Investigation of In-Vitro Adaptation toward Sodium Bituminosulfonate in Staphylococcus aureus




Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains
2.2. Preparation of Bi-Layered Linear Gradient Agar Plates
2.3. Exposure to Sub-Inhibitory Concentrations to Select the Least Susceptible Phenotypes
2.4. Antimicrobial Susceptibility Testing
2.5. Genetic Analysis of Fusidic Acid Resistance
3. Results
3.1. Baseline Assessment of MIC Values
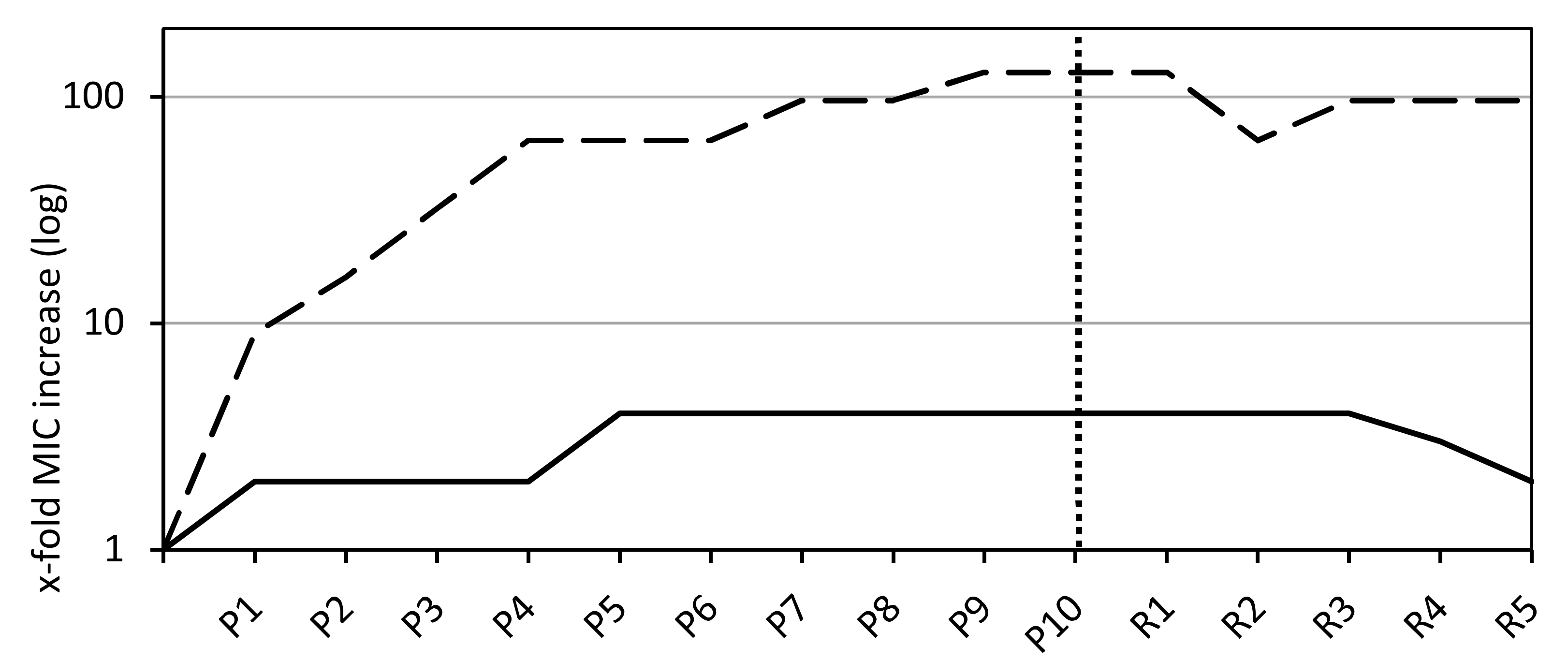
3.2. Exposure to Sub-Inhibitory Concentrations to Select Least Susceptible Phenotypes
3.3. Analyzing the Stability of Phenotypes with Increased MICs in Recovery Passages
3.4. Genetic Mechanisms of the Fusidic Acid Resistance
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Wernli, D.; Jørgensen, P.S.; Harbarth, S.; Carroll, S.P.; Laxminarayan, R.; Levrat, N.; Røttingen, J.-A.; Pittet, D. Antimicrobial resistance: The complex challenge of measurement to inform policy and the public. PLoS Med. 2017, 14, e1002378. [Google Scholar] [CrossRef] [PubMed]
- Fair, R.J.; Tor, Y. Antibiotics and bacterial resistance in the 21st century. Perspect. Medicin. Chem. 2014, 6, 25–64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shrivastava, S.R.; Shrivastava, P.S.; Ramasamy, J. World health organization releases global priority list of antibiotic-resistant bacteria to guide research, discovery, and development of new antibiotics. J. Med. Soc. 2018, 32, 76–77. [Google Scholar] [CrossRef]
- Ventola, C.L. The antibiotic resistance crisis: Part 1: Causes and threats. Pharm. Ther. 2015, 40, 277–283. [Google Scholar]
- Theuretzbacher, U.; Gottwalt, S.; Beyer, P.; Butler, M.; Czaplewski, L.; Lienhardt, C. Analysis of the clinical antibacterial and antituberculosis pipeline. Lancet Infect. Dis. 2018, 19, e40–e50. [Google Scholar] [CrossRef]
- Pulcini, C.; Mohrs, S.; Beovic, B.; Gyssens, I.; Theuretzbacher, U.; Cars, O. Forgotten antibiotics: A follow-up inventory study in Europe, the USA, Canada and Australia. Int. J. Antimicrob. Agents 2017, 49, 98–101. [Google Scholar] [CrossRef]
- Theuretzbacher, U.; Van Bambeke, F.; Canton, R.; Giske, C.G.; Mouton, J.W.; Nation, R.L. Reviving old antibiotics. J. Antimicrob. Chemother. 2015, 70, 2177–2181. [Google Scholar] [CrossRef] [Green Version]
- Unna, P.G. Aphorismen über Schwefeltherapie und Schwefelpräparate. IV. Ichthyol. Mon. Prakt. Dermatol. 1882, 1, 328–333. (In German) [Google Scholar]
- Wernicke, E.A. A Process for the Preparation of Sulfur-Rich, Bright Colored Sulfonation of Sulfur-Rich Mineral Oils, Tar Oils or Schieferoelen. Patent DE624317C, 23 January 1936. [Google Scholar]
- Wernicke, E.A. Schieferöl-Präparate. Münchener Med. Wochenzeitschr. 1936, 13, 522. (In German) [Google Scholar]
- Boyd, A.S. Ichthammol revisited. Int. J. Dermatol. 2010, 49, 757–760. [Google Scholar] [CrossRef]
- Latteux, P. Bakteriologische Untersuchungen, die antiseptischen Eigenschaften des Ichthyols betreffend. Mon. Prakt. Dermatol. 1892, 14, 389–397. [Google Scholar]
- Nilssen, E.; Wormald, P.J.; Oliver, S. Glycerol and ichthammol: Medicinal solution or mythical potion? J. Laryngol. Otol. 1996, 110, 319–321. [Google Scholar] [CrossRef] [PubMed]
- Pantke, R. Zur Kenntnis der Wirkung von Seefelder Schieferölprodukten auf Staphylococcen und Streptococcen (Action of Seefeld slate oil products on staphylococci and streptococci). Arzneim. Forsch. Drug Res. 1951, 1, 415–416. (In German) [Google Scholar]
- Pantke, R. Bakteriologische Untersuchung von Arzneimitteln aus Schieferöl (Bacteriological studies of drugs from shale oil). Arzneim. Forsch. Drug Res. 1965, 15, 570–573. (In German) [Google Scholar]
- Idelevich, E.A.; Becker, K. In vitro activity of sodium bituminosulfonate: Susceptibility data for the revival of an old antimicrobial. Microb. Drug Resist. 2020, 26, 1405–1409. [Google Scholar] [CrossRef] [PubMed]
- Unna, P.G. Über Ichthyol. Mon. Prakt. Dermatol. 1897, 25, 534–539. (In German) [Google Scholar]
- Unna, P.G. Ichthyol. Br. Med. J. 1887, 1, 800. [Google Scholar]
- Baumann, E.; Schotten, C. Über das “Ichthyol”. Mon. Prakt. Dermatol. 1883, 2, 257–262. (In German) [Google Scholar]
- Williamson, D.A.; Carter, G.P.; Howden, B.P. Current and Emerging Topical Antibacterials and Antiseptics: Agents, Action, and Resistance Patterns. Clin. Microbiol. Rev. 2017, 30, 827–860. [Google Scholar] [CrossRef] [Green Version]
- Punjataewakupt, A.; Napavichayanun, S.; Aramwit, P. The downside of antimicrobial agents for wound healing. Eur. J. Clin. Microbiol. Infect. Dis. 2019, 38, 39–54. [Google Scholar] [CrossRef]
- Desroches, M.; Potier, J.; Laurent, F.; Bourrel, A.S.; Doucet-Populaire, F.; Decousser, J.W.; Microbs Study Group. Prevalence of mupirocin resistance among invasive coagulase-negative staphylococci and methicillin-resistant Staphylococcus aureus (MRSA) in France: Emergence of a mupirocin-resistant MRSA clone harbouring mupA. J. Antimicrob. Chemother. 2013, 68, 1714–1717. [Google Scholar] [CrossRef] [PubMed]
- Elazhari, M.; Abu-Quatouseh, L.F.; Elhabchi, D.; Zerouali, K.; Dersi, N.; Saile, R.; Timinouni, M.; Becker, K. Characterization of fusidic acid-resistant Staphylococcus aureus isolates in the community of Casablanca (Morocco). Int. J. Med. Microbiol. 2012, 302, 96–100. [Google Scholar] [CrossRef] [PubMed]
- Von Eiff, C.; Becker, K.; Machka, K.; Stammer, H.; Peters, G. Nasal carriage as a source of Staphylococcus aureus bacteremia. N. Eng. J. Med. 2001, 344, 11–16. [Google Scholar] [CrossRef] [PubMed]
- Schaumburg, F.; Kock, R.; Mellmann, A.; Richter, L.; Hasenberg, F.; Kriegeskorte, A.; Friedrich, A.W.; Gatermann, S.; Peters, G.; Von Eiff, C.; et al. Population Dynamics among Methicillin-Resistant Staphylococcus aureus Isolates in Germany during a 6-Year Period. J. Clin. Microbiol. 2012, 50, 3186–3192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Alen, S.; Ballhausen, B.; Peters, G.; Friedrich, A.W.; Mellmann, A.; Köck, R. In the centre of an epidemic: Fifteen years of LA-MRSA CC398 at the University Hospital Münster. Vet. Microbiol. 2017, 200, 19–24. [Google Scholar] [CrossRef] [PubMed]
- Kriegeskorte, A.; Ballhausen, B.; Idelevich, E.A.; Köck, R.; Friedrich, A.W.; Karch, H.; Peters, G.; Becker, K. Human MRSA Isolates with Novel Genetic Homolog, Germany. Emerg. Infect. Dis. 2012, 18, 1016–1018. [Google Scholar] [CrossRef]
- Clinical and Laboratory Standards Institute (CLSI). Methods for Dilution Antimicrobial Susceptibility Tests for Bacteria that Grow Aerobically, 11th ed.; Approved Standard; M07-A11; CLSI: Wayne, PA, USA, 2018. [Google Scholar]
- Clinical and Laboratory Standards Institute (CLSI). Methods for Antimicrobial Susceptibility Testing of Anaerobic Bacteria, 9th ed.; Approved Standard; M11-A9; CLSI: Wayne, PA, USA, 2018. [Google Scholar]
- ISO. International Standard. ISO 20776-1:2006(E). Clinical Laboratory Testing and in Vitro Diagnostic Test Systems—Susceptibility Testing of Infectious Agents and Evaluation of Performance of Antimicrobial Susceptibility Test Devices—Part 1: Reference Method for Testing the in Vitro Activity of Antimicrobial Agents against Rapidly Growing Aerobic Bacteria Involved in Infectious Diseases; ISO: Geneva, Switzerland, 2006. [Google Scholar]
- Chen, C.M.; Huang, M.; Chen, H.F.; Ke, S.C.; Li, C.R.; Wang, J.H.; Wu, L.T. Fusidic acid resistance among clinical isolates of methicillin-resistant Staphylococcus aureus in a Taiwanese hospital. BMC Microbiol. 2011, 11, 98. [Google Scholar] [CrossRef] [Green Version]
- Drago, L.; De Vecchi, E.; Nicola, L.; Legnani, D.; Prenna, M. In vitro selection of resistance to clarithromycin in Streptococcus pneumoniae clinical isolates. J. Chemother. 2005, 17, 161–168. [Google Scholar] [CrossRef]
- De Vecchi, E.; Nicola, L.; Ossola, F.; Drago, L. In vitro selection of resistance in Streptococcus pneumoniae at in vivo fluoroquinolone concentrations. J. Antimicrob. Chemother. 2009, 63, 721–727. [Google Scholar] [CrossRef] [Green Version]
- Drago, L.; Nicola, L.; Mattina, R.; De Vecchi, E. In vitro selection of resistance in Escherichia coli and Klebsiella spp. at in vivo fluoroquinolone concentrations. BMC Microbiol. 2010, 10, 119. [Google Scholar] [CrossRef] [Green Version]
- Barber, M.; Waterworth, P.M. Antibacterial activity in vitro of fucidin. Lancet 1962, 1, 931–932. [Google Scholar] [CrossRef]
- Godtfredsen, W.; Roholt, K.; Tybring, L. Fucidin: A new orally active antibiotic. Lancet 1962, 1, 928–931. [Google Scholar] [CrossRef]
- Hilson, G.R. In-vitro studies of a new antibiotic (fucidin). Lancet 1962, 1, 932–933. [Google Scholar] [CrossRef]
- O’Neill, A.J.; Cove, J.H.; Chopra, I. Mutation frequencies for resistance to fusidic acid and rifampicin in Staphylococcus aureus. J. Antimicrob. Chemother. 2001, 47, 647–650. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sunderkötter, C.; Becker, K. Frequent bacterial skin and soft tissue infections: Diagnostic signs and treatment. J. Dtsch. Dermatol. Ges. 2015, 13, 501–524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- The European Committee on Antimicrobial Susceptibility Testing. Breakpoint Tables for Interpretation of MICs and Zone Diameters. Version 10.0. 2020. Available online: http://www.eucast.org (accessed on 3 March 2020).
- Laurberg, M.; Kristensen, O.; Martemyanov, K.; Gudkov, A.T.; Nagaev, I.; Hughes, D.; Liljas, A. Structure of a mutant EF-G reveals domain III and possibly the fusidic acid binding site. J. Mol. Biol. 2000, 303, 593–603. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Besier, S.; Ludwig, A.; Brade, V.; Wickelhaus, T.A. Molecular analysis of fusidic acid resistance in Staphylococcus aureus. Mol. Microbiol. 2003, 47, 463–469. [Google Scholar] [CrossRef]
- O’Neill, A.J.; McLaws, F.; Kahlmeter, G.; Henriksen, A.S.; Chopra, I. Genetic basis of resistance to fusidic acid in staphylococci. Antimicrob. Agents Chemother. 2007, 51, 1737–1740. [Google Scholar] [CrossRef] [Green Version]
- Brown, E.M.; Wise, R. Fusidic acid cream for impetigo. Fusidic acid should be used with restraint. BMJ 2002, 324, 1394. [Google Scholar] [CrossRef]


{kind=link}
| MIC Values | x-Fold MIC Increase | |||||
|---|---|---|---|---|---|---|
| Baseline | After 10 Passages | After 5 Recovery Passages | After 10 Passages | After 5 Recovery Passages | ||
| Sodium bituminosulfonate (n = 30) | MIC90 | 0.25 g/L | 1 g/L | 0.5 g/L | - | - |
| Median | 0.125 g/L | 1 g/L | 0.25 g/L | 4 | 2 | |
| Range | 0.06–0.25 g/L | 0.25–2 g/L | 0.125–1 g/L | 2–16 | 1–8 | |
| Fusidic acid (n = 6) | MIC90 | 0.125 mg/L | 16 mg/L | 16 mg/L | - | - |
| Median | 0.094 mg/L | 12 mg/L | 8 mg/L | 128 | 96 | |
| Range | 0.06–0.125 mg/L | 1–16 mg/L | 2–16 mg/L | 16–256 | 32–256 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Blisse, M.; Idelevich, E.A.; Becker, K. Investigation of In-Vitro Adaptation toward Sodium Bituminosulfonate in Staphylococcus aureus. Microorganisms 2020, 8, 1962. https://doi.org/10.3390/microorganisms8121962
Blisse M, Idelevich EA, Becker K. Investigation of In-Vitro Adaptation toward Sodium Bituminosulfonate in Staphylococcus aureus. Microorganisms. 2020; 8(12):1962. https://doi.org/10.3390/microorganisms8121962
Chicago/Turabian StyleBlisse, Marko, Evgeny A. Idelevich, and Karsten Becker. 2020. "Investigation of In-Vitro Adaptation toward Sodium Bituminosulfonate in Staphylococcus aureus" Microorganisms 8, no. 12: 1962. https://doi.org/10.3390/microorganisms8121962
APA StyleBlisse, M., Idelevich, E. A., & Becker, K. (2020). Investigation of In-Vitro Adaptation toward Sodium Bituminosulfonate in Staphylococcus aureus. Microorganisms, 8(12), 1962. https://doi.org/10.3390/microorganisms8121962