] [CrossRef] [Green Version]
- Dufossé, L. Pigments, Microbial. In Reference Module in Life Sciences; Elsevier: London, UK, 2016. [Google Scholar]
- Rodriguez-Amaya, D.B. Natural Food Pigments and Colorants. Curr. Opin. Food Sci. 2016, 7, 20–26. [Google Scholar] [CrossRef]
- Kumar, A.; Vishwakarma, H.S.; Singh, J.; Dwivedi, S.; Kumar, M. Microbial Pigments: Production and Their Applications in Various Industries. Int. J. Pharm. Chem. Biol. Sci. 2015, 5, 203–212. [Google Scholar]
- Numan, M.; Bashir, S.; Mumtaz, R.; Tayyab, S.; Rehman, N.; Khan, A.L.; Shinwari, Z.K.; Al‑Harrasi, A. Therapeutic Applications of Bacterial Pigments: A Review of Current Status and Future Opportunities. 3Biotech. 2018, 8, 207. [Google Scholar] [CrossRef]
- The Menace of Synthetic Non-Food Colours. Business Recorder. Available online: https://fp.brecorder.com/2008/02/20080216695580/ (accessed on 4 June 2020).
- Galasso, C.; Corinaldesi, C.; Sansone, C. Carotenoids from Marine Organisms: Biological Functions and Industrial Applications. Antioxidants 2017, 6, 96. [Google Scholar] [CrossRef] [Green Version]
- Petruk, G.; Roxo, M.; Lise, F.D.; Mensitieri, F.; Notomista, F.; Wink, M.; Izzo, V.; Monti, D.M. The Marine Gram-Negative Bacterium Novosphingobium sp. PP1Y as a Potential Source of Novel Metabolites with Antioxidant Activity. Biotechnol. Lett. 2019, 41, 273–281. [Google Scholar] [CrossRef]
- Stolz, P.; Obermayer, B. Manufacturing Microalgae for Skin Care. Cosmet. Toilet. 2015, 120, 99–106. [Google Scholar] [CrossRef] [Green Version]
- Bruckner, A.W. Life-Saving Products from Coral Reefs. Issues Sci. Technol. 2002, 18, 35. [Google Scholar]
- Soliev, A.B.; Hosokawa, K.; Enomoto, K. Bioactive Pigments from Marine Bacteria: Applications and Physiological Roles. Evid. Based Complement. Altern. Med. 2011, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Pawar, R.; Mohandass, C.; Rajasabapathy, R.; Meena, R.M. Molecular Diversity of Marine Pigmented Bacteria in the Central Arabian Sea with Special Reference to Antioxidant Properties. Cah. Biol. Mar. 2018, 59, 409–420. [Google Scholar]
- Baharum, S.N.; Beng, E.K.M.; Mokhtar, M.A.A. Marine Microorganisms: Potential Application and Challenges. J. Biol. Sci. 2010, 10, 555–564. [Google Scholar] [CrossRef] [Green Version]
- Podar, M.; Reysenbach, A.-L. New Opportunities Revealed by Biotechnological Explorations of Extremophiles. Curr. Opin. Biotechnol. 2006, 17, 250–255. [Google Scholar] [CrossRef]
- Giani, M.; Garbayo, I.; Vílchez, C.; Martínez-Espinosa, R.M. Haloarchaeal Carotenoids: Healthy Novel Compounds from Extreme Environments. Mar. Drugs 2019, 17, 524. [Google Scholar] [CrossRef] [Green Version]
- Oren, A. Halophilic Microbial Communities and Their Environments. Curr. Opin. Biotechnol. 2015, 33, 119–124. [Google Scholar] [CrossRef]
- Pierson, L.S.; Pierson, E.A. Metabolism and Function of Phenazines in Bacteria: Impact on the Behavior of Bacteria in the Environment and Biotechnological Process. Appl. Microbiol. Biotechnol. 2010, 86, 1659–1670. [Google Scholar] [CrossRef] [Green Version]
- Kim, D.; Kim, J.F.; Yim, J.H.; Kwon, S.-K.; Lee, C.H.; Lee, H.K. Red to Red—The Marine Bacterium Hahella chejuensis and Its Product Prodigiosin for Mitigation of Harmful Algal Blooms. J. Microbiol. Biotechnol. 2008, 18, 1621–1629. [Google Scholar]
- Setiyono, E.; Adhiwibawa, M.A.S.; Indrawati, R.; Prihastyanti, M.N.U.; Shioi, Y.; Brotosudarmo, T.H.P. An Indonesian Marine Bacterium, Pseudoalteromonas rubra, Produces Antimicrobial Prodiginine Pigments. ACS Omega 2020, 5, 4626–4635. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; Liu, G.; Li, J.; Huang, H.; Zhang, X.; Zhang, H.; Ju, J. Cytotoxic and Antibacterial Angucycline- and Prodigiosin-Analogues from the Deep-Sea Derived Streptomyces sp. SCSIO 11594. Mar. Drugs 2015, 13, 1304–1316. [Google Scholar] [CrossRef] [PubMed]
- Morgan, S.; Thomas, M.J.; Walstrom, K.M.; Warrick, E.C.; Gasper, B.J. Characterization of Prodiginine Compounds Produced by a Vibrio species Isolated from Salt Flat Sediment along the Florida Gulf Coast. Fine Focus 2016, 3, 33–51. [Google Scholar] [CrossRef] [Green Version]
- Ibrahim, D.; Nazari, T.F.; Kassim, J.; Lim, S.-H. Prodigiosin—An Antibacterial Red Pigment Produced by Serratia marcescens IBRL USM 84 Associated with a Marine Sponge Xestospongia testudinaria. J. Appl. Pharm. Sci. 2014, 4, 1–6. [Google Scholar] [CrossRef]
- Yi, H.; Chang, Y.-H.; Oh, H.W.; Bae, K.S.; Chun, J. Zooshikella ganghwensis gen. nov., sp. nov., Isolated from Tidal Flat Sediments. Int. J. Syst. Evol. Microbiol. 2003, 53, 1013–1018. [Google Scholar] [CrossRef]
- Abidin, Z.A.Z.; Ahmad, A.; Latip, J.; Usup, G. Marine Streptomyces sp. UKMCC_PT15 Producing Undecylprodigiosin with Algicidal Activity. J. Teknol. 2016, 78, 11-2. [Google Scholar] [CrossRef] [Green Version]
- Bramhachari, P.V.; Mutyala, S.; Bhatnagar, I.; Pallela, R. Novel Insights on the Symbiotic Interactions of Marine Sponge-Associated Microorganisms: Marine Microbial Biotechnology Perspective. In Marine Sponges: Chemicobiological and Biomedical Applications, 1st ed.; Pallela, R., Ehrlich, H., Eds.; Springer: New Delhi, India, 2016; pp. 69–95. [Google Scholar] [CrossRef]
- Huang, Z.; Dong, L.; Lai, Q.; Liu, J. Spartinivicinus ruber gen. nov., sp. nov., a Novel Marine Gamma proteobacterium Producing Heptylprodigiosin and Cycloheptylprodigiosin as Major Red Pigments. Front. Microbiol. 2020, 11, 11. [Google Scholar] [CrossRef]
- Xie, B.-B.; Shu, Y.-L.; Qin, Q.-L.; Rong, J.-C.; Zhang, X.-Y.; Chen, X.-L.; Zhou, B.-C.; Zhang, Y.-Z. Genome Sequence of the Cycloprodigiosin-Producing Bacterial Strain Pseudoalteromonas rubra ATCC 29570 T. J. Bacteriol. 2012, 194, 1637–1638. [Google Scholar] [CrossRef] [Green Version]
- Jafarzade, M.; Yahya, N.A.; Shayesteh, F.; Usup, G.; Ahmad, A. Influence of Culture Conditions and Medium Composition on the Production of Antibacterial Compounds by Marine Serratia sp. WPRA3. J. Microbiol. 2013, 51, 373–379. [Google Scholar] [CrossRef]
- Liu, H.; Zhang, C.; Zhang, X.; Tana, K.; Zhang, H.; Cheng, D.; Ye, T.; Li, S.; Ma, H.; Zheng, H. A Novel Carotenoids-Producing Marine Bacterium from Noble Scallop Chlamys nobilis and Antioxidant Activities of Its Carotenoid Compositions. Food Chem. 2020, 320, 126629. [Google Scholar] [CrossRef]
- Henke, N.A.; Heider, S.A.E.; Peters-Wendisch, P.; Wendisch, V.F. Production of the Marine Carotenoid Astaxanthin by Metabolically Engineered Corynebacterium glutamicum. Mar. Drugs 2016, 14, 124. [Google Scholar] [CrossRef] [PubMed]
- Asker, D. Isolation and Characterization of a Novel, Highly Selective Astaxanthin-Producing Marine Bacterium. J. Agric. Food Chem. 2017, 65, 9101–9109. [Google Scholar] [CrossRef] [PubMed]
- Shahina, M.; Hameed, A.; Lin, S.-Y.; Hsu, Y.-H.; Liu, Y.-C.; Cheng, I.-C.; Lee, M.-R.; Lai, W.-A.; Lee, R.-J.; Young, C.-C. Sphingomicrobium astaxanthinifaciens sp. nov., an Astaxanthin-Producing Glycolipid-Rich Bacterium Isolated from Surface Seawater and Emended Description of the Genus Sphingomicrobium. Int. J. Syst. Evol. Microbiol. 2013, 63, 3415–3422. [Google Scholar] [CrossRef] [PubMed]
- Mukoyama, D.; Takeyama, H.; Kondo, Y.; Matsunaga, T. Astaxanthin Formation in the Marine Photosynthetic Bacterium Rhodovulum sulfidophilum Expressing crtI, crtY, crtW and crtZ. FEMS Microbiol. Lett. 2006, 265, 69–75. [Google Scholar] [CrossRef]
- Pachaiyappan, A.; Sadhasivam, G.; Kumar, M.; Muthuvel, A. Biomedical Potential of Astaxanthin from Novel Endophytic Pigment Producing Bacteria Pontibacter korlensis AG6. Waste Biomass Valoriz. 2020, 1–11. [Google Scholar] [CrossRef]
- Balraj, J.; Pannerselvam, K.; Jayaraman, A. Isolation of Pigmented Marine Bacteria Exiguobacterium sp. from Peninsular Region of India and a Study on Biological Activity of Purified Pigment. Int. J. Sci. Techol. Res. 2014, 3, 375–384. [Google Scholar]
- Shi, X.-L.; Wu, Y.-H.; Cheng, H.; Zhang, X.-Q.; Wang, C.-S.; Xu, X.-W. Complete Genome Sequence of Astaxanthin-Producing Bacterium Altererythrobacter ishigakiensis. Mar. Genom. 2016, 30, 77–79. [Google Scholar] [CrossRef]
- Zhao, Y.; Guo, L.; Xia, Y.; Zhuang, X.; Chu, W. Isolation, Identification of Carotenoid-Producing Rhodotorula sp. from Marine Environment and Optimization for Carotenoid Production. Mar. Drugs 2019, 17, 161. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.H.; Kim, Y.T. Cloning and Characterization of the Astaxanthin Biosynthesis gene Cluster from the Marine Bacterium Paracoccus haeundaensis. Gene 2006, 370, 86–95. [Google Scholar] [CrossRef]
- Lee, J.H.; Hwang, Y.M.; Baik, K.S.; Choi, K.S.; Ka, J.-O.; Seong, C.N. Mesoflavibacter aestuarii sp. nov., a Zeaxanthin Producing Marine Bacterium Isolated from Seawater. Int. J. Syst. Evol. Microbiol. 2014, 64, 1932–1937. [Google Scholar] [CrossRef]
- Hameed, A.; Shahina, M.; Lin, S.-Y.; Lai, W.-A.; Hsu, Y.-H.; Liu, Y.-C.; Young, C.-C. Aquibacter zeaxanthinifaciens gen. nov., sp. nov., a Zeaxanthin-Producing Bacterium of the Family Flavobacteriaceae Isolated from Surface Seawater, and Emended Descriptions of the Genera Aestuariibaculum and Gaetbulibacter. Int. J. Syst. Evol. Microbiol. 2014, 64, 138–145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Asker, D.; Beppu, T.; Ueda, K. Zeaxanthinibacter enoshimensis gen. nov., sp. nov., a Novel Zeaxanthin-Producing Marine Bacterium of the Family Flavobacteriaceae, Isolated from Seawater Off Enoshima Island, Japan. Int. J. Syst. Evol. Microbiol. 2007, 57, 837–843. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shahina, M.; Hameed, A.; Lin, S.-Y.; Lee, R.-J.; Lee, M.-R.; Young, C.-C. Gramella planctonica sp. nov., a Zeaxanthin-Producing Bacterium Isolated from Surface Seawater, and Emended Descriptions of Gramella aestuarii and Gramella echinicola. Antonie van Leeuwenhoek 2014, 105, 771–779. [Google Scholar] [CrossRef] [PubMed]
- Asker, D.; Beppu, T.; Ueda, K. Mesoflavibacter zeaxanthinifaciens gen. nov., sp. nov., a Novel Zeaxanthin Producing Marine Bacterium of the Family Flavobacteriaceae. Syst. Appl. Microbiol. 2007, 30, 291–296. [Google Scholar] [CrossRef]
- Sowmya, R.; Sachindra, N.M. Carotenoid Production by Formosa sp. KMW, Marine Bacteria of Flavobacteriaceae Family: Influence of Culture Conditions and Nutrient Composition. Biocatal. Agric. Biotechnol. 2015, 4, 559–567. [Google Scholar] [CrossRef]
- Thawornwiriyanun, P.; Tanasupawat, S.; Dechsakulwatana, C.; Techkarnjanaruk, S.; Suntornsuk, W. Identification of Newly Zeaxanthin-Producing Bacteria Isolated from Sponges in the Gulf of Thailand and Their Zeaxanthin Production. Appl. Biochem. Biotechnol. 2012, 167, 2357–2368. [Google Scholar] [CrossRef]
- Hameed, A.; Arun, A.B.; Ho, H.-P.; Chang, C.-M.J.; Rekha, P.D.; Lee, M.-R.; Young, C.-C. Supercritical Carbon Dioxide Micronization of Zeaxanthin from Moderately Thermophilic Bacteria Muricauda lutaonensis CC-HSB-11T. J. Agric. Food Chem. 2011, 59, 4119–4124. [Google Scholar] [CrossRef]
- Seto, R.; Takaichi, S.; Kurihara, T.; Kishi, R.; Honda, M.; Takenaka, S.; Tsukatani, Y.; Madigan, M.T.; Wang-Otomo, Z.Y.; Kimura, Y. Lycopene-Family Carotenoids Confer Thermostability on Photocomplexes from a New Thermophilic Purple Bacterium. Biochemistry 2020, 59, 2351–2358. [Google Scholar] [CrossRef]
- Ramanathan, G.; Ramalakshmi, P. Studies on Efficacy of Marine Bacterium Salinicoccus roseus Pigment for Their Bioactive Potential. Eur. J. Biomed. Pharm. Sci. 2017, 4, 330–334. [Google Scholar]
- Montero, O.; Macías-Sánchez, M.D.; Lama, C.M.; Lubián, L.M.; Mantell, C.; Rodríguez, M.; De la Ossa, E.M. Supercritical CO2 Extraction of â-Carotene from a Marine Strain of the Cyanobacterium Synechococcus Species. J. Agric. Food Chem. 2005, 53, 9701–9707. [Google Scholar] [CrossRef]
- Hamidi, M.; Kozani, P.S.; Kozani, P.S.; Pierre, G.; Michaud, P.; Delattre, C. Marine Bacteria Versus Microalgae: Who Is the Best for Biotechnological Production of Bioactive Compounds with Antioxidant Properties and Other Biological Applications? Mar. Drugs. 2020, 18, 28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sibero, M.T.; Bachtiarini, T.U.; Trianto, A.; Lupita, A.H.; Sari, D.P.; Igarashi, Y.; Harunari, E.; Sharma, A.R.; Radjasa, O.K.; Sabdono, A. Characterization of a Yellow Pigmented Coral-Associated Bacterium Exhibiting Anti-Bacterial Activity against Multidrug Resistant (MDR) Organism. Egypt. J. Aquat. Res. 2018, 45, 81–87. [Google Scholar] [CrossRef]
- Teramoto, M.; Nishijima, M. Flavicella marina gen. nov., sp. nov., a Carotenoid-Producing Bacterium from Surface Seawater. Int. J. Syst. Evol. Microbiol. 2015, 65, 799–804. [Google Scholar] [CrossRef] [PubMed]
- Loh, W.L.C.; Huang, K.-C.; Ng, H.S.; Lan, J.C.-W. Exploring the Fermentation Characteristics of a Newly Isolated Marine Bacteria Strain, Gordonia terrae TWRH01 for Carotenoids Production. J. Biosci. Bioeng. 2020, 130, 187–194. [Google Scholar] [CrossRef]
- Saha, S.; Thavasi, T.R.; Jayalakshmi, S. Phenazine Pigments from Pseudomonas aeruginosa and Their Application as Antibacterial Agent and Food Colourants. Res. J. Microbiol. 2008, 3, 122–128. [Google Scholar] [CrossRef] [Green Version]
- Fulton, J.M.; Arthur, M.A.; Freeman, K.H. Subboreal Aridity and Scytonemin in the Holocene Black Sea. Org. Geochem. 2012, 49, 47–55. [Google Scholar] [CrossRef]
- Soule, T.; Palmer, K.; Gao, Q.; Potrafka, R.M.; Stout, V.; Garcia-Pichel, F. A Comparative Genomics Approach to Understanding the Biosynthesis of The sunscreen Scytonemin in Cyanobacteria. BMC Genom. 2009, 10, 336. [Google Scholar] [CrossRef] [Green Version]
- Thøgersen, M.S.; Delpin, M.W.; Melchiorsen, J.; Kilstrup, M.; Månsson, M.; Bunk, B.; Spröer, C.; Overmann, J.; Nielsen, K.F.; Gram, L. Production of the Bioactive Compounds Violacein and Indolmycin Is Conditional in a maeA Mutant of Pseudoalteromonas luteoviolacea S4054 Lacking the Malic Enzyme. Front. Microbiol. 2016, 7. [Google Scholar] [CrossRef] [Green Version]
- Aye, A.M.; Bonnin-Jusserand, M.; Brian-Jaisson, F.; Ortalo-Magne, A.; Culioli, G.; Nevry, R.K.; Rabah, N.; Blache, Y.; Molmeret, M. Modulation of Violacein Production and Phenotypes Associated with Biofilm by Exogenous Quorum Sensing N-acylhomoserine Lactones in the Marine Bacterium Pseudoalteromonas ulvae TC14. Microbiology 2015, 161, 2039–2052. [Google Scholar] [CrossRef]
- Dang, H.T.; Yotsumoto, K.; Enomoto, K. Draft Genome Sequence of Violacein-Producing Marine Bacterium Pseudoalteromonas sp. 520P1. Genome Announc. 2014, 2. [Google Scholar] [CrossRef] [Green Version]
- Ballestriero, F.; Daim, M.; Penesyan, A.; Nappi, J.; Schleheck, D.; Bazzicalupo, P.; Schiavi, E.D.; Egan, S. Antinematode Activity of Violacein and the Role of the Insulin/IGF-1 Pathway in Controlling Violacein Sensitivity in Caenorhabditis elegans. PLoS ONE 2014, 9, e109201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, Y.-H.; Cheng, H.; Xu, L.; Jin, X.-B.; Wang, C.-S.; Xu, X.-W. Physiological and Genomic Features of a Novel Violacein-Producing Bacterium Isolated from Surface Seawater. PLoS ONE 2017, 12, e0179997. [Google Scholar] [CrossRef] [PubMed]
- Yada, S.; Wang, Y.; Zou, Y.; Nagasaki, K.; Hosokawa, K.; Osaka, I.; Arakawa, R.; Enomoto, K. Isolation and Characterization of Two Groups of Novel Marine Bacteria Producing Violacein. Mar. Biotechnol. 2008, 10, 128–132. [Google Scholar] [CrossRef] [PubMed]
- Hakvåg, S.; Fjærvik, E.; Klinkenberg, G.; Borgos, S.E.F.; Josefsen, K.D.; Ellingsen, T.E.; Zotchev, S.B. Violacein-Producing Collimonas sp. from the Sea Surface Microlayer of Coastal Waters in Trøndelag, Norway. Mar. Drugs 2009, 7, 576–588. [Google Scholar] [CrossRef] [Green Version]
- Vasanthabharathi, V.; Lakshminarayanan, R.; Jayalakshmi, S. Melanin Production from Marine Streptomyces. Afr. J. Biotechnol. 2011, 10, 11224–11234. [Google Scholar] [CrossRef] [Green Version]
- Tarangini, K.; Mishra, S. Production, Characterization and Analysis of Melanin from Isolated Marine Pseudomonas sp. Using Vegetable Waste. Res. J. Eng. Sci. 2013, 2, 40–46. [Google Scholar]
- Lucas-Elío, P.; Goodwin, L.; Woyke, T.; Pitluck, S.; Nolan, M.; Kyrpides, N.C.; Detter, J.C.; Copeland, A.; Teshima, H.; Bruce, D.; et al. The Genomic Standards Consortium Complete Genome Sequence of the Melanogenic Marine Bacterium Marinomonas mediterranea Type Strain (MMB-1T). Stand. Genom. Sci. 2012, 6, 63–73. [Google Scholar] [CrossRef]
- Manirethan, V.; Raval, K.; Balakrishnan, R.M. Adsorptive Removal of Trivalent and Pentavalent Arsenic from Aqueous Solutions Using Iron and Copper Impregnated Melanin Extracted from the Marine Bacterium Pseudomonas stutzeri. Environ. Pollut. 2019, 257, 113576. [Google Scholar] [CrossRef]
- Kurian, N.K.; Nair, H.P.; Bhat, S.G. Evaluation of Anti-Inflammatory Property of Melanin from Marine Bacillus spp. BTCZ31. Asian J. Pharm. Clin. Res. 2015, 8, 251–255. [Google Scholar]
- Sivaperumal, P.; Kamala, K.; Rajaram, R.; Mishra, S.S. Melanin from Marine Streptomyces sp. (MVCS13) with Potential Effect against Ornamental Fish Pathogens of Carassius auratus. Biocatal. Agric. Biotechnol. 2014, 3, 134–141. [Google Scholar] [CrossRef]
- Kurian, N.K.; Bhat, S.G. Photoprotection and Anti-Inflammatory Properties of Non–Cytotoxic Melanin from Marine Isolate Providencia rettgeri Strain BTKKS1. Biosci. Biotechnol. Res. Asia 2017, 14, 1475–1484. [Google Scholar] [CrossRef]
- Kurian, N.K.; Nair, H.P.; Bhat, S.G. Characterization of Melanin Producing Bacteria Isolated from 96m depth Arabian Sea Sediments. Res. J. Biotechnol. 2019, 14, 64–71. [Google Scholar]
- Kamarudheen, N.; Naushad, T.; Rao, K.V.B. Biosynthesis, Characterization and Antagonistic Applications of Extracellular Melanin Pigment from Marine Nocardiopsis Sps. Indian J. Pharm. Educ. Res. 2019, 53, 112–120. [Google Scholar] [CrossRef] [Green Version]
- Kurian, N.K.; Bhat, S.G. Food, Cosmetic and Biological Applications of Characterized DOPA-Melanin from Vibrio alginolyticus strain BTKKS3. Appl. Biol. Chem. 2018, 61, 163–171. [Google Scholar] [CrossRef]
- Sakai-Kawada, F.E.; Ip, C.G.; Hagiwara, K.A.; Awaya, J.D. Biosynthesis and Bioactivity of Prodiginine Analogs in Marine Bacteria, Pseudoalteromonas: A Mini Review. Front. Microbiol. 2019, 10, 1715. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Picott, K.J.; Deichert, J.A.; De Kemp, E.M.; Schatte, G.; Sauriol, F.; Ross, A.C. Isolation and Characterization of Tambjamine MYP1, a Macrocyclic Tambjamine Analogue from Marine Bacterium Pseudoalteromonas citrea. MedChemComm 2019, 10, 478–483. [Google Scholar] [CrossRef]
- Harris, A.K.P.; Williamson, N.R.; Slater, H.; Cox, A.; Abbasi, S.; Foulds, I.; Simonsen, H.T.; Leeper, F.J.; Salmond, G.P.C. The Serratia Gene Cluster Encoding Biosynthesis of the Red Antibiotic, Prodigiosin, Shows Species- and Strain-Dependent Genome Context Variation. Microbiology 2004, 150, 3547–3560. [Google Scholar] [CrossRef]
- Williamson, N.R.; Simonsen, H.T.; Ahmed, R.A.A.; Goldet, G.; Slater, H.; Woodley, L.; Leeper, F.J.; Salmond, G.P.C. Biosynthesis of the Red Antibiotic, Prodigiosin, in Serratia: Identification of a Novel 2-methyl-3-n-amyl-Pyrroie (MAP) Assembly Pathway, Definition of the Terminal Condensing Enzyme, and Implications for Undecylprodigiosin Biosynthesis in Streptomyces. Mol. Microbiol. 2005, 56, 971–989. [Google Scholar] [CrossRef]
- Garneau-Tsodikova, S.; Dorrestein, P.C.; Kelleher, N.L.; Walsh, T.C. Protein Assembly Line Components in Prodigiosin Biosynthesis: Characterization of PigA, G, H, I, J. J. Am. Chem. Soc. 2006, 128, 12600–12601. [Google Scholar] [CrossRef]
- Kimata, S.; Izawa, M.; Kawasaki, T.; Hayakawa, Y. Identification of a Prodigiosin Cyclization Gene in the Roseophilin Producer and Production of a New Cyclized Prodigiosin in a Heterologous Host. J. Antibiot. 2017, 70, 196–199. [Google Scholar] [CrossRef]
- De Rond, T.; Stow, P.; Eigl, I.; Johnson, R.E.; Chan, L.J.G.; Goyal, G.; Baidoo, E.E.K.; Hillson, N.J.; Petzold, C.J.; Sarpong, R.; et al. Oxidative Cyclization of Prodigiosin by an Alkylglycerol Monooxygenase-Like Enzyme. Nat. Chem. Biol. 2017, 13, 1155–1159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burke, C.; Thomas, T.; Egan, S.; Kjelleberg, S. The Use of Functional Genomics for the Identification of a Gene Cluster Encoding for the Biosynthesis of an Antifungal Tambjamine in the Marine Bacterium Pseudoalteromonas tunicata: Brief Report. Environ. Microbiol. 2007, 9, 814–818. [Google Scholar] [CrossRef] [PubMed]
- Marchetti, P.M.; Kelly, V.; Simpson, J.P.; Ward, M.; Campopiano, D.J. The Carbon chain-Selective Adenylation Enzyme TamA: The Missing Link between Fatty Acid and Pyrrole Natural Product Biosynthesis. Org. Biomol. Chem. 2018, 16, 2735–2740. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Asker, D.; Awad, T.S.; Beppu, T.; Ueda, K. Rapid and Selective Screening Method for Isolation and Identification of Carotenoid-Producing Bacteria. Methods Mol. Biol. 2018, 1852, 143–170. [Google Scholar] [CrossRef]
- Yabuzaki, J. Carotenoids Database: Structures, Chemical Fingerprints and Distribution among Organisms. Database 2017, 2017. [Google Scholar] [CrossRef] [Green Version]
- Scaife, M.A.; Burja, A.M.; Wright, P.C. Characterization of Cyanobacterial β-Carotene Ketolase and Hydroxylase Genes in Escherichia coli, and Their Application for Astaxanthin Biosynthesis. Biotechnol. Bioeng. 2009, 103, 944–955. [Google Scholar] [CrossRef]
- Soule, T.; Stout, V.; Swingley, W.D.; Meeks, J.C.; Garcia-Pichel, F. Molecular Genetics and Genomic Analysis of Scytonemin Biosynthesis in Nostoc punctiforme ATCC 29133. J. Bacteriol. 2007, 189, 4465–4472. [Google Scholar] [CrossRef] [Green Version]
- Balskus, E.P.; Case, R.J.; Walsh, C.T. The Biosynthesis of Cyanobacterial Sunscreen Scytonemin in Intertidal Microbial Mat Communities. FEMS Microbiol. Ecol. 2011, 77, 322–332. [Google Scholar] [CrossRef]
- Estrada, A.F.; Maier, D.; Scherzinger, D.; Avalos, J.; Al-Babili, S. Novel Apo Carotenoid Intermediates in Neurospora crassa Mutants Imply a New Biosynthetic Reaction Sequence Leading to Neurosporaxanthin Formation. Fungal Genet. Biol. 2008, 45, 1497–1505. [Google Scholar] [CrossRef]
- Liao, L.; Su, S.; Zhao, B.; Fan, C.; Zhang, J.; Li, H.; Chen, B. Biosynthetic Potential of a Novel Antarctic Actinobacterium Marisediminicola antarctica ZS314T Revealed by Genomic Data Mining and Pigment Characterization. Mar. Drugs. 2019, 17, 388. [Google Scholar] [CrossRef]
- Balashov, S.P.; Imasheva, E.S.; Boichenko, V.A.; Antón, J.; Wang, J.M.; Lanyi, J.K. Xanthorhodopsin: A Proton Pump with a Light-Harvesting Carotenoid Antenna. Science 2005, 309, 2061–2064. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lanyi, J.K.; Balashov, S.P. Xanthorhodopsin: A Bacteriorhodopsin-Like Proton Pump with a Carotenoid Antenna. Biochim. Biophys. Acta 2008, 1777, 684–688. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Graham, J.E.; Bryant, D.A. The Biosynthetic Pathway for Myxol-2’fucoside (Myxoxanthophyll) in the Cyanobacterium Synechococcus sp. Strain PCC 7002. J. Bacteriol. 2009, 191, 3292–3300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hannibal, L.; Lorquin, J.; Dortoli, N.A.; Garcia, N.; Chaintreuil, C.; Massonboivin, C.; Dreyfus, B.; Giraud, E. Isolation and Characterization of Canthaxanthin Biosynthesis Genes from the Photosynthetic Bacterium Bradyrhizobium sp. Strain ORS278. J. Bacteriol. 2000, 182, 3850–3853. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.C.; Kim, Y.-S.; Park, S.; Kim, J.; Kang, S.-J.; Lee, M.-H.; Ryu, S.; Choi, J.M.; Oh, T.-K.; Yoon, J.-H. Exceptional Production of Both Prodigiosin and Cycloprodigiosin as Major Metabolic Constituents by a Novel Marine Bacterium, Zooshikella rubidus S1-1. Appl. Environ. Microbiol. 2011, 77, 4967–4973. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karbalaei-Heidari, H.R.; Partovifar, M.; Memarpoor-Yazdi, M. Evaluation of the Bioactive Potential of Secondary Metabolites Produced by a New Marine Micrococcus Species Isolated from the Persian Gulf. Avicenna J. Med. Biotechnol. 2020, 12, 61–65. [Google Scholar] [PubMed]
- Atalah, J.; Blamey, L.; Muñoz‑Ibacache, S.; Gutierrez, F.; Urzua, M.; Encinas, M.V.; Páez, M.; Sun, J.; Blamey, J.M. Isolation and Characterization of Violacein from an Antarctic Iodobacter: A Non‑Pathogenic Psychrotolerant Microorganism. Extremophiles 2019, 24, 43–52. [Google Scholar] [CrossRef]
- Srilekha, V.; Krishna, G.; Srinivas, V.S.; Charya, M.A.S. Antimicrobial evaluation of bioactive pigment from Salinicoccus sp. isolated from Nellore sea coast. Int. J. Biotechnol. Biochem. 2017, 13, 211–217. [Google Scholar]
- Asencio, G.; Lavin, P.; Alegría, K.; Domínguez, M.; Bello, H.; González-Rocha, G.; González-Aravena, M. Antibacterial Activity of the Antarctic Bacterium Janthinobacterium sp. SMN 33.6 against Multi-Resistant Gram-Negative Bacteria. Electron. J. Biotechnol. 2014, 17, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Sasidharan, A.; Sasidharan, N.K.; Amma, D.B.N.S.; Vasu, R.K.; Nataraja, A.V.; Bhaskaran, K. Antifungal Activity of Violacein Purified from a Novel Strain of Chromobacterium sp. NIIST (MTCC 5522). J. Microbiol. 2015, 53, 694–701. [Google Scholar] [CrossRef]
- Fehér, D.; Barlow, R.S.; Lorenzo, P.S.; Hemscheidt, T.K. A 2-Substituted Prodiginine, 2-(p-Hydroxybenzyl) Prodigiosin, from Pseudoalteromonas rubra. J. Nat. Prod. 2008, 71, 1970–1972. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patil, S.; Paradeshi, J.; Chaudhari, B. Anti-Melanoma and UV-B Protective Effect of Microbial Pigment Produced by Marine Pseudomonas aeruginosa GS-33. Nat. Prod. Res. 2016, 30, 2835–2839. [Google Scholar] [CrossRef] [PubMed]
- Alem, D.; Marizcurrena, J.J.; Saravia, V.; Davyt, D.; Martinez‑Lopez, W.; Castro‑Sowinski, S. Production and Antiproliferative Effect of Violacein, a Purple Pigment Produced by an Antarctic Bacterial Isolate. World J. Microbiol. Biotechnol. 2020, 36, 120. [Google Scholar] [CrossRef] [PubMed]
- Pinkerton, D.M.; Banwell, M.G.; Garson, M.J.; Kumar, N.; De Moraes, M.O.; Cavalcanti, B.C.; Pessoa, C. Antimicrobial and Cytotoxic Activities of Synthetically Derived Tambjamines C and E - J, BE-18591, and a Related Alkaloid from the Marine Bacterium Pseudoalteromonas tunicata. Chem. Biodivers. 2010, 7, 1311–1324. [Google Scholar] [CrossRef]
- Afra, S.; Makhdoumi, A.; Matin, M.M.; Feizy, J. A Novel Red Pigment from Marine Arthrobacter sp. G20 with Specific Anticancer Activity. J. Appl. Microbiol. 2017, 123, 1228–1236. [Google Scholar] [CrossRef]
- Miao, L.; Wang, X.; Jiang, W.; Yang, S.; Zhou, H.; Zhai, Y.; Zhou, X.; Dong, K. Optimization of the Culture Condition for an Antitumor Bacterium Serratia proteamacula 657 and Identification of the Active Compounds. World J. Microbiol. Biotechnol. 2012, 29, 855–863. [Google Scholar] [CrossRef]
- Shindo, K.; Kikuta, K.; Suzuki, A.; Katsuta, A.; Kasai, H.; Yasumoto-Hirose, M.; Matsuo, Y.; Misawa, N.; Takaichi, S. Rare Carotenoids, (3R)-saproxanthin and (3R,2′S)-myxol, Isolated from Novel Marine Bacteria (Flavobacteriaceae) and Their Antioxidative Activities. Appl. Microbiol. Biotechnol. 2007, 74, 1350–1357. [Google Scholar] [CrossRef]
- Prabhu, S.; Rekha, P.D.; Young, C.C.; Hameed, A.; Lin, S.-Y.; Arun, A.B. Zeaxanthin Production by Novel Marine Isolates from Coastal Sand of India and Its Antioxidant Properties. Appl. Biochem. Biotechnol. 2013, 171, 817–831. [Google Scholar] [CrossRef]
- Kumar, G.; Sahu, N.; Reddy, G.N.; Prasad, R.B.N.; Nagesh, N.; Kamal, A. Production of Melanin Pigment from Pseudomonas stutzeri Isolated from Red Seaweed Hypneamusci formis. Lett. Appl. Microbiol. 2013, 57, 295–302. [Google Scholar] [CrossRef]
- Renugadevi, K.; Nachiyar, C.V.; Sowmiya, P.; Sunkar, S. Antioxidant Activity of Phycocyanin Pigment Extracted from Marine Filamentous Cyanobacteria Geitlerinema sp TRV57. Biocatal. Agric. Biotechnol. 2018, 16, 237–242. [Google Scholar] [CrossRef]
- Srinivasan, M.; Keziah, S.M.; Hemalatha, M.; Devi, C.S. Pigment from Streptomyces bellus MSA1 Isolated from Marine Sediments. IOP Conf. Ser. Mater. Sci. Eng. 2017, 263. [Google Scholar] [CrossRef]
- Silva, T.R.; Tavares, R.S.N.; Canela-Garayoa, R.; Eras, J.; Rodrigues, M.V.N.; Neri-Numa, I.A.; Pastore, G.M.; Rosa, L.H.; Schultz, J.A.A.; Debonsi, H.M.; et al. Chemical Characterization and Biotechnological Applicability of Pigments Isolated from Antarctic Bacteria. Mar. Biotechnol. 2019, 21, 416–429. [Google Scholar] [CrossRef] [PubMed]
- Metwally, R.A.; Abeer, A.; El-Sikaily, A.; El-Sersy, N.A.; Ghozlan, H.; Sabry, S. Biological Activity of Prodigiosin from Serratia rubidaea RAM_Alex. Res. J. Biotechnol. 2019, 14, 100. [Google Scholar]
- Hegazy, G.E.; Abu-Serie, M.M.; Abo-Elela, G.M.; Ghozlan, H.; Sabry, S.A.; Soliman, N.A.; Abdel-Fattah, Y.R. In Vitro Dual (Anticancer and Antiviral) Activity of the Carotenoids Produced by Haloalkaliphilic Archaeon Natrialba sp. M6. Sci. Rep. 2020, 10, 5986. [Google Scholar] [CrossRef]
- Torregrosa-Crespo, J.; Montero, Z.; Fuentes, J.L.; García-Galbis, M.R.; Garbayo, I.; Vílchez, C.; Martínez-Espinosa, R.M. Exploring the Valuable Carotenoids for the Large-Scale Production by Marine Microorganisms. Mar. Drugs 2018, 16, 203. [Google Scholar] [CrossRef] [Green Version]
- Dharmaraj, S.; Ashokkumar, B.; Dhevendaran, K. Food-Grade Pigments from Streptomyces sp. Isolated from the Marine Sponge Callyspongia diffusa. Food Res. Int. 2009, 42, 487–492. [Google Scholar] [CrossRef]
- Ramesh, C.; Vinithkumar, N.V.; Kirubagaran, R.; Venil, C.K.; Dufossé, L. Applications of Prodigiosin Extracted from Marine Red Pigmented Bacteria Zooshikella sp. and Actinomycete Streptomyces sp. Microorganisms 2020, 8, 556. [Google Scholar] [CrossRef]
- Alihosseini, F.; Ju, K.-S.; Lango, J.; Hammock, B.D.; Sun, G. Antibacterial Colorants: Characterization of Prodiginines and Their Applications on Textile Materials. Biotechnol. Prog. 2008, 24, 742–747. [Google Scholar] [CrossRef] [Green Version]
- Anti-Bacterial Fabric Holds Promise for Fighting Superbug. Science Daily. Ulsan National Institute of Science and Technology (UNIST). Available online: https://www.sciencedaily.com/releases/2016/03/160308091646.htm (accessed on 8 March 2020).
- Michael, R. New Antibiotic Dye May Help Prevent Infectious Diseases. Contagion Live. Available online: https://www.contagionlive.com/news/new-antibiotic-dye-may-help-prevent-infectious-diseases (accessed on 4 April 2020).
- Krishna, J.G.; Jacob, A.; Kurian, P.; Elyas, K.K.; Chandrasekaran, M. Marine Bacterial Prodigiosin as Dye for Rubber Latex, Polymethyl Methacrylate Sheets and Paper. Afr. J. Biotechnol. 2013, 12, 2266–2269. [Google Scholar] [CrossRef] [Green Version]
- Derikvand, P.; Llewellyn, C.A.; Purton, S. Cyanobacterial Metabolites as a Source of Sunscreens and Moisturizers: A Comparison with Current Synthetic Compounds. Eur. J. Phycol. 2017, 52, 43–56. [Google Scholar] [CrossRef]
- Consumption Value of Cosmetics and Personal Care in Europe in 2018, by Country (in Million Euros). Available online: https://www.statista.com/statistics/382100/european-cosmetics-market-volume-by-country/ (accessed on 24 June 2020).
- Poulose, N.; Sajayan, A.; Ravindran, A.; Sreechitra, T.; Vardhan, V.; Selvin, J.; Kiran, G.S. Photoprotective Effect of Nanomelanin-Seaweed Concentrate in Formulated Cosmetic Cream: With Improved Antioxidant and Wound Healing Properties. J. Photochem. Photobiol. B Biol. 2020, 205. [Google Scholar] [CrossRef]
- Wang, Z.; Tschirhart, T.; Schultzhaus, Z.; Kelly, E.E.; Chen, A.; Oh, E.; Nag, O.; Glaser, E.R.; Kim, E.; Lloyd, P.F.; et al. Characterization and Application of Melanin Produced by the Fast-Growing Marine Bacterium Vibrio natriegens Through Heterologous Biosynthesis. Appl. Environ. Microbiol. 2019, 86. [Google Scholar] [CrossRef] [PubMed]
- Priya, K.A.; Satheesh, S.; Balasubramaniem, A.K.; Varalakshmi, P.; Gopal, S.; Natesan, S. Antifouling Activity of Prodigiosin from Estuarine Isolate of Serratia marcescens CMST 07. In Microbiological Research In Agroecosystem Management, 1st ed.; Velu, R.K., Ed.; Springer: New Delhi, India, 2013; pp. 11–21. [Google Scholar] [CrossRef]
- Zeng, Z.; Guo, X.-P.; Cai, X.; Wang, P.; Li, B.; Yang, J.-L.; Wang, X. Pyomelanin from Pseudoalteromonas lipolytica Reduces Biofouling. Microbiol. Biotechnol. 2017, 10, 1718–1731. [Google Scholar] [CrossRef] [PubMed]
- Hernández-Velasco, P.; Morales-Atilano, I.; Rodríguez-Delgado, M.; Rodríguez-Delgado, J.M.; Luna-Moreno, D.; Ávalos-Alanís, F.G.; Villarreal-Chiu, J.F. Photoelectric Evaluation of Dye-Sensitized Solar Cells Based on Prodigiosin Pigment Derived from Serratia marcescens 11E. Dyes Pigment. 2020, 177, 108278. [Google Scholar] [CrossRef]
- Global Astaxanthin Market—Sources, Technologies and Applications. Available online: https://www.marketresearch.com/Industry-Experts-v3766/Global-Astaxanthin-Sources-Technologies-Applications-8827191/ (accessed on 30 March 2020).
- Astaxanthin Market Size, Share & Trends Analysis Report by Source, by Product, by Application and Segment Forecasts, 2020–2027. Available online: https://www.reportlinker.com/p05868776/Astaxanthin-Market-Size-Share-Trends-Analysis-Report-By-Source-By-Product-By-Application-And-Segment-Forecasts.html (accessed on 1 June 2020).
- Prodigiosin Serratia marcescens - CAS 82-89-3 - Calbiochem. Merck: A Leader in Life Sciences. Available online: https://www.merckmillipore.com/INTL/en/product/Prodigiosin-Serratia-marcescens-CAS-82-89-3-Calbiochem,EMD_BIO-529685?ReferrerURL=https%3A%2F%2Fwww.google.com%2F&bd=1 (accessed on 6 June 2020).
- Paracoccus Powder (Astaxanthin Powder) 2OZ. Brine Shrimp Direct. Available online: https://www.brineshrimpdirect.com/paracoccus-powder-astaxanthin-powder-2-ounce (accessed on 6 June 2020).
- Paracoccus Powder, Natural Source of Astaxanthin, 50g. NoCoast Aquatics. Available online: https://www.nocoastaquatics.com/products/paracoccus-powder-astaxanthin (accessed on 7 June 2020).
- Violacein (V9389)—Datasheet—Sigma-Aldrich. Available online: https://www.sigmaaldrich.com/content/dam/sigmaaldrich/docs/Sigma/Datasheet/2/v9389dat.pdf (accessed on 6 August 2020).
- Prodigiosin antibiotic. My Biosource. Available online: https://www.mybiosource.com/antibiotic/prodigiosin/651317 (accessed on 6 August 2020).
- Suppliers for Prodigiosin 25c. BuyersGuideChem. Available online: https://www.buyersguidechem.com/AliefAus.php?pnumm=121199138105 (accessed on 15 June 2020).
- Suppliers for Prodigiosin. BuyersGuideChem. Available online: https://www.buyersguidechem.com/AliefAus.php?pnumm=553227439869&modus=einprod (accessed on 21 June 2020).
- Lucantin® Pink (Astaxanthin). Animal Nutrition Products. BASF. Available online: https://nutrition.basf.com/global/en/animal-nutrition/products/lucantin-pink.html (accessed on 15 June 2020).
- AstaSana™ 10% FS. UL Prospector. Available online: https://www.ulprospector.com/pt/eu/Food/Detail/15324/541591/AstaSana-10-FS (accessed on 15 June 2020).
- J-Bio™ Astxanthin. GMP Global Marketing, Inc. Available online: https://www.gmpglobalmarketing.com/portfolio-items/j-bio-astaxanthin/ (accessed on 27 June 2020).
- BioAstin® Hawaiian Astaxanthin. Cyanotech Corporation. Foods of Hawaii. Available online: https://www.foodsofhawaii.com/author/cyanotech-corporation/?user=132 (accessed on 27 June 2020).
- Suppliers for Astaxanthin: BuyersGuideChem. Available online: https://www.buyersguidechem.com/AliefAus.php?pnumm=244231380730 (accessed on 1 July 2020).
- ZeaGold®, Zeaxanthin. Kalsec. Available online: https://www.kalsec.com/products/zeaxanthin/ (accessed on 1 July 2020).
- MacuShield® softgel capsule. AGP Limited. Available online: https://agp.com.pk/agp_products/macushield/ (accessed on 3 July 2020).
- OPTISHARP® (Zeaxanthin) 20% FS by DSM Nutritional Products, Inc. Prospector. Available online: https://www.ulprospector.com/en/na/Food/Detail/6295/239896/OPTISHARP-Zeaxanthin-20-FS (accessed on 1 July 2020).
- Suppliers for Zeaxanthin. Available online: https://www.buyersguidechem.com/AliefAus.php?pnumm=750686624198 (accessed on 1 July 2020).
- Redivivo® (Lycopene) 10% FS by DSM Nutritional Products, Inc. UL Prospector. Available online: https://www.ulprospector.com/en/na/Food/Detail/6295/239893/redivivo-Lycopene-10-FS (accessed on 3 September 2020).
- Lyc-O-Mato® -Packed with Powder of Tomato by LycoRed Ltd. NUTRA Ingredients. Available online: https://www.nutraingredients.com/Product-innovations/Lyc-O-Mato-R-Packed-with-the-Power-of-the-Tomato (accessed on 3 September 2020).
- Suppliers for CAS 502-65-8. BuyersGuideChem. Available online: https://www.buyersguidechem.com/AliefAus.php?pnumm=130817126231 (accessed on 2 June 2020).
- Food, Beverage and Nutrition. Barrington Nutritionals. Available online: https://www.ulprospector.com/en/na/Food/Suppliers/2252/Barrington-Nutritionals?st=1 (accessed on 3 September 2020).
- CaroCare®, Beta-Carotene by DSM Nutritional Products, Inc. New Hope Network. Available online: https://www.newhope.com/ingredients-general/carocare-dsm-natural-choice-carotene (accessed on 7 September 2020).
- Lyc-O-Beta 7.5% VBA by LycoRed Ltd. Available online: https://www.lycored.com/lyc-o-beta-7-5-vbaf/ (accessed on 7 September 2020).
- Suppliers for CAS 7235-40-7. BuyersGuideChem. Available online: https://www.buyersguidechem.com/AliefAus.php?pnumm=180131918792 (accessed on 7 September 2020).
- FloraGLO® Lutein 20% SAF by DSM Nutritional Products, Inc. UL Prospector. Available online: https://www.ulprospector.com/en/na/Food/Detail/6295/239891/FloraGLO-Lutein-20-SAF (accessed on 10 August 2020).
- Suppliers for Lutein. BuyersGuideChem. Available online: https://www.buyersguidechem.com/AliefAus.php?pnumm=330278064387&modus=einprod&anzahl_produkte_cas=4 (accessed on 1 July 2020).
- Summary of Color Additives for Use in the United States in Foods, Drugs, Cosmetics, and Medical Devices, U.S. Food and Drug Administration. Available online: https://www.fda.gov/industry/color-additive-inventories/summary-color-additives-use-united-states-foods-drugs-cosmetics-and-medical-devices (accessed on 15 November 2017).
- Global Lycopene Market Forecast Report to 2026: Includes Company Profiling with Detailed Strategies, Financials and Recent Developments—Research and Markets.com. Available online: https://www.businesswire.com/news/home/20190430005701/en/Global-Lycopene-Market-Forecast-Report-2026-Includes (accessed on 30 November 2020).
- Beta Carotene Market Size, Share, Trends, & Industry Analysis Report by Source (Algae, Fruits & Vegetables, & Synthetic); by Application (Food & Beverages, Dietary Supplements, Cosmetics, & Animal Feed); by Regions: Segment Forecast, 2018–2026. Available online: https://www.polarismarketresearch.com/industry-analysis/beta-carotene-market (accessed on 1 December 2020).
- Astaxanthin Market 2020: CAGR of 5.2% with top Countries Data, Latest Trends, Market Size, Share, Global Industry Analysis & Forecast to 2026. Press Release. Available online: https://www.wfmj.com/story/42684950/astaxanthin-market-2020-cagr-of-52-with-top-countries-data-latest-trends-market-size-share-global-industry-analysis-amp-forecast-to-2026 (accessed on 27 September 2020).
- Lutein Market to Reach USD 454.8 million by 2026. Reports and Data. Available online: https://www.prnewswire.com/news-releases/lutein-market-to-reach-usd-454-8-million-by-2026--reports-and-data-300941985.html (accessed on 1 December 2020).
- Fucoxanthin (CAS 3351-86-8) Market 2020 Is Expected to See Magnificent Spike in CAGR with Global Industry Brief Analysis by Top Countries Data Which Includes Driving Factors by Manufacturers Growth and Forecast 2026. Press Release. Available online: https://www.marketwatch.com/press-release/fucoxanthin-cas-3351-86-8-market-2020-is-expected-to-see-magnificent-spike-in-cagr-with-global-industry-brief-analysis-by-top-countries-data-which-includes-driving-factors-by-manufacturers-growth-and-forecast-2026-2020-09-17 (accessed on 2 December 2020).
- Lycopene—Global Market Outlook (2017–2026). Research and Markets. Available online: https://www.researchandmarkets.com/reports/4765062/lycopene-global-market-outlook-2017-2026 (accessed on 1 December 2020).
- Saini, R.K.; Keum, Y.-S. Microbial Platforms to Produce Commercially Vital Carotenoids at Industrial Scale: An Updated Review of Critical Issues. J. Ind. Microbiol. Biotechnol. 2019, 46, 657–674. [Google Scholar] [CrossRef]
- Global Carotenoids Market—Premium Insight, Competitive News Feed Analysis, Company Usability Profiles, Market Sizing & Forecasts to 2025. Available online: https://www.marketresearch.com/360iResearch-v4164/Global-Carotenoids-Premium-Insight-Competitive-13036149/ (accessed on 4 July 2020).
- Venil, C.K.; Dufossé, L.; Devi, P.R. Bacterial Pigments: Sustainable Compounds with Market Potential for Pharma and Food Industry. Front. Sustain. Food Syst. 2020, 4, 100. [Google Scholar] [CrossRef]
- Global Organic Pigments Market Is Segmented by Type (Azo Pigments, Phthalocyanine Pigments, High Performance Pigments (HPPs), Others), by Application (Printing Inks, Paints & Coatings, Plastics, Others (Textiles, Cosmetics, Food)), by Source (Natural Organic Pigments, Synthetic Organic Pigments), and by Region (North America, Latin America, Europe, Asia Pacific, Middle East, and Africa)—Share, Size, Outlook, and Opportunity Analysis, 2019–2026. Available online: https://www.datamintelligence.com/research-report/organic-pigments-market (accessed on 4 December 2020).
- Global Inorganic Pigments Market Is Driven by Rising Demand from End Users and Expanding Paint and Coating Industry. Data Bridge Market Research. Available online: https://www.databridgemarketresearch.com/news/global-inorganic-pigments-market (accessed on 4 December 2020).
- Global Metallic Pigments Market Is Driven by Prevailing Demand of Packaging Applications Mainly in the Food and Beverages, Tobacco Industry, Gifts Wraps among Others. Data Bridge Market Research. Available online: https://www.databridgemarketresearch.com/news/global-metallic-pigments-market (accessed on 5 December 2020).
- Global High-Performance Pigment Market Is Segmented by Type (Organic High-Performance Pigments, In-Organic High-Performance Pigments), by Application (Coatings, Plastics, Cosmetic Products, Others (Includes Inks)), and by Region (North America, Latin America, Europe, Asia Pacific, Middle East, and Africa)—Share, Size, Outlook, and Opportunity Analysis, 2019–2026. Available online: https://www.datamintelligence.com/research-report/high-performance-pigment-market (accessed on 5 December 2020).
- Global Phosphorescent Pigments Market—Industry Trends and Forecast to 2026. Data Bridge Market Research. Available online: https://www.databridgemarketresearch.com/reports/global-phosphorescent-pigments-market (accessed on 6 December 2020).









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pigments | Marine Bacterial Species | References |
|---|---|---|
| Prodigiosin | Hahella chejuensis sp. Pseudoalteromonas rubra sp. Streptomyces sp. SCSIO 11594 Vibrio sp. (Strain MI-2) Serratia marcescens sp. IBRL USM 84 Zooshikella ganghwensis gen. nov., sp. nov. | [22,23,24,25,26,27] |
| Undecylprodigiosin | Streptomyces sp. UKMCC_PT15 Streptomyces sp.SCSIO 11594 Novel strain of Actinobacterium sp., Saccharopolyspora sp. | [24,28,29] |
| Heptylprodigiosin | Spartinivicinus ruber gen. nov., sp. nov | [30] |
| Cycloprodigiosin | Pseudoalteromonas denitrificans sp. Pseudoalteromonas rubra sp. ATCC 29570 | [15,31] |
| Norprodigiosin | Serratia sp. WPRA3 | [32] |
| Astaxanthin | Brevundimonas scallop sp. Zheng & Liu Corynebacterium glutamicum sp. Brevundimonas sp. strain N-5 Sphingomicrobium astaxanthinifaciens sp. nov Rhodovulum sulfidophilum sp. Pontibacter korlensis sp. AG6 Exiguobacterium sp. Altererythrobacter ishigakiensis sp. NBRC 107699 Rhodotorula sp. Paracoccus haeundaensis sp. | [33,34,35,36,37,38,39,40,41,42] |
| Zeaxanthin | Sphingomonas phyllosphaerae sp. KODA19-6 Mesoflavibacter aestuarii sp. nov. Aquibacter zeaxanthinifaciens gen. nov., sp. nov. Zeaxanthinibacter enoshimensis gen. nov., sp. nov Gramella planctonica sp. nov Mesoflavibacter zeaxanthinifaciens gen. nov., sp. nov Formosa sp. KMW Sphingomonas phyllosphaerae sp. FA2T Sphingomonas (Blastomonas) natatoria sp. DSM 3183T Muricauda lutaonensis sp. CC-HSB-11T | [29,43,44,45,46,47,48,49,50] |
| Lycopene | Blastochloris tepida sp. Salinicoccus roseus sp. | [51,52] |
| Beta carotene | Cyanobacterium Synechococcus sp. Micrococcus sp. Vibrio owensii sp. Flavicella marina gen. nov., sp. nov. Gordonia terrae sp.TWRH01 | [53,54,55,56,57] |
| Canthaxanthin | Brevibacterium sp. Gordonia sp. Dietzia sp. | [54] |
| Pyocyanin | Pseudomonas aeruginosa sp. | [58] |
| Scytonemin | Cyanobacterial sp. Nostoc punctiforme sp. ATCC | [59,60] |
| Violacein | Pseudoalteromonas luteoviolacea sp. Pseudoalteromonas ulvae sp. TC14 Pseudoalteromonas sp. 520P1 Microbulbifer sp. D250 Pseudoalteromonas amylolytica sp. nov Chromobacterium violaceum sp. Collimonas sp. | [61,62,63,64,65,66,67] |
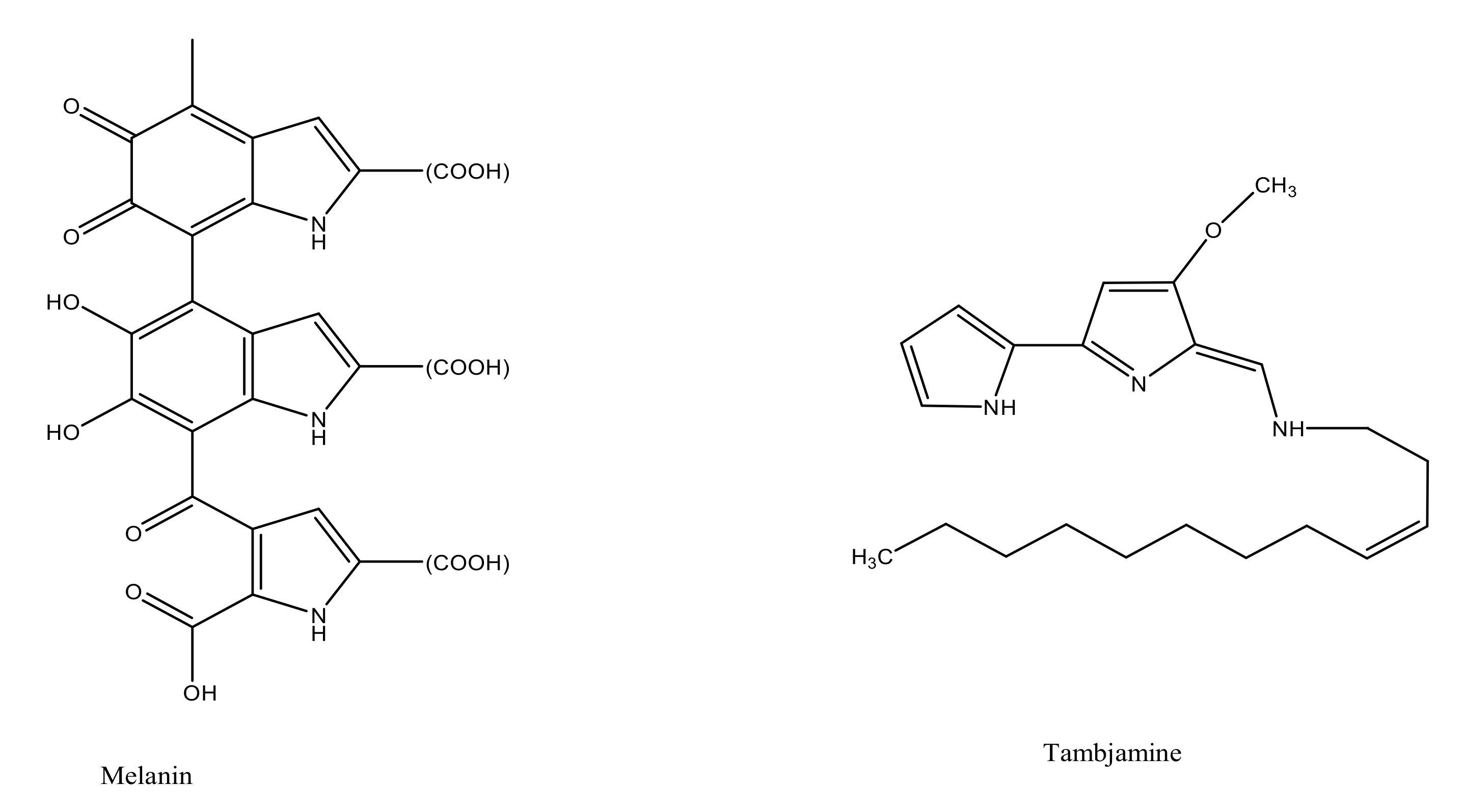
| Melanin | Streptomyces sp. Pseudomonas sp. Marinomonas mediterranea sp. MMB-1T Pseudomonas stutzeri sp. Bacillus sp. BTCZ31 Streptomyces sp. MVCS13 Providencia rettgeri sp. strain BTKKS1 Marinobacter alkaliphilus sp. Leclercia sp. Halomonas meridian sp. Nocardiopsis dassonvillei sp. strain JN1 Vibrio alginolyticus sp. strain BTKKS3 | [68,69,70,71,72,73,74,75,76,77] |
| Tambjamines | Pseudoalteromonas tunicata sp. Pseudoalteromonas citrea sp. | [78,79] |
| Pigments | Marine Bacterial Species | Therapeutic Applications | References |
|---|---|---|---|
| Prodigiosin and cycloprodigiosin | Zooshikella rubidus sp. S1-1 | Antibacterial | [98] |
| Pseudoalteromonas rubra sp. PS1 and SB14 | Antifungal | [23] | |
| Prodigiosin | Serratia rubidaea sp. RAM_Alex | Antiviral | [116] |
| 2-(p-hydroxybenzyl) prodigiosin | Psuedoalteromonas rubra sp. | Cytotoxic | [104] |
| Astaxanthin | Pontibacter korlensis sp. AG6 | Anticancer | [38] |
| 2,2 dihydroxyastaxanthin | Brevundimonas sp. | Antioxidant | [33] |
| Zeaxanthin | Muricauda aquimarina sp. Muricauda olearia sp. | Nitric oxide scavenging Inhibition of lipid peroxidation DPPH radical scavenging activities | [111] |
| Lycopene | Arthrobacter sp. G2O | Antitumor | [108] |
| Beta Carotene | Cyanobacterium sp. | Antioxidant Antidiabetic Antitumor | [118] |
| Pyocyanin and pyorubrin | Psuedomonas aeroginosa sp. | Antibacterial | [58] |
| Melanin | Streptomyces sp. | Antibacterial | [68] |
| Nocardiopsis sp. | Antiquorum sensing | [76] | |
| Pseudomonas stutzeri sp. | Antioxidant | [112] | |
| Poly melanin | Leclercia sp. BTCZ22 | Antibiotic resistance | [75] |
| Tambjamines | Psuedoalteromonas tunicata sp. | Anticancer | [107] |
| Violacein | Janthinobacterium sp. SMN 33.6 | Antibacterial | [102] |
| Collimonas sp. | Antibacterial | [67] | |
| Iodobacter sp. | Antibacterial | [100] | |
| Chromobacterium sp. | Antifungal | [103] | |
| Janthinobacterium sp. UV13 | Anticancer | [106] | |
| 3R Saproxanthin and Myxol | Flavobacteriacae sp. | Antioxidant | [110] |
| PCA | Pseudomonas aeruginosa sp. GS-33 | Effectivity against melanoma cell cancer | [105] |
| Bright pink-orange colored pigment | Salinicoccus sp. | Antibacterial | [101] |
| Yellow pigment | Micrococcus sp. MP76 | Antibacterial | [99] |
| Pigments | Marine Bacterial Species | Industrial Applications | References |
|---|---|---|---|
| Prodigiosin and cycloprodigiosin | Zooshikella rubidus sp. S1-1 | Dyeing potential | [98] |
| Prodigiosin | Vibrio sp. | Dyeing of fabrics | [121] |
| Serratia marcescens sp.11E | Photosensitizers | [131] | |
| Serratia sp. BTWJ8 | Dyeing of paper, PMMA and rubber latex | [124] | |
| Zooshikella sp. | Food colorant Staining | [120] | |
| Serratia marcescens sp. CMST 07 | Antifouling | [129] | |
| Lycopene | Streptomyces sp. | Food grade pigments Feed additive Colorant | [119] |
| Pyocyanin and pyorubrin | Psuedomonas aeroginosa sp. | Food colorings | [58] |
| Scytonemin | Lyngbya aestuarii sp. | Sunscreen | [91] |
| Melanin | Vibrio natriegens sp. | Protection from UV irradiation | [128] |
| Halomonas venusta sp. | Sunscreen Wound healing | [127] | |
| Nocardiopsis sp. | Antibiofilm | [76] | |
| Poly melanin | Pseudoalteromonas lipolytica sp. | Antifouling agent | [130] |
| Vibrio natriegens sp. | Removal of heavy metals and environmental pollutants | [128] | |
| PCA | Pseudomonas aeruginosa sp. GS-33 | Enhance SPF values UV-B protection | [105] |
| Pigments | Products/Nutritional Supplements | Company Coverage/Manufacturers/ Product Brands/Suppliers | References/Links |
|---|---|---|---|
| Pigments from Bacterial Origin | |||
| Prodigiosin | Prodigiosin Serratia marcescens-CAS 82-89-3-CalbiocheM | Sigma-Aldrich | [134] |
| Astaxanthin | Paracoccus Powder (Astaxanthin Powder) 2Z | Brine Shrimp Direct | [135] |
| Paracoccus Powder, Natural Source of Astaxanthin, 50g | NoCoast AQUATICS | [136] | |
| Violacein | Violacein (from Janthinobacterium lividum) | Sigma-Aldrich | [137] |
| Synthetic Pigments and Pigments derived from other sources | |||
| Prodigiosin | Prodigiosin, Antibiotic Prodigiosin 25c | My BioSource Leap Chem Co., Ltd. | [138,139,140] |
| Prodigiosin Suppliers | |||
| Hangzhou Dayangchem Co., Ltd. Puyer Bio Pharm Ltd Santa Cruz Biotechnology | |||
| Astaxanthin | Lucantin® Pink (Astaxanthin) AstaSana™ 10% FS J-BioTMAstaxanthin BioAstin® Hawaiian Astaxanthin | BASF Nutrition DSM GMP Global Marketing, Inc. Cyanotech Corporation | [141,142,143,144,145] |
| Astaxanthin Suppliers | |||
| Aecochem Corp. Simagchem Corporation Hangzhou Dayangchem Co., Ltd. Xiamen Hisunny Chemical Co., Ltd. | |||
| Zeaxanthin | ZeaGold®, Zeaxanthin MacuShield® softgel capsule OPTISHARP® (Zeaxanthin) 20% FS. | Kalsec AGP Limited DSM Nutritional Products, Inc. | [146,147,148,149] |
| Zeaxanthin Suppliers | |||
| Shanghai Worldyang Chemical Co., Ltd. Sancai Industry Co., Ltd. Carbone Scientific Co., Ltd. BLD Pharmatech Ltd. | |||
| Lycopene | redivivo® (Lycopene) 10% FS Lyc-O-Mato® | DSM Nutritional Products, Inc. LycoRed Ltd. | [150,151,152] |
| Lycopene Suppliers | |||
| Haihang Industry Co., Ltd. Shanghai Worldyang Chemical Co., Ltd. Junwee Chemical Co., Ltd. B.M.P. Bulk Medicines & Pharmaceuticals GmbH. | |||
| Beta-Carotene | Beta-Carotene 10% DC Beta-Carotene 1% SD Beta-Carotene 30% in corn oil CaroCare®, Beta-Carotene Lyc-O-Beta 7.5% VBA | Barrington Nutritionals DSM Nutritional Products LycoRed Ltd. | [153,154,155,156] |
| Beta-Carotene Suppliers | |||
| Puyer BioPharma Ltd. Sancai Industry Co., Ltd. United New Materials Technology SDN.BHD. Hangzhou Keying Chem Co., Ltd. | |||
| Lutein | FloraGLO® Lutein 20% SAF | DSM Nutritional Products, Inc. | [157,158] |
| Lutein Suppliers | |||
| Beckmann-Kenko GmbH. New Natural Biotechnology Co., Ltd. BuGuCh & Partners. Stauber Performance Ingredients, Inc. (previous Pharmline, Inc.) | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nawaz, A.; Chaudhary, R.; Shah, Z.; Dufossé, L.; Fouillaud, M.; Mukhtar, H.; ul Haq, I. An Overview on Industrial and Medical Applications of Bio-Pigments Synthesized by Marine Bacteria. Microorganisms 2021, 9, 11. https://doi.org/10.3390/microorganisms9010011
Nawaz A, Chaudhary R, Shah Z, Dufossé L, Fouillaud M, Mukhtar H, ul Haq I. An Overview on Industrial and Medical Applications of Bio-Pigments Synthesized by Marine Bacteria. Microorganisms. 2021; 9(1):11. https://doi.org/10.3390/microorganisms9010011
Chicago/Turabian StyleNawaz, Ali, Rida Chaudhary, Zinnia Shah, Laurent Dufossé, Mireille Fouillaud, Hamid Mukhtar, and Ikram ul Haq. 2021. "An Overview on Industrial and Medical Applications of Bio-Pigments Synthesized by Marine Bacteria" Microorganisms 9, no. 1: 11. https://doi.org/10.3390/microorganisms9010011
APA StyleNawaz, A., Chaudhary, R., Shah, Z., Dufossé, L., Fouillaud, M., Mukhtar, H., & ul Haq, I. (2021). An Overview on Industrial and Medical Applications of Bio-Pigments Synthesized by Marine Bacteria. Microorganisms, 9(1), 11. https://doi.org/10.3390/microorganisms9010011