Combating Parasitic Nematode Infections, Newly Discovered Antinematode Compounds from Marine Epiphytic Bacteria


Abstract
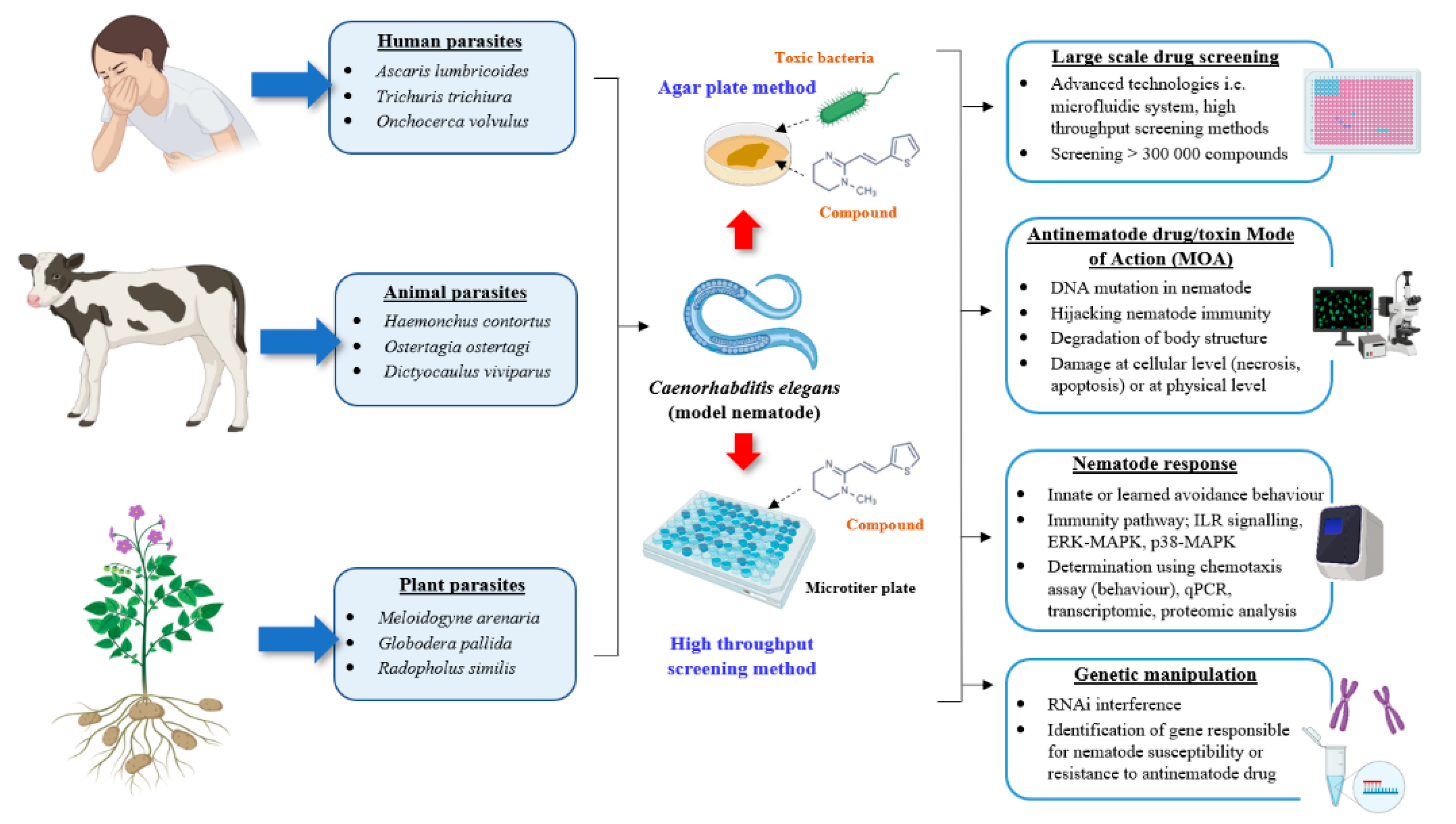
:1. Introduction
2. Antinematode Drug Discovery: Transition from Terrestrial to Marine-Derived Microbial Compounds

{kind=link}
{kind=link}
| Microbial Producer | Compound | Mode of Action | Target Nematode | Affected Nematode Region | Reference |
|---|---|---|---|---|---|
| Bacillus thuringiensis | Crystal toxin Cry5B, Cry21A | Toxin binds to nematode glycoconjugate receptor and disrupt the intestinal cells membrane integrity. This action causes fall of nematode brood size and mortality | Ancylostoma ceylanicum, Ascaris suum, C. elegans | Gastrointestinal system | [70,71,72] |
| Bacillus simplex, B. subtilis, B. weihenstephanensis, Microbacterium oxydans, Stenotrophomonas maltophilia, Streptomyces lateritius and Serratia marcescens | Volatile organic compound (VOC) i.e., benzaldehyde, benzeneacetaldehyde, decanal, 2-nonanone, 2-undecanone, cyclohexene and dimethyl disulfide | VOCs reduce nematode motility and death | Panagrellus redivivus, Bursaphelenchus xylophilus | Unknown | [73] |
| Streptomyces avermectinius | Avermectin and ivermectin (semi-synthetic) | Compound exposure resulted in pharyngeal paralysis and nematode death | Haemonchus contortus, Brugia malayi, C. elegans | Neuromuscular system | [48,74,75,76] |
| Serratia marcescens | Prodigiosin | Compound is toxic against juveniles larvae and inhibit egg hatching competency | Radopholus similis, Meloidogyne javanica | Unknown | [77] |
| Pseudomonas aeruginosa | Phenazine toxin (phenazine-1-carboxylic, pyocyanin and 1-hydroxyphenazine) | Phenazine-1-carboxylic shows fast killing activity against nematode in acidic environment whilst pyocyanin is toxic in neutral or basic pH. The toxicity of 1-hydroxyphenazine is not dependent on environmental pH. Continuous exposure to phenazine affects protein homeostasis and causes neurodegeneration | C. elegans | Neuromuscular system, cell mitochondria and protein folding | [78,79] |
| Pseudomonas aeruginosa | Exotoxin A and other undetermined effectors | Slow-killing activity against nematode is based on infection-like process thus resulting in accumulation of bacteria in the gut. Continuous exposure leads to ceased pharyngeal pumping, nematode immobility and death | C. elegans | Gastrointestinal system | [80] |
| Pseudomonas aeruginosa | Chitinase enzyme | Chitinase degrades nematode cuticle, intestine and egg shell leading to the animal death | C. elegans | Cuticle, eggs, gastrointestinal system | [81] |
| Pseudomonas plecoglossicida | Glycolipid biosurfactant | Reduction of nematode development, survival and fecundity | C. elegans | Unknown | [82] |
3. Surface Associated Marine Bacteria: A Reservoir for Novel Antimicrobial and Antinematode Drug Discovery
| Marine Microbial Producer | Compound | Associated Surface/Host | Mode of Action | Responsible Gene(S) | Reference |
|---|---|---|---|---|---|
| Microbulbifer sp. D250 | Violacein | Algae Delisea pulchra | Facilitate bacterial accumulation accompanied by tissue damage and apoptosis | VioA-VioE | [118] |
| Pseudoalteromonas tunicata D2 | Tambjamine | Algae Ulva australis | Slow-killing activity by a heat-resistant tambjamine and substantial bacterial colonization in the nematode gut | TamA-TamT | [119] |
| Pseudoalteromonas tunicata D2 | Unknown | Algae Ulva australis | Fast-killing activity by a heat sensitive unknown compound through colonization-independent manner | Unknown | [119] |
| Uncultured alpha-proteobacterium, JN874385 (strain U95) | Unknown | Algae Ulva australis | Undetermined | Possibly NRPS gene | [120] |
| Aequorivita sp. | Unknown | Antarctic marine sediment | Undetermined | Unknown | [121] |
| Pseudovibrio sp. Pv348, 1413, HE818384 (strain D323) | Unknown | Algae Delisea pulchra | Undetermined | Unknown | [120] |
| Heterologous clone jj117 (NCBI Accession number SRX4339430) | Unknown | Ulva australis metagenomic library | Undetermined | ATP-grasp protein/alpha-E protein/transglutaminase protein/protease | [122] |
| Vibrio atlanticus strain S-16 | Volatile organic compounds (VOC) | Scallop Argopecten irradians | Undetermined | Unknown | [123] |
| Virgibacillus dokdonensis MCCC 1A00493 | VOC (acetaldehyde, dimethyl disulfide, ethylbenzene and 2-butanone | Polymetallic nodules in the deep sea | Direct contact killing activity, fumigation. Acetaldehyde had a fumigant activity to impede egg hatching | Unknown | [124] |
| Pseudoalteromonas rubra | Unknown | Marine organisms (copepod or fish) or environmental samples | Undetermined | Unknown | [125,126] |
| Pseudoalteromonas piscicida | Unknown | Marine organisms (copepod or fish) or environmental samples | Undetermined | Unknown | [125,126] |
| Arthrobacter davidanieli | Unknown | Marine environmental sample | Undetermined | Unknown | [125,127] |
| Pseudoalteromonas luteoviolacea | Unknown | Marine organisms (copepod or fish) or environmental samples | Undetermined | Unknown | [125,126] |
| Photobacterium halotolerans | Unknown | Marine organisms (copepod or fish) or environmental samples | Undetermined | Unknown | [125,126] |
4. Caenorhabditis elegans as a Model Organism for Antinematode Drug Discovery and Development
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Cox, F.E.G. History of human parasitic diseases. Infect. Dis. Clin. N. Am. 2004, 18, 171–188. [Google Scholar] [CrossRef] [PubMed]
- Bañuls, A.-L.; Thomas, F.; Renaud, F. Of parasites and men. Infect. Genet. Evol. 2013, 20, 61–70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Viney, M. How can we understand the genomic basis of nematode parasitism? Trends Parasitol. 2017, 33, 444–452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garcia-Bustos, J.F.; Sleebs, B.E.; Gasser, R.B. An appraisal of natural products active against parasitic nematodes of animals. Parasites Vectors 2019, 12, 306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ashford, R.; Crewe, W. Parasites of Homo Sapiens: An Annotated Checklist of the Protozoa, Helminths and Arthropods for Which We are Home, 2nd ed.; Taylor & Francis: London, UK; New York, NY, USA, 2003; pp. 87–93. [Google Scholar]
- Stoltzfus, D.J.; Pilgrim, A.A.; Herbert, D.B.R. Perusal of parasitic nematode ‘omics in the post-genomic era. Mol. Biochem. Parasitol. 2017, 215, 11–22. [Google Scholar] [CrossRef] [PubMed]
- Papadi, B.; Eberhard, M.E.; Boudreaux, C.; Bishop, H.; Mathison, B.; Tucker, J.A. Halicephalobus gingivalis: A rare cause of fatal meningoencephalomyelitis in humans. Am. J. Trop. Med. Hyg. 2013, 88, 1062–1064. [Google Scholar] [CrossRef] [Green Version]
- Bryant, A.S.; Ruiz, F.; Gang, S.S.; Castelletto, M.L.; Lopez, J.B.; Hallem, E.A. A critical role for thermosensation in host seeking by skin-penetrating nematodes. Curr. Biol. 2018, 28, 2338–2347.e6. [Google Scholar] [CrossRef] [Green Version]
- Bundy, D.A.; De Silva, N.; Appleby, L.J.; Brooker, S.J. Intestinal nematodes: Ascariasis. In Hunter’s Tropical Medicine and Emerging Infectious Diseases, 10th ed.; Ryan, E.T., Hill, D.R., Solomon, T., Aronson, N.E., Endy, T.P., Eds.; Elsevier: Amsterdam, The Netherlands, 2020; pp. 840–844. [Google Scholar] [CrossRef]
- Jourdan, P.M.; Lamberton, P.H.L.; Fenwick, A.; Addiss, D.G. Soil-transmitted helminth infections. Lancet 2018, 391, 252–265. [Google Scholar] [CrossRef] [Green Version]
- Frean, J. Gnathostomiasis acquired by visitors to the Okavango Delta, Botswana. Trop. Med. Infect. Dis. 2020, 5, 39. [Google Scholar] [CrossRef] [Green Version]
- Eberhard, M.L.; Cleveland, C.A.; Zirimwabagabo, H.; Yabsley, M.J.; Ouakou, P.T.; Ruiz-Tiben, E. Guinea worm (Dracunculus medinensis) infection in a wild-caught frog, Chad. Emerg. Infect. Dis. 2016, 22, 1961–1962. [Google Scholar] [CrossRef]
- Magnino, S.; Colin, P.; Dei-Cas, E.; Madsen, M.; McLauchlin, J.; Nöckler, K.; Maradona, M.P.; Tsigarida, E.; Vanopdenbosch, E.; Van Peteghem, C. Biological risks associated with consumption of reptile products. Int. J. Food Microbiol. 2009, 134, 163–175. [Google Scholar] [CrossRef] [PubMed]
- Persson, G.; Ekmann, J.R.; Hviid, T.V.F. Reflections upon immunological mechanisms involved in fertility, pregnancy and parasite infections. J. Reprod. Immunol. 2019, 136, 102610. [Google Scholar] [CrossRef] [PubMed]
- Ezeamama, A.E.; McGarvey, S.T.; Olveda, R.M.; Acosta, L.P.; Kurtis, J.D.; Bellinger, D.C.; Langdon, G.C.; Manalo, D.L.; Friedman, J.F. Helminth infection and cognitive impairment among Filipino children. Am. J. Trop. Med. Hyg. 2005, 72, 540–548. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doni, N.Y.; Zeyrek, F.Y.; Simsek, Z.; Gurses, G.; Sahin, I. Risk factors and relationship between intestinal parasites and the growth retardation and psychomotor development delays of children in Şanlıurfa, Turkey. Turk. Parazitolojii Derg. 2015, 39, 270–276. [Google Scholar] [CrossRef] [PubMed]
- Abad, P.; Gouzy, J.; Aury, J.; Castagnone-Sereno, P.; Danchin, E.G.J.; Deleury, E.; Perfus-Barbeoch, L.; Anthouard, V.; Artiguenave, F.; Blok, V.C.; et al. Genome sequence of the metazoan plant-parasitic nematode Meloidogyne incognita. Nat. Biotechnol. 2008, 26, 909–915. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bernard, G.C.; Egnin, M.; Bonsi, C. The impact of plant-parasitic nematodes on agriculture and methods of control. In Nematology-Concepts, Diagnosis and Control; Manjur-Shah, M., Mahamood, M., Eds.; IntechOpen: London, UK, 2017. [Google Scholar] [CrossRef] [Green Version]
- Jones, J.T.; Haegeman, A.; Danchin, E.G.J.; Gaur, H.S.; Helder, J.; Jones, M.G.K.; Kikuchi, T.; Manzanilla-López, R.; Palomares-Rius, J.E.; Wesemael, W.M.L.; et al. Top 10 plant-parasitic nematodes in molecular plant pathology. Mol. Plant Pathol. 2013, 14, 946–961. [Google Scholar] [CrossRef]
- Kassie, Y.G.; Ebrahim, A.S.; Mohamed, M.Y. Interaction effect between Meloidogyne incognita and Fusarium oxysporum f. sp. lycopersici on selected tomato (Solanum lycopersicum L.) genotypes. Afr. J. Agric. Res. 2020, 15, 330–342. [Google Scholar] [CrossRef]
- Liu, W.; Park, S.-W. Underground mystery: Interactions between plant roots and parasitic nematodes. Curr. Plant Biol. 2018, 15, 25–29. [Google Scholar] [CrossRef]
- Villate, L.; Morin, E.; Demangeat, G.; Van Helden, M.; Esmenjaud, D. Control of Xiphinema index populations by fallow plants under greenhouse and field conditions. Phytopathology 2012, 102, 627–634. [Google Scholar] [CrossRef] [Green Version]
- Abd-Elgawad, M.; Askary, T.H. Impact of phytonematodes on agriculture economy. In Biocontrol Agents of Phytonematodes; Askary, T.H., Martinelli, P.R.P., Eds.; CAB International Publishing: Oxfordshire, UK; Boston, MA, USA, 2015; pp. 12–17. [Google Scholar]
- Shinn, A.; Pratoomyot, J.; Bron, J.E.; Paladini, G.; Brooker, E.; Brooker, A.J. Economic costs of protistan and metazoan parasites to global mariculture. Parasitology 2015, 142, 196–270. [Google Scholar] [CrossRef]
- Mehrdana, F.; Buchmann, K. Excretory/secretory products of anisakid nematodes: Biological and pathological roles. Acta Vet. Scand. 2017, 59. [Google Scholar] [CrossRef] [Green Version]
- Levsen, A.; Svanevik, C.S.; Cipriani, P.; Mattiucci, S.; Gay, M.; Hastie, L.C.; Bušelić, I.; Mladineo, I.; Karl, H.; Ostermeyer, U.; et al. A survey of zoonotic nematodes of commercial key fish species from major European fishing grounds—Introducing the FP7 PARASITE exposure assessment study. Fish. Res. 2018, 202, 4–21. [Google Scholar] [CrossRef]
- Aibinu, I.E.; Smooker, P.M.; Lopata, A.L. Anisakis nematodes in fish and shellfish- from infection to allergies. Int. J. Parasitol. Parasites Wildl. 2019, 9, 384–393. [Google Scholar] [CrossRef] [PubMed]
- Shamsi, S. Seafood-borne parasitic diseases: A “one-health” approach is needed. Fishes 2019, 4, 9. [Google Scholar] [CrossRef] [Green Version]
- Mushonga, B.; Habumugisha, D.; Kandiwa, E.; Madzingira, O.; Samkange, A.; Segwagwe, B.E.; Jaja, I.F. Prevalence of Haemonchus contortus infections in sheep and goats in Nyagatare District, Rwanda. J. Vet. Med. 2018, 2018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elseadawy, R.; Abbas, I.; Al-Araby, M.; Hildreth, M.B.; Abu-Elwafa, S. First evidence of Teladorsagia circumcincta infection in sheep from Egypt. J. Parasitol. 2019, 105, 484–490. [Google Scholar] [CrossRef]
- McLeod, R. Costs of major parasites to the Australian livestock industries. Int. J. Parasitol. 1995, 25, 1363–1367. [Google Scholar] [CrossRef]
- Roeber, F.; Jex, A.R.; Gasser, R.B. Impact of gastrointestinal parasitic nematodes of sheep, and the role of advanced molecular tools for exploring epidemiology and drug resistance—An Australian perspective. Parasites Vectors 2013, 6, 153. [Google Scholar] [CrossRef] [Green Version]
- Sackett, D.; Sackett, H.; Abbott, K.; Barber, M. Assessing the Economic Cost of Endemic Disease on the Profitability of Australian Beef Cattle and Sheep Producers. Meat & Livestock Australia Report AHW. 2006. Available online: https://www.mla.com.au/Research-and-development/Search-RD-reports/RD-report-details/Animal-Health-and-Biosecurity/Assessing-the-economic-cost-of-endemic-disease-on-the-profitability-of-Australian-beef-cattle-and-sheep-producers/120 (accessed on 16 May 2020).
- McLeod, R.S. Economic impact of worm infestations in small ruminants in South East Asia, India and Austrailia. In Worm Control of Small Ruminants in Tropical Asia; Sani, R.A., Gray, G.D., Baker, R.L., Eds.; ACIAR Monograph: Canberra, Australia, 2004; Volume 113, pp. 23–33. [Google Scholar]
- Maqbool, I.; Wani, Z.A.; Shahardar, R.A.; Allaie, I.M.; Shah, M.M. Integrated parasite management with special reference to gastro-intestinal nematodes. J. Parasit. Dis. 2017, 41, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Rashid, M.; Akbar, H.; Ahmad, L.; Hassan, M.A.; Ashraf, K.; Saeed, K.; Gharbi, M. A systematic review on modelling approaches for economic losses studies caused by parasites and their associated diseases in cattle. Parasitology 2019, 146, 129–141. [Google Scholar] [CrossRef]
- May, K.; Brügemann, K.; König, S.; Strube, C. The effect of patent Dictyocaulus viviparus (re)infections on individual milk yield and milk quality in pastured dairy cows and correlation with clinical signs. Parasites Vectors 2018, 11, 24. [Google Scholar] [CrossRef] [PubMed]
- Abongwa, M.; Martin, R.J.; Robertson, A.P. A brief review on the mode of action of antinematodal drugs. Acta Vet. Beogr. 2017, 67, 137–152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zajíčková, M.; Nguyen, L.T.; Skálová, L.; Stuchlíková, L.R.; Matoušková, P. Anthelmintics in the future: Current trends in the discovery and development of new drugs against gastrointestinal nematodes. Drug Discov. Today 2020, 25, 430–437. [Google Scholar] [CrossRef] [PubMed]
- Fox, N.J.; Marion, G.; Davidson, R.S.; White, P.C.L.; Hutchings, M.R. Climate-driven tipping-points could lead to sudden, high-intensity parasite outbreaks. R. Soc. Open Sci. 2015, 2, 140296. [Google Scholar] [CrossRef] [Green Version]
- Verschave, S.H.; Charlier, J.; Rose, H.; Claerebout, E.; Morgan, E.R. Cattle and nematodes under global change: Transmission models as an ally. Trends Parasitol. 2016, 32, 724–738. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morgan, E.R.; Aziz, N.A.A.; Blanchard, A.; Charlier, J.; Charvet, C.; Claerebout, E.; Geldhof, P.; Greer, A.W.; Hertzberg, H.; Hodgkinson, J.; et al. 100 questions in livestock helminthology research. Trends Parasitol. 2019, 35, 52–71. [Google Scholar] [CrossRef] [Green Version]
- Genchi, C.; Rinaldi, L.; Mortarino, M.; Genchi, M.; Cringoli, G. Climate and Dirofilaria infection in Europe. Vet. Parasitol. 2009, 163, 286–292. [Google Scholar] [CrossRef]
- McIntyre, K.M.; Setzkorn, C.; Hepworth, P.J.; Morand, S.; Morse, A.P.; Baylis, M. Systematic assessment of the climate sensitivity of important human and domestic animals pathogens in Europe. Sci. Rep. 2017, 7, 7134. [Google Scholar] [CrossRef] [Green Version]
- Scott, I.; Pomroy, W.; Kenyon, P.; Smith, G.; Adlington, B.; Moss, A. Lack of efficacy of monepantel against Teladorsagia circumcincta and Trichostrongylus colubriformis. Vet. Parasitol. 2013, 198, 166–171. [Google Scholar] [CrossRef]
- Brom, R.V.D.; Moll, L.; Kappert, C.; Vellema, P. Haemonchus contortus resistance to monepantel in sheep. Vet. Parasitol. 2015, 209, 278–280. [Google Scholar] [CrossRef]
- Ploeger, H.W.; Everts, R.R. Alarming levels of anthelmintic resistance against gastrointestinal nematodes in sheep in the Netherlands. Vet. Parasitol. 2018, 262, 11–15. [Google Scholar] [CrossRef] [PubMed]
- Holden-Dye, L.; Walker, R.J. Anthelmintic drugs and nematocides: Studies in Caenorhabditis elegans. In WormBook: The Online Review of C. elegans Biology; The C. elegans Research Community, Ed.; WormBook, 2014; Available online: http://www.wormbook.org/chapters/www_anthelminticdrugs.2/anthelminticdrugs.2.html (accessed on 24 June 2020).
- Idris, A.O.; Wintola, O.A.; Afolayan, A.J. Helminthiases; prevalence, transmission, host-parasite interactions, resistance to common synthetic drugs and treatment. Heliyon 2019, 5, e01161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elfawal, M.A.; Savinov, S.N.; Aroian, R.V. Drug screening for discovery of broad-spectrum agents for soil-transmitted nematodes. Sci. Rep. 2019, 9, 12347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sekurova, O.N.; Schneider, O.; Zotchev, S.B. Novel bioactive natural products from bacteria via bioprospecting, genome mining and metabolic engineering. Microb. Biotechnol. 2019, 12, 828–844. [Google Scholar] [CrossRef] [Green Version]
- Deng, Q.; Zhou, L.; Luo, M.; Deng, Z.; Zhao, C. Heterologous expression of Avermectins biosynthetic gene cluster by construction of a Bacterial Artificial Chromosome library of the producers. Synth. Syst. Biotechnol. 2017, 2, 59–64. [Google Scholar] [CrossRef]
- Monaghan, R.L.; Tkacz, J.S. Bioactive microbial products: Focus upon mechanism of action. Annu. Rev. Microbiol. 1990, 44, 271–331. [Google Scholar] [CrossRef]
- Giovanelli, F.; Mattellini, M.; Fichi, G.; Flamini, G.; Perrucci, S. In vitro anthelmintic activity of four plant-derived compounds against sheep gastrointestinal nematodes. Vet. Sci. 2018, 5, 78. [Google Scholar] [CrossRef] [Green Version]
- Spiegler, V.; Liebau, E.; Hensel, A. Medicinal plant extracts and plant-derived polyphenols with anthelmintic activity against intestinal nematodes. Nat. Prod. Rep. 2017, 34, 627–643. [Google Scholar] [CrossRef]
- Gaudêncio, S.P.; Pereira, F. Dereplication: Racing to speed up the natural products discovery process. Nat. Prod. Rep. 2015, 32, 779–810. [Google Scholar] [CrossRef]
- Shalaby, H.; Ashry, H.; Saad, M.; Farag, T. In vitro effects of Streptomyces tyrosinase on the egg and dult worm of Toxocara vitulorum. Iran. J. Parasitol. 2020, 15, 67–75. [Google Scholar]
- Bérdy, J. Bioactive microbial metabolites. J. Antibiot. 2005, 58, 1–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiong, Z.-Q.; Wang, J.-F.; Hao, Y.-Y.; Wang, Y. Recent advances in the discovery and development of marine microbial natural products. Mar. Drugs 2013, 11, 700–717. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kong, D.-X.; Jiang, Y.-Y.; Zhang, H.-Y. Marine natural products as sources of novel scaffolds: Achievement and concern. Dug Discov. Today 2010, 15, 884–886. [Google Scholar] [CrossRef] [PubMed]
- Costello, M.J.; Cheung, A.; De Hauwere, N. Surface area and the seabed area, volume, depth, slope, and topographic variation for the world’s seas, oceans, and countries. Environ. Sci. Technol. 2010, 44, 8821–8828. [Google Scholar] [CrossRef] [PubMed]
- Fenical, W.; Jensen, P.R. Developing a new resource for drug discovery: Marine actinomycete bacteria. Nat. Chem. Biol. 2006, 2, 666–673. [Google Scholar] [CrossRef]
- Kanase, H.; Singh, K.N. Marine pharmacology: Potential, challenges, and future in India. J. Med. Sci. 2018, 38, 49–53. [Google Scholar] [CrossRef]
- Flemming, H.-C.; Wuertz, S. Bacteria and archaea on earth and their abundance in biofilms. Nat. Rev. Microbiol. 2019, 17, 247–260. [Google Scholar] [CrossRef]
- Bar-On, Y.M.; Phillips, R.; Milo, R. The biomass distribution on earth. Proc. Natl. Acad. Sci. USA 2018, 115, 6506–6511. [Google Scholar] [CrossRef] [Green Version]
- Magnabosco, C.; Lin, L.-H.; Dong, H.; Bomberg, M.; Ghiorse, W.; Stan-Lotter, H.; Pedersen, K.; Kieft, T.L.; Van Heerden, E.; Onstott, T.C. The biomass and biodiversity of the continental subsurface. Nat. Geosci. 2018, 11, 707–717. [Google Scholar] [CrossRef]
- Pascoal, F.; Magalhães, C.; Costa, R. The link between the ecology of the prokaryotic rare biosphere and its biotechnological potential. Front. Microbiol. 2020, 11, 231. [Google Scholar] [CrossRef] [Green Version]
- Blockley, A.; Elliott, D.R.; Roberts, A.P.; Sweet, M.J. Symbiotic microbes from marine invertebrates: Driving a new era of natural product drug discovery. Diversity 2017, 9, 49. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; Ding, W.; Li, Y.-X.; Tam, C.; Bougouffa, S.; Wang, R.; Pei, B.; Chiang, H.; Leung, P.; Lu, Y.; et al. Marine biofilms constitute a bank of hidden microbial diversity and functional potential. Nat. Commun. 2019, 10, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Conlan, J.V.; Thompson, R.C.A.; Khamlome, B.; Pallant, L.; Fenwick, S.; Elliot, A.; Sripa, B.; Blacksell, S.D.; Vongxay, K. Soil-transmitted helminthiasis in Laos: A community-wide cross-sectional study of humans and dogs in a mass drug administration environment. Am. J. Trop. Med. Hyg. 2012, 86, 624–634. [Google Scholar] [CrossRef] [PubMed]
- Urban, J.F., Jr.; Hu, Y.; Miller, M.M.; Scheib, U.; Yiu, Y.Y.; Aroian, R.V. Bacillus thuringiensis-derived Cry5B has potent anthelmintic activity against Ascaris suum. PLoS Negl. Trop. Dis. 2013, 7, e2263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, J.-Z.; Hale, K.; Carta, L.; Platzer, E.; Wong, C.; Fang, S.-C.; Aroian, R.V. Bacillus thuringiensis crystal proteins that target nematodes. Proc. Natl. Acad. Sci. USA 2003, 100, 2760–2765. [Google Scholar] [CrossRef] [Green Version]
- Gu, Y.-Q.; Mo, M.-H.; Zhou, J.-P.; Zou, C.-S.; Zhang, K.-Q. Evaluation and identification of potential organic nematicidal volatiles from soil bacteria. Soil Biol. Biochem. 2007, 39, 2567–2575. [Google Scholar] [CrossRef]
- Burg, R.W.; Miller, B.M.; Baker, E.E.; Birnbaum, J.; Currie, S.A.; Hartman, R.; Kong, Y.-L.; Monaghan, R.L.; Olson, G.; Putter, I.; et al. Avermectins, new family of potent anthelmintic agents: Producing organism and fermentation. Antimicrob. Agents Chemother. 1979, 15, 361–367. [Google Scholar] [CrossRef] [Green Version]
- Omura, S. Ivermectin: 25 years and still going strong. Int. J. Antimicrob. Agents 2008, 31, 91–98. [Google Scholar] [CrossRef]
- Campbell, C.W. History of avermectin and ivermectin, with notes on the history of other macrocyclic lactone antiparasitic agents. Curr. Pharm. Biotechnol. 2012, 13, 853–865. [Google Scholar] [CrossRef]
- Rahul, S.; Chandrashekhar, P.; Hemant, B.; Chandrakant, N.; Laxmikant, S.; Satish, P. Nematicidal activity of microbial pigment from Serratia marcescens. Nat. Prod. Res. 2014, 28, 1399–1404. [Google Scholar] [CrossRef]
- Cezairliyan, B.; Vinayavekhin, N.; Grenfell-Lee, D.; Yuen, G.J.; Saghatelian, A.; Ausubel, F.M. Identification of Pseudomonas aeruginosa Phenazines that kill Caenorhabditis elegans. PLoS Pathog. 2013, 9, e1003101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ray, A.; Rentas, C.; Caldwell, G.A.; Caldwell, K.A. Phenazine derivatives cause proteotoxicity and stress in C. elegans. Neurosci. Lett. 2015, 584, 23–27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, M.-W.; Mahajan-Miklos, S.; Ausubel, F.M. Killing of Caenorhabditis elegans by Pseudomonas aeruginosa used to model mammalian bacterial pathogenesis. Proc. Natl. Acad. Sci. USA 1999, 96, 715–720. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, L.; Jiang, H.; Cheng, Q.; Chen, J.; Wu, G.; Kumar, A.; Sun, M.; Liu, Z. Enhanced nematicidal potential of the chitinase pachi from Pseudomonas aeruginosa in association with Cry21Aa. Sci. Rep. 2015, 5, 14395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sabarinathan, D.; Amirthalingam, M.; Sabapathy, P.C.; Govindan, S.; Palanisamy, S.; Kathirvel, P. Anthelmintic efficacy of glycolipid biosurfactant produced by Pseudomonas plecoglossicida: An insight from mutant and transgenic forms of Caenorhabditis elegans. Biodegradation 2019, 30, 203–214. [Google Scholar] [CrossRef]
- Rao, D.; Webb, J.S.; Kjelleberg, S. Competitive interactions in mixed-species biofilms containing the marine bacterium Pseudoalteromonas tunicata. Appl. Environ. Microbiol. 2005, 71, 1729–1736. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Welsh, R.M.; Zaneveld, J.R.; Rosales, S.M.; Payet, J.P.; Burkepile, D.E.; Thurber, R.V. Bacterial predation in a marine host-associated microbiome. ISME J. 2016, 10, 1540–1544. [Google Scholar] [CrossRef]
- de Carvalho, C.C. Marine biofilms: A successful microbial strategy with economic implications. Front. Mar. Sci. 2018, 5. [Google Scholar] [CrossRef] [Green Version]
- Chiu, J.M.Y.; Thiyagarajan, V.; Tsoi, M.M.Y.; Qian, P.Y. Qualitative and quantitative changes in marine biofilms as a function of temperature and salinity in summer and winter. Biofilms 2005, 2, 183–195. [Google Scholar] [CrossRef]
- Ortega-Morales, B.O.; Chan-Bacab, M.J.; Rosa-García, S.D.C.D.L.; Camacho-Chab, J.C. Valuable processes and products from marine intertidal microbial communities. Curr. Opin. Biotechnol. 2010, 21, 346–352. [Google Scholar] [CrossRef]
- Antunes, J.; Leão, P.; Vasconcelos, V. Marine biofilms: Diversity of communities and of chemical cues. Environ. Microbiol. Rep. 2019, 11, 287–305. [Google Scholar] [CrossRef]
- Vlamakis, H.; Chai, Y.; Beauregard, P.B.; Losick, R.; Kolter, R. Sticking together: Building a biofilm the Bacillus subtilis way. Nat. Rev. Microbiol. 2013, 11, 157–168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Egan, S.; Harder, T.; Burke, C.; Steinberg, P.; Kjelleberg, S.; Thomas, T. The seaweed holobiont: Understanding seaweed–bacteria interactions. FEMS Microbiol. Rev. 2013, 37, 462–476. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katharios-Lanwermeyer, S.; Xi, C.; Jakubovics, N.; Rickard, A.H. Mini-review: Microbial coaggregation: Ubiquity and implications for biofilm development. Biofouling 2014, 30, 1235–1251. [Google Scholar] [CrossRef] [PubMed]
- Steinberg, P.D.; De Nys, R. Chemical mediation of colonisation of seaweed surfaces. J. Phycol. 2002, 38, 621–629. [Google Scholar] [CrossRef]
- Dang, H.; Lovell, C.R. Microbial surface colonization and biofilm development in marine environments. Microbiol. Mol. Biol. Rev. 2016, 80, 91–138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Armstrong, E.; Yan, L.; Boyd, K.G.; Wright, P.C.; Burgess, J.G. The symbiotic role of marine microbes on living surfaces. Hydrobiologia 2001, 461, 37–40. [Google Scholar] [CrossRef]
- Haas, A.F.; Wild, C. Composition analysis of organic matter released by cosmopolitan coral reef-associated green algae. Aquat. Biol. 2010, 10, 131–138. [Google Scholar] [CrossRef] [Green Version]
- Thomas, T.; Evans, F.F.; Schleheck, D.; Mai-Prochnow, A.; Burke, C.; Penesyan, A.; Dalisay, D.S.; Stelzer-Braid, S.; Saunders, N.F.W.; Johnson, J.; et al. Analysis of the Pseudoalteromonas tunicata genome reveals properties of a surface-associated life style in the marine environment. PLoS ONE 2008, 3, e3252. [Google Scholar] [CrossRef]
- Matz, C.; Webb, J.S.; Schupp, P.J.; Phang, S.Y.; Penesyan, A.; Egan, S.; Steinberg, P.; Kjelleberg, S. Marine biofilm bacteria evade eukaryotic predation by targeted chemical defense. PLoS ONE 2008, 3, e2744. [Google Scholar] [CrossRef] [Green Version]
- Patterson, D.J.; Lee, W.J. Geographic distribution and diversity of free-living heterotrophic flagellates. In The Flagellates: Unity, Diversity and Evolution; Leadbeater, B.S.C., Green, J.C., Eds.; Taylor & Francis: London, UK, 2000; pp. 277–283. [Google Scholar]
- Hisatugo, K.F.; Mansano, A.S.; Seleghim, M.H. Protozoans bacterivory in a subtropical environment during a dry/cold and a rainy/warm season. Braz. J. Microbiol. 2014, 45, 145–151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Corte, D.; Paredes, G.; Yokokawa, T.; Sintes, E.; Herndl, G.J. Differential response of Cafeteria roenbergensis to different bacterial and archaeal prey characteristics. Microb. Ecol. 2019, 78, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Weitere, M.; Erken, M.; Majdi, N.; Arndt, H.; Norf, H.; Reinshagen, M.; Traunspurger, W.; Walterscheid, A.; Wey, J.K. The food web perspective on aquatic biofilms. Ecol. Monogr. 2018, 88, 543–559. [Google Scholar] [CrossRef]
- Moens, T.; Traunspurger, W.; Bergtold, M. Feeding ecology of free-living benthic nematodes. In Freshwater Nematodes. Ecology and Taxonomy; Abebe, E., Traunspurger, W., Andrássy, I., Eds.; CAB International Publishing: Oxfordshire, UK, 2006; pp. 105–131. [Google Scholar]
- Adnan, M.; Alshammari, E.M.; Patel, M.; Ashraf, S.A.; Khan, S.; Hadi, S. Significance and potential of marine microbial natural bioactive compounds against biofilms/biofouling: Necessity for green chemistry. PeerJ 2018, 6, e5049. [Google Scholar] [CrossRef] [PubMed]
- Penesyan, A.; Kjelleberg, S.; Egan, S. Development of novel drugs from marine surface associated microorganisms. Mar. Drugs 2010, 8, 438–459. [Google Scholar] [CrossRef] [Green Version]
- Rocha-Martin, J.; Harrington, C.; Dobson, A.D.W.; O’Gara, F. Emerging strategies and integrated systems microbiology technologies for biodiscovery of marine bioactive compounds. Mar. Drugs 2014, 12, 3516–3559. [Google Scholar] [CrossRef]
- Thomas, T.R.A.; Kavlekar, D.P.; LokaBharathi, P.A. Marine drugs from sponge-microbe association-a review. Mar. Drugs 2010, 8, 1417–1468. [Google Scholar] [CrossRef] [Green Version]
- Proksch, P.; Edrada, R.A.; Ebel, R. Drugs from the seas—current status and microbiological implications. Appl. Microbiol. Biotechnol. 2002, 59, 125–134. [Google Scholar] [CrossRef]
- Oclarit, J.M.; Okada, H.; Ohta, S.; Kaminura, K.; Yamaoka, Y.; Iizuka, T.; Miyashiro, S.; Ikegami, S. Anti-bacillus substance in the marine sponge, Hyatella species, produced by an associated Vibrio species bacterium. Microbios 1994, 78, 7–16. [Google Scholar]
- Kobayashi, M.; Aoki, S.; Gato, K.; Matsunami, K.; Kurosu, M.; Kitagawa, I. Marine natural products. XXXIV. Trisindoline, a new antibiotic indole trimer, produced by a bacterium of Vibrio sp. separated from the marine sponge Hyrtios altum. Chem. Pharm. Bull. 1994, 42, 2449–2451. [Google Scholar] [CrossRef] [Green Version]
- König, G.M.; Kehraus, S.; Seibert, S.F.; Abdel-Lateff, A.; Müller, D. Natural products from marine organisms and their associated microbes. ChemBioChem 2006, 7, 229–238. [Google Scholar] [CrossRef] [PubMed]
- Rinehart, K.L.; Gloer, J.B.; Hughes, R.G.; Renis, H.E.; McGovren, J.P.; Swynenberg, E.B.; Stringfellow, D.A.; Kuentzel, S.L.; Li, L.H. Didemnins: Antiviral and antitumor depsipeptides from a Caribbean tunicate. Science 1981, 212, 933–935. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Kersten, R.D.; Nam, S.-J.; Lu, L.; Al-Suwailem, A.M.; Zheng, H.; Fenical, W.H.; Dorrestein, P.C.; Moore, B.S.; Qian, P.-Y. Bacterial biosynthesis and maturation of the didemnin anti-cancer agents. J. Am. Chem. Soc. 2012, 134, 8625–8632. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsukimoto, M.; Nagaoka, M.; Shishido, Y.; Fujimoto, J.; Nishisaka, F.; Matsumoto, S.; Harunari, E.; Imada, C.; Matsuzaki, T. Bacterial production of the tunicate-derived antitumor cyclic depsipeptide didemnin B. J. Nat. Prod. 2011, 74, 2329–2331. [Google Scholar] [CrossRef]
- Romano, G.; Costantini, M.; Sansone, C.; Lauritano, C.; Ruocco, N.; Ianora, A. Marine microorganisms as a promising and sustainable source of bioactive molecules. Mar. Environ. Res. 2017, 128, 58–69. [Google Scholar] [CrossRef]
- Sunagawa, S.; Coelho, L.P.; Chaffron, S.; Kultima, J.R.; Labadie, K.; Salazar, G.; Djahanschiri, B.; Zeller, G.; Mende, D.R.; Alberti, A.; et al. Structure and function of the global ocean microbiome. Science 2015, 348, 1261359. [Google Scholar] [CrossRef] [Green Version]
- Venter, J.C.; Remington, K.; Heidelberg, J.F.; Halpern, A.L.; Rusch, D.; Eisen, J.A.; Wu, D.; Paulsen, I.T.; Nelson, K.E.; Nelson, W.; et al. Environmental genome shotgun sequencing of the Sargasso Sea. Science 2004, 304, 66–74. [Google Scholar] [CrossRef] [Green Version]
- Rusch, D.B.; Halpern, A.L.; Sutton, G.; Heidelberg, K.B.; Williamson, S.; Yooseph, S.; Wu, D.; Eisen, J.A.; Hoffman, J.M.; Remington, K.; et al. The Sorcerer II Global Ocean Sampling Expedition: Northwest Atlantic through Eastern Tropical Pacific. PLoS Biol. 2007, 5, e77. [Google Scholar] [CrossRef]
- Ballestriero, F.; Daim, M.; Penesyan, A.; Nappi, J.; Schleheck, D.; Bazzicalupo, P.; Di Schiavi, E.; Egan, S. Antinematode activity of violacein and the role of the insulin/IGF-1 pathway in controlling violacein sensitivity in Caenorhabditis elegans. PLoS ONE 2014, 9, e109201. [Google Scholar] [CrossRef] [Green Version]
- Ballestriero, F.; Thomas, T.; Burke, C.; Egan, S.; Kjelleberg, S. Identification of compounds with bioactivity against the nematode Caenorhabditis elegans by a screen based on the functional genomics of the marine bacterium Pseudoalteromonas tunicata D2. Appl. Environ. Microbiol. 2010, 76, 5710–5717. [Google Scholar] [CrossRef] [Green Version]
- Penesyan, A.; Ballestriero, F.; Daim, M.; Kjelleberg, S.; Thomas, T.; Egan, S. Assessing the effectiveness of functional genetic screens for the identification of bioactive metabolites. Mar. Drugs 2013, 11, 40–49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Esposito, F.P.; Ingham, C.; Hurtado-Ortiz, R.; Bizet, C.; Tasdemir, D.; De Pascale, D. Isolation by miniaturized culture chip of an Antarctic bacterium Aequorivita sp. with antimicrobial and anthelmintic activity. Biotechnol. Rep. 2018, 20, e00281. [Google Scholar] [CrossRef] [PubMed]
- Nappi, J. Discovery of Novel Bioactive Metabolites from Marine Epiphytic Bacteria and Assessment of Their Ecological Role. Ph.D. Thesis, The University of New South Wales, Sydney, Australia, February 2019. [Google Scholar]
- Yu, J.; Du, G.; Li, R.; Li, L.; Li, Z.; Zhou, C.; Chen, C.; Guo, D. Nematicidal activities of bacterial volatiles and components from two marine bacteria, Pseudoalteromonas marina strain H-42 and Vibrio atlanticus strain S-16, against the pine wood nematode, Bursaphelenchus xylophilus. Nematology 2015, 17, 1011–1025. [Google Scholar] [CrossRef]
- Huang, D.; Yu, C.; Shao, Z.; Cai, M.-M.; Li, G.-Y.; Zheng, L.; Yu, Z.-N.; Zhang, J. Identification and characterization of nematicidal volatile organic compounds from deep-sea Virgibacillus dokdonensis MCCC 1A00493. Molecules 2020, 25, 744. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neu, A.-K.; Månsson, M.; Gram, L.; Prol-García, M.J. Toxicity of bioactive and probiotic marine bacteria and their secondary metabolites in Artemia sp. and Caenorhabditis elegans as eukaryotic model organisms. Appl. Environ. Microbiol. 2014, 80, 146–153. [Google Scholar] [CrossRef] [Green Version]
- Gram, L.; Melchiorsen, J.; Bruhn, J.B. Antibacterial activity of marine culturable bacteria collected from a global sampling of ocean surface waters and surface swabs of marine organisms. Mar. Biotechnol. 2010, 12, 439–451. [Google Scholar] [CrossRef]
- Wietz, M.; Månsson, M.; Bowman, J.S.; Blom, N.; Ng, Y.; Gram, L. Wide Distribution of closely related, antibiotic-producing Arthrobacter strains throughout the Arctic Ocean. Appl. Environ. Microbiol. 2012, 78, 2039–2042. [Google Scholar] [CrossRef] [Green Version]
- Mathew, M.D.; Mathew, N.D.; Miller, A.; Simpson, M.; Au, V.; Garland, S.; Gestin, M.; Edgley, M.L.; Flibotte, S.; Balgi, A.; et al. Using C. elegans forward and reverse genetics to identify new compounds with anthelmintic activity. PLoS Negl. Trop. Dis. 2016, 10, e0005058. [Google Scholar] [CrossRef]
- Blasco-Costa, I.; Poulin, R. Parasite life-cycle studies: A plea to resurrect an old parasitological tradition. J. Helminthol. 2017, 91, 647–656. [Google Scholar] [CrossRef] [Green Version]
- Burns, A.R.; Luciani, G.M.; Musso, G.; Bagg, R.; Yeo, M.; Zhang, Y.; Rajendran, L.; Glavin, J.; Hunter, R.; Redman, E.; et al. Caenorhabditis elegans is a useful model for anthelmintic discovery. Nat. Commun. 2015, 6, 7485. [Google Scholar] [CrossRef]
- Leung, M.C.K.; Williams, P.L.; Benedetto, A.; Au, C.; Helmcke, K.J.; Aschner, M.; Meyer, J.N. Caenorhabditis elegans: An emerging model in biomedical and environmental toxicology. Toxicol. Sci. 2008, 106, 5–28. [Google Scholar] [CrossRef] [PubMed]
- Queirós, L.; Pereira, J.; Gonçalves, F.J.M.; Pacheco, M.; Aschner, M.; Pereira, P. Caenorhabditis elegans as a tool for environmental risk assessment: Emerging and promising applications for a “nobelized worm”. Crit. Rev. Toxicol. 2019, 49, 411–429. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Park, H.; Liu, J.; Sternberg, P.W. An efficient genome editing strategy to generate putative null mutants in Caenorhabditis elegans using CRISPR/Cas9. G3 2018, 8, 3607–3616. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Riddle, D.L.; Blumenthal, T.; Meyer, B.J.; Priess, J.R. C elegans II, 2nd ed.; Cold Spring Harbor Laboratory Press: New York, NY, USA, 1997; Section I, The Biological Model. Available online: https://www.ncbi.nlm.nih.gov/books/NBK20086/ (accessed on 5 July 2020).
- Kong, C.; Eng, S.-A.; Lim, M.-P.; Nathan, S. Beyond traditional antimicrobials: A Caenorhabditis elegans model for discovery of novel anti-infectives. Front. Microbiol. 2016, 7, 1956. [Google Scholar] [CrossRef]
- Frézal, L.; Félix, M.-A. The natural history of model organisms: C. elegans outside the Petri dish. eLife 2015, 4, e05849. [Google Scholar] [CrossRef]
- Artal-Sanz, M.; de Jong, L.; Tavernarakis, N. Caenorhabditis elegans: A versatile platform for drug discovery. Biotechnol. J. 2006, 1, 1405–1418. [Google Scholar] [CrossRef]
- Lok, J.B.; Unnasch, T.R. Transgenesis in Animal Parasitic Nematodes: Strongyloides spp. and Brugia spp. In WormBook: The Online Review of C. elegans Biology; The C. elegans Research Community, Ed.; WormBook, 2018. Available online: https://www.ncbi.nlm.nih.gov/books/NBK174830/ (accessed on 23 June 2020).
- Stiernagle, T. Maintenance of C. elegans. In WormBook: The Online Review of C. elegans Biology; The C. elegans Research Community, Ed.; WormBook, 2006; Available online: http://www.wormbook.org/chapters/www_strainmaintain/strainmaintain.html (accessed on 30 May 2020).
- Brenner, S. The genetics of Caenorhabditis elegans. Genetics 1974, 77, 71–94. [Google Scholar]
- Fraietta, I.; Gasparri, F. The development of high-content screening (HCS) technology and its importance to drug discovery. Expert Opin. Drug Discov. 2016, 11, 501–514. [Google Scholar] [CrossRef]
- Midkiff, D.; San-Miguel, A. Microfluidic technologies for high throughput screening through sorting and on-chip culture of C. elegans. Molecules 2019, 24, 4292. [Google Scholar] [CrossRef] [Green Version]
- Driscoll, M.; Dean, E.; Reilly, E.; Bergholz, E.; Chalfie, M. Genetic and molecular analysis of a Caenorhabditis elegans beta-tubulin that conveys benzimidazole sensitivity. J. Cell Biol. 1989, 109, 2993–3003. [Google Scholar] [CrossRef] [Green Version]
- Haber, C.L.; Heckaman, C.L.; Li, G.P.; Thompson, D.P.; Whaley, H.A.; Wiley, V.H. Development of a mechanism of action-based screen for anthelmintic microbial metabolites with avermectin like activity and isolation of milbemycin-producing Streptomyces strains. Antimicrob. Agents Chemother. 1991, 35, 1811–1817. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marroquin, L.D.; Elyassnia, D.; Griffitts, J.S.; Feitelson, J.S.; Aroian, R.V. Bacillus thuringiensis (Bt) toxin susceptibility and isolation of resistance mutants in the nematode Caenorhabditis elegans. Genetics 2000, 155, 1693–1699. [Google Scholar]
- Mitreva, M.; McCarter, J.P.; Martin, J.; Dante, M.; Wylie, T.; Chiapelli, B.; Pape, D.; Clifton, S.W.; Nutman, T.B.; Waterston, R.H. Comparative genomics of gene expression in the parasitic and free-living nematodes Strongyloides stercoralis and Caenorhabditis elegans. Genome Res. 2004, 14, 209–220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumarasingha, R.; Young, N.D.; Manurung, R.; Lim, D.S.L.; Tu, C.-L.; Palombo, E.A.; Shaw, J.M.; Gasser, R.B.; Boag, P.R. Transcriptional alterations in Caenorhabditis elegans following exposure to an anthelmintic fraction of the plant Picria fel-terrae Lour. Parasites Vectors 2019, 12, 181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mir, D.A.; Balamurugan, K. Global proteomic response of Caenorhabditis elegans against PemKSa toxin. Front. Cell. Infect. Microbiol. 2019, 9, 172. [Google Scholar] [CrossRef] [PubMed]
- Angstadt, J.D.; Donmoyer, J.E.; Stretton, A.O. Retrovesicular ganglion of the nematode Ascaris. J. Comp. Neurol. 1989, 284, 374–388. [Google Scholar] [CrossRef] [PubMed]
- Johnson, C.D.; Stretton, A.O. Localization of choline acetyltransferase within identified motoneurons of the nematode Ascaris. J. Neurosci. 1985, 5, 1984–1992. [Google Scholar] [CrossRef] [Green Version]
- Johnson, C.D.; Stretton, A.O. GABA-immunoreactivity in inhibitory motor neurons of the nematode Ascaris. J. Neurosci. 1987, 7, 223–235. [Google Scholar] [CrossRef] [Green Version]
- Davis, R. Neurophysiology of glutamatergic signalling and anthelmintic action in Ascaris suum: Pharmacological evidence for a kainate receptor. Parasitology 1998, 116, 471–486. [Google Scholar] [CrossRef]
- Weeks, J.C.; Robinson, K.J.; Lockery, S.R.; Roberts, W.M. Anthelmintic drug actions in resistant and susceptible C. elegans revealed by electrophysiological recordings in a multichannel microfluidic device. Int. J. Parasitol. Drugs Drug Resist. 2018, 8, 607–628. [Google Scholar] [CrossRef]
- Lewis, J.; Wu, C.-H.; Levine, J.; Berg, H. Levamisole-resitant mutants of the nematode Caenorhabditis elegans appear to lack pharmacological acetylcholine receptors. Neuroscience 1980, 5, 967–989. [Google Scholar] [CrossRef]
- Kaminsky, R.; Ducray, P.; Jung, M.; Clover, R.; Rufener, L.; Bouvier, J.; Weber, S.S.; Wenger, A.; Wieland-Berghausen, S.; Goebel, T.; et al. A new class of anthelmintics effective against drug-resistant nematodes. Nature 2008, 452, 176–180. [Google Scholar] [CrossRef] [PubMed]
- Yamazaki, M.; Okuyama, E.; Kobayashi, M.; Inoue, H. The structure of paraherquamide, a toxic metabolite from Penicillium paraherquei. Tetrahedron Lett. 1981, 22, 135–136. [Google Scholar] [CrossRef]
- Hernando, G.; Turani, O.; Bouzat, C. Caenorhabditis elegans muscle Cys-loop receptors as novel targets of terpenoids with potential anthelmintic activity. PLoS Negl. Trop. Dis. 2019, 13, e0007895. [Google Scholar] [CrossRef] [PubMed]
- Schaeffer, J.M.; Blizzard, T.A.; Ondeyka, J.; Goegelman, R.; Sinclair, P.J.; Mrozik, H. [3H]Paraherquamide binding to Caenorhabditis elegans: Studies on a potent new anthelmintic agent. Biochem. Pharmacol. 1992, 43, 679–684. [Google Scholar] [CrossRef]
- Ruiz-Lancheros, E.; Viau, C.; Walter, T.N.; Francis, A.; Geary, T. Activity of novel nicotinic anthelmintics in cut preparations of Caenorhabditis elegans. Int. J. Parasitol. 2011, 41, 455–461. [Google Scholar] [CrossRef] [PubMed]
- Guo, Z. The modification of natural products for medical use. Acta Pharm. Sin. B 2017, 7, 119–136. [Google Scholar] [CrossRef] [Green Version]
- Taman, A.; Azab, M. Present-day anthelmintics and perspectives on future new targets. Parasitol. Res. 2014, 113, 2425–2433. [Google Scholar] [CrossRef]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Salikin, N.H.; Nappi, J.; Majzoub, M.E.; Egan, S. Combating Parasitic Nematode Infections, Newly Discovered Antinematode Compounds from Marine Epiphytic Bacteria. Microorganisms 2020, 8, 1963. https://doi.org/10.3390/microorganisms8121963
Salikin NH, Nappi J, Majzoub ME, Egan S. Combating Parasitic Nematode Infections, Newly Discovered Antinematode Compounds from Marine Epiphytic Bacteria. Microorganisms. 2020; 8(12):1963. https://doi.org/10.3390/microorganisms8121963
Chicago/Turabian StyleSalikin, Nor Hawani, Jadranka Nappi, Marwan E. Majzoub, and Suhelen Egan. 2020. "Combating Parasitic Nematode Infections, Newly Discovered Antinematode Compounds from Marine Epiphytic Bacteria" Microorganisms 8, no. 12: 1963. https://doi.org/10.3390/microorganisms8121963
APA StyleSalikin, N. H., Nappi, J., Majzoub, M. E., & Egan, S. (2020). Combating Parasitic Nematode Infections, Newly Discovered Antinematode Compounds from Marine Epiphytic Bacteria. Microorganisms, 8(12), 1963. https://doi.org/10.3390/microorganisms8121963