Spent Coffee Grounds Alter Bacterial Communities in Latxa Dairy Ewes

 , , , ,
, , , ,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Spent Coffee Grounds Collection and Drying
2.2. Experimental Design, Animals and Feeding
2.3. Samplings and Measurements
2.3.1. Sampling of Ruminal Contents
2.3.2. DNA Extraction and Illumina Library Generation
2.4. Chemical Analyses
2.5. Calculations and Statistical Analysis
3. Results
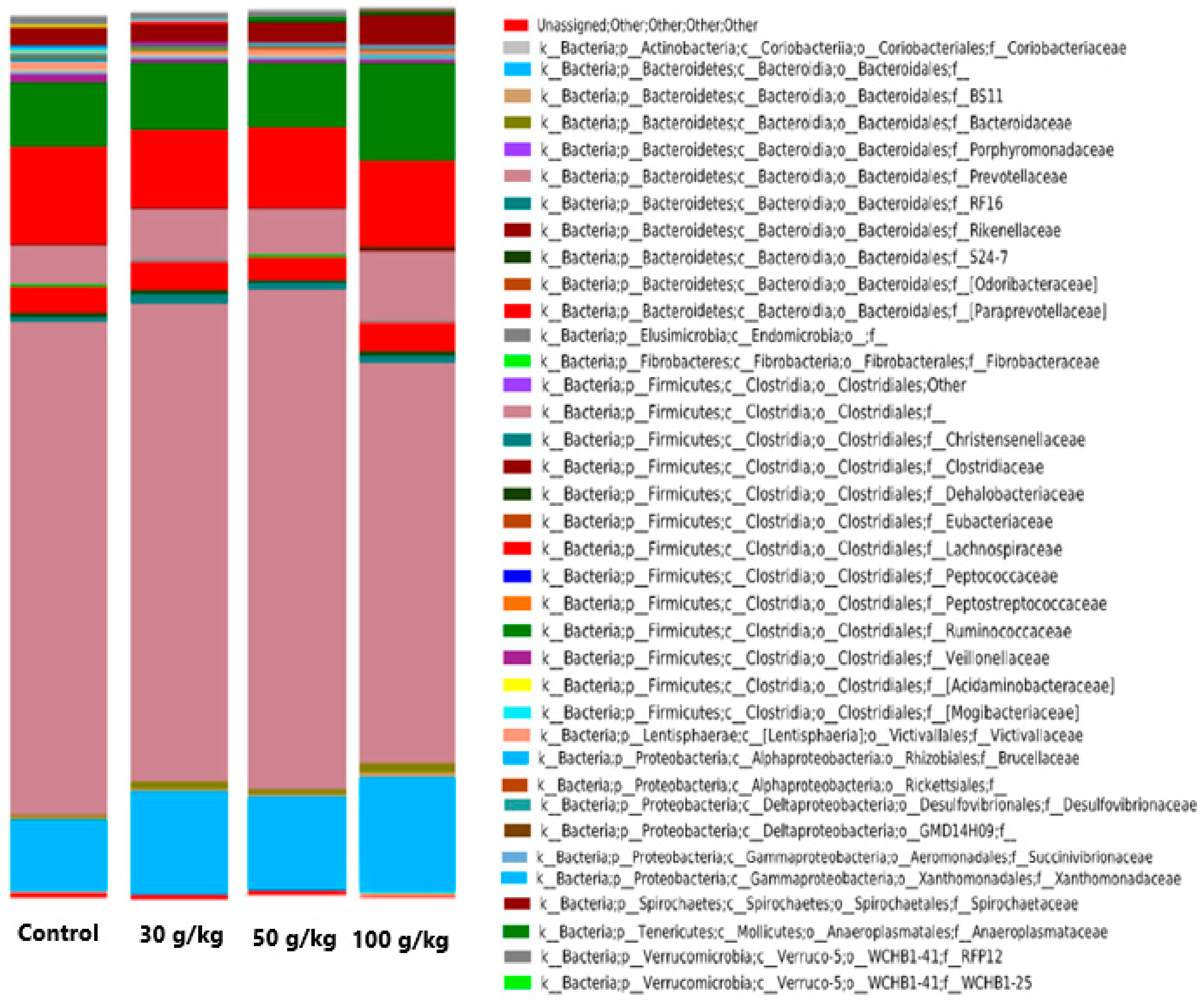
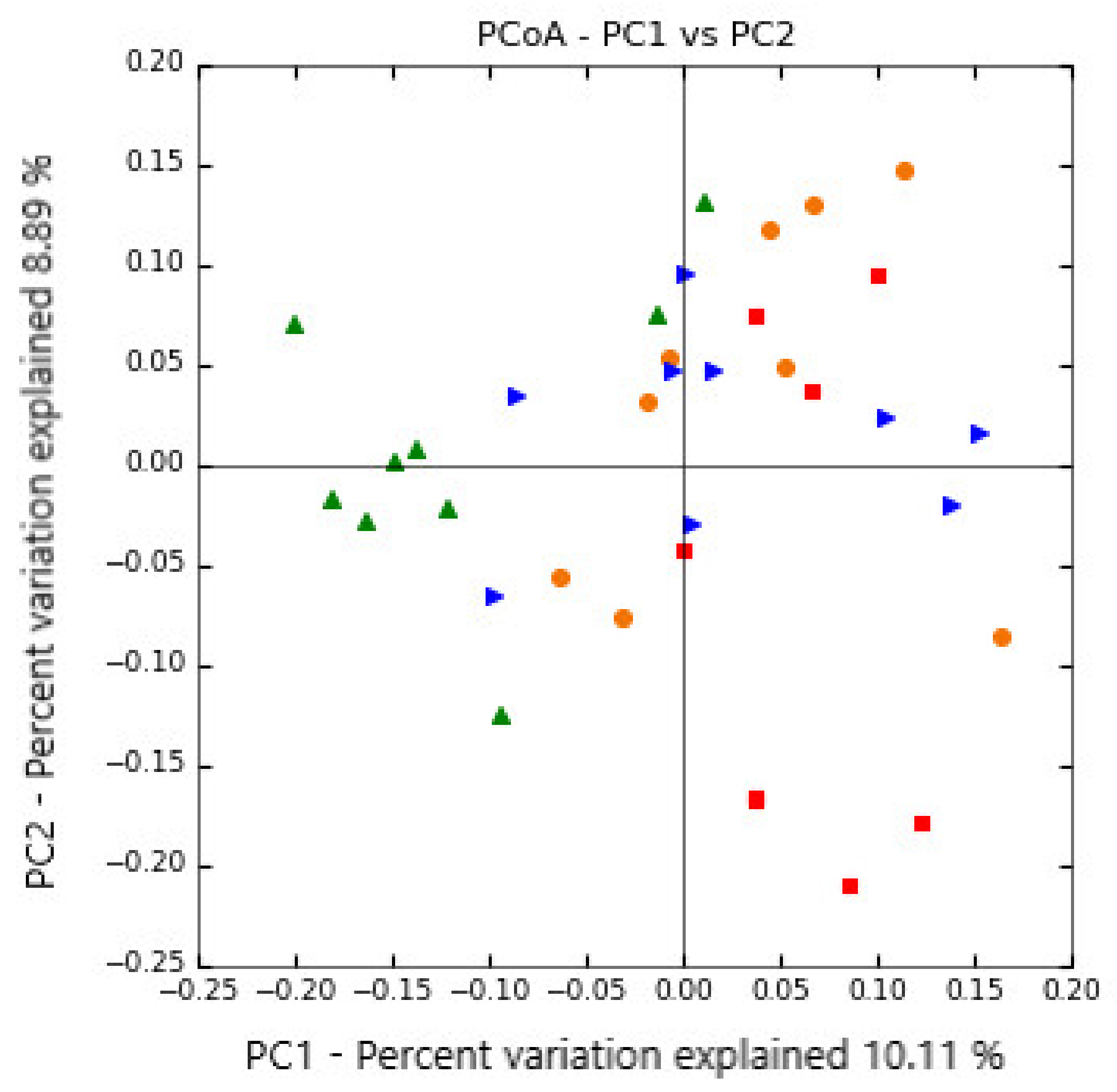
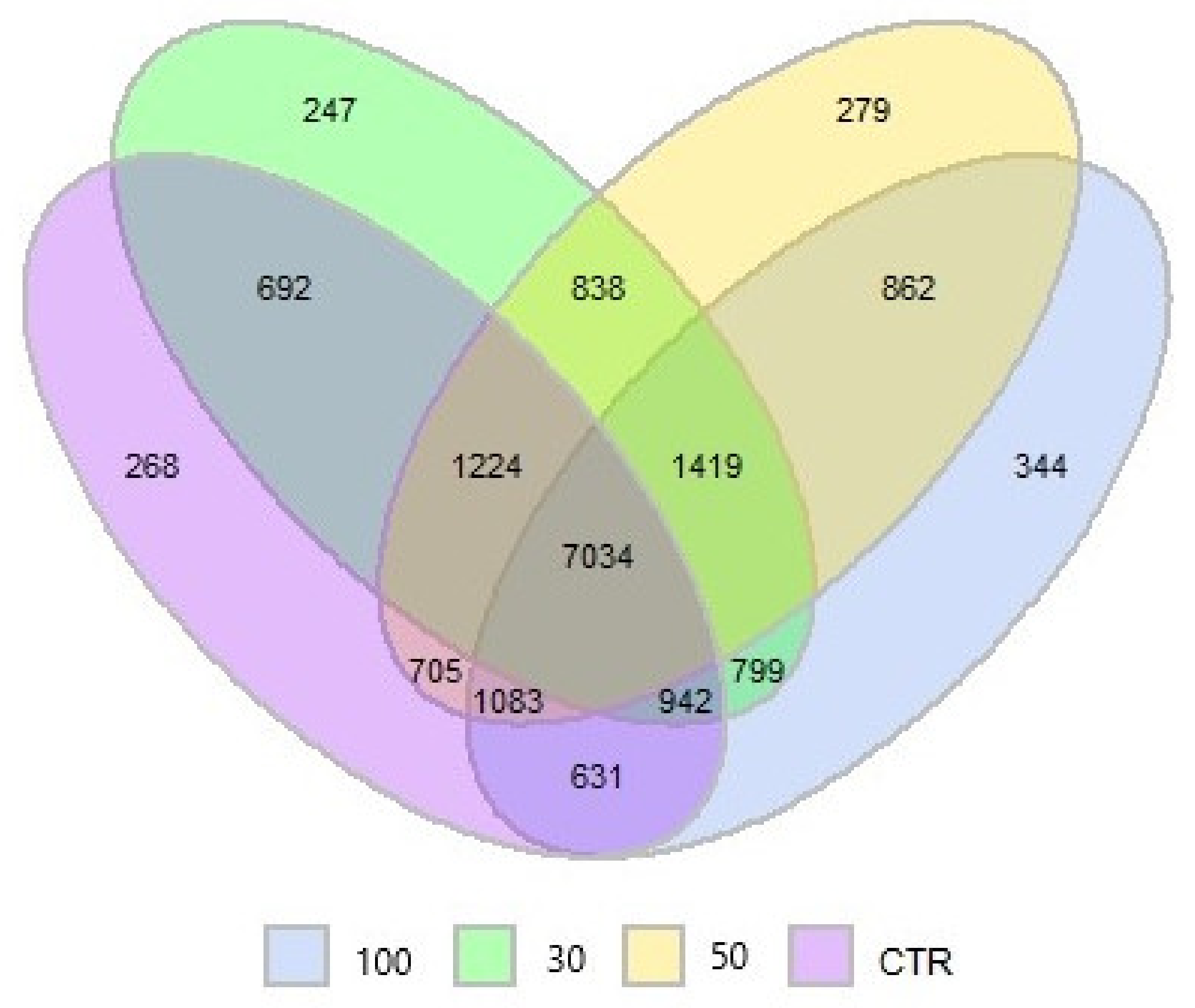
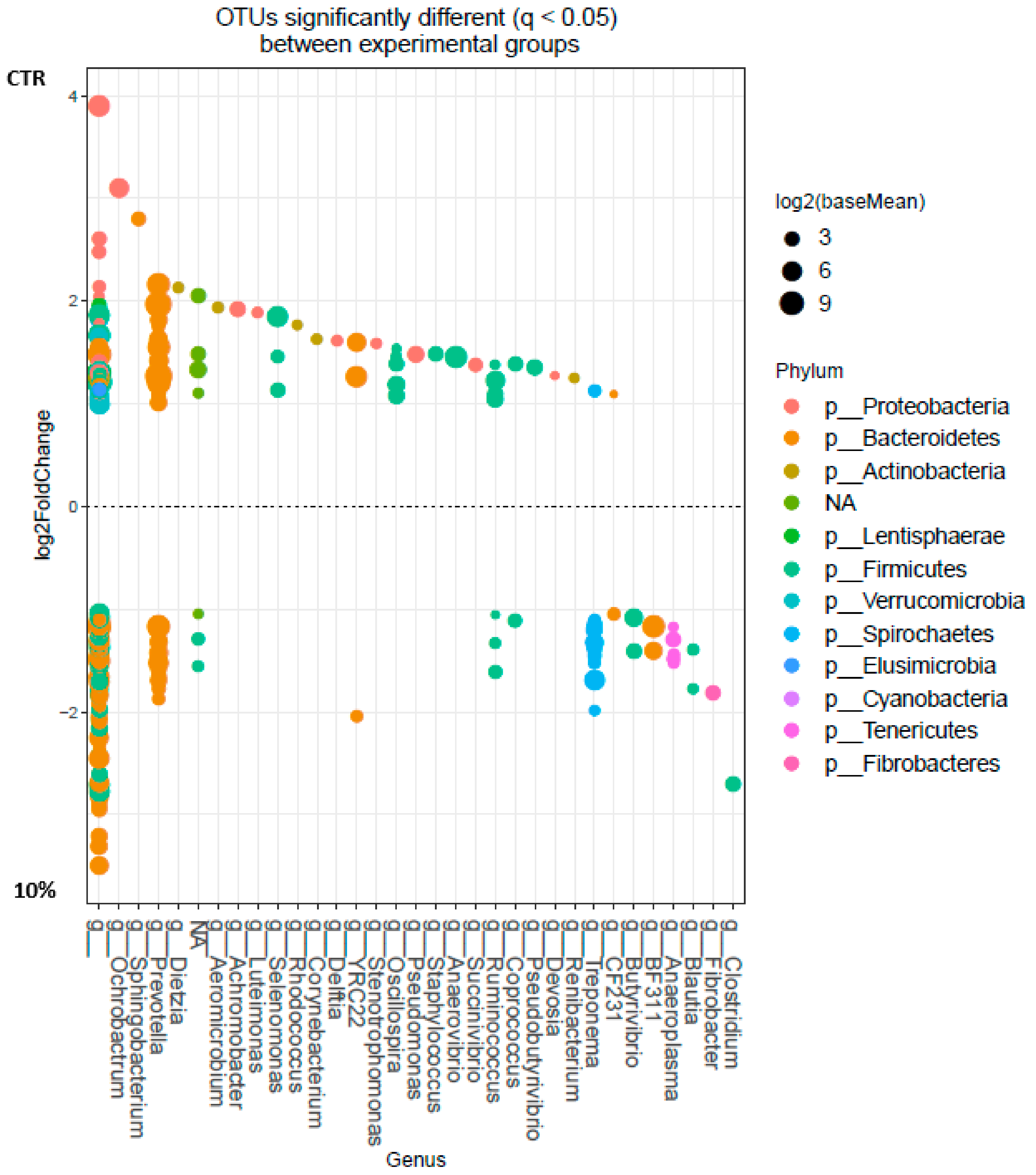
3.1. Ruminal Bacterial Community
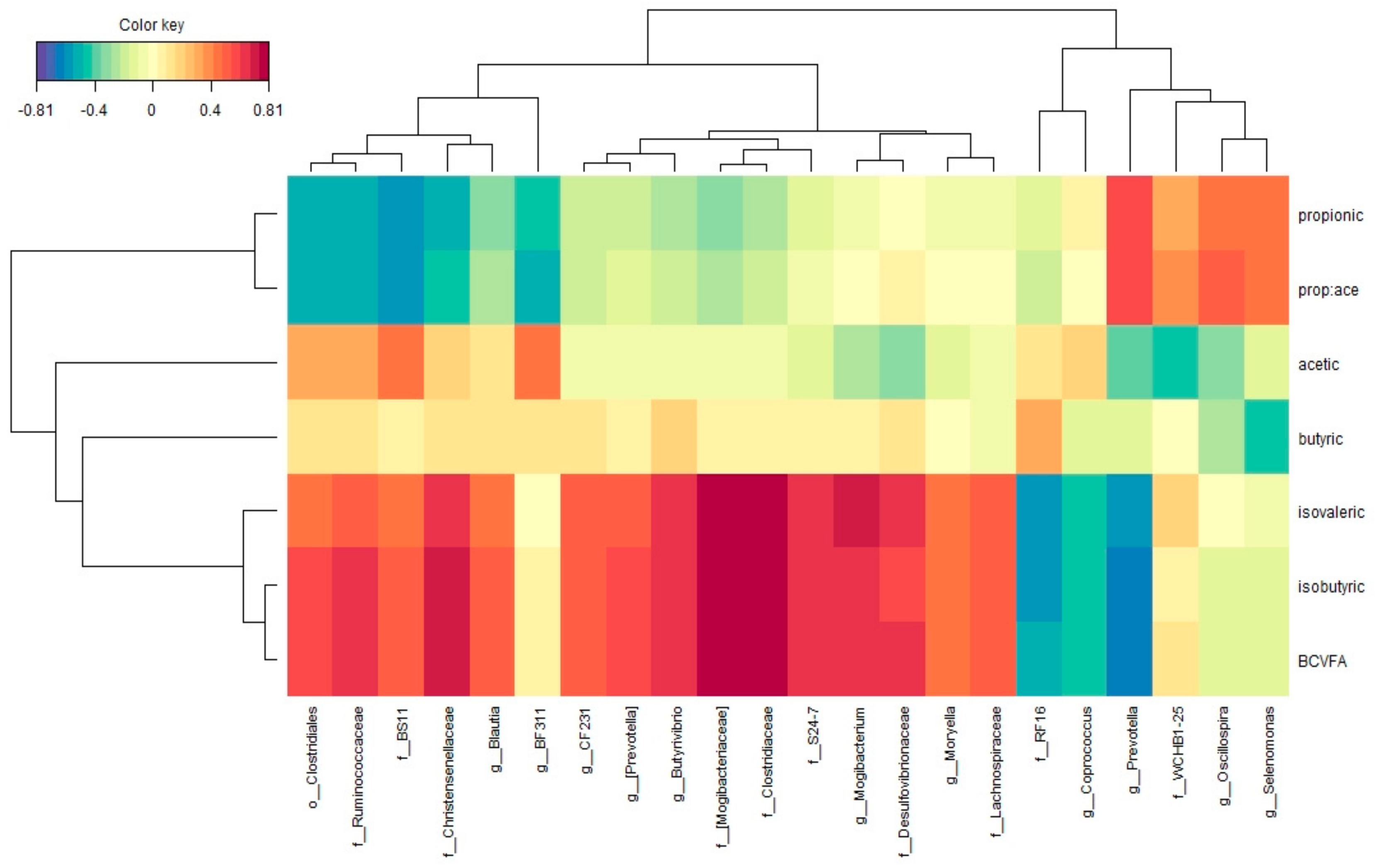
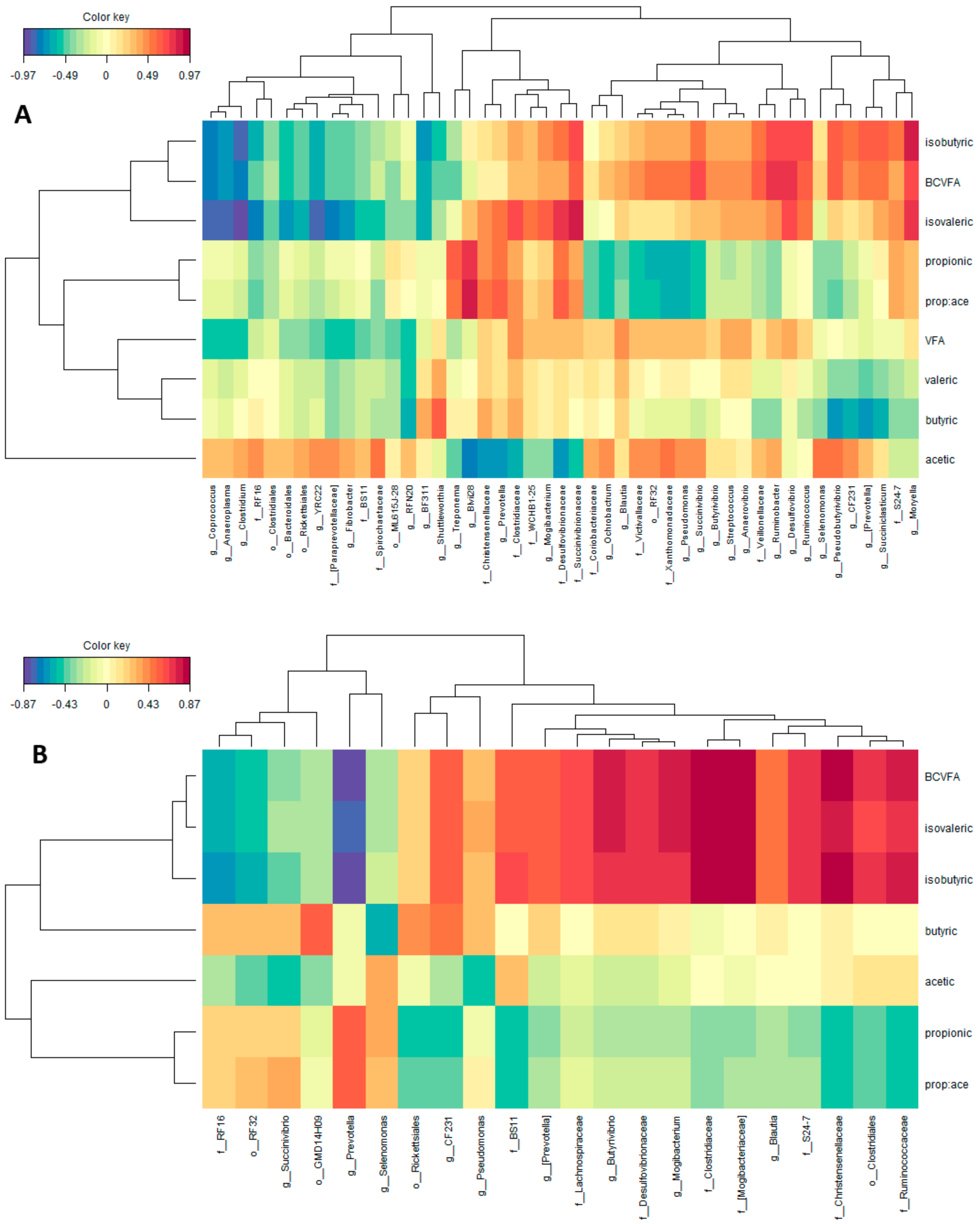
3.2. Correlations between Bacterial Taxa and Ruminal Volatile Fatty Acids
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Girotto, F.; Pivato, A.; Cossu, R.; Nkeng, G.E.; Lavagnolo, M.C. The broad spectrum of possibilities for spent coffee grounds valorisation. J. Mater. Cycles Waste Manag. 2018, 20, 695–701. [Google Scholar] [CrossRef]
- Borrelli, R.C.; Esposito, F.; Napolitano, A.; Ritieni, A.; Fogliano, V. Characterization of a new potential functional ingredient: Coffee silverskin. J. Agric. Food Chem. 2004, 52, 1338–1343. [Google Scholar] [CrossRef] [PubMed]
- Mussatto, S.I.; Machado, E.M.S.; Martins, S.; Teixeira, J.A. Production, composition, and application of coffee and its industrial residues. Food Bioprocess Technol. 2011, 4, 661–672. [Google Scholar] [CrossRef] [Green Version]
- Priolo, A.; Bella, M.; Lanza, M.; Galofaro, V.; Biondi, L.; Barbagallo, D.; Ben Salem, H.; Pennisi, P. Carcass and meat quality of lambs fed fresh sulla (Hedysarum coronarium L.) with or without polyethylene glycol or concentrate. Small Rumin. Res. 2005, 59, 281–288. [Google Scholar] [CrossRef]
- Díaz de Otálora, X.; Ruiz, R.; Goiri, I.; Rey, J.; Atxaerandio, R.; San Martin, D.; Orive, M.; Iñarra, B.; Zufia, J.; Urkiza, J.; et al. Valorisation of spent coffee grounds as functional feed ingredient improves productive performance of Latxa dairy ewes. Anim. Feed Sci. Technol. 2020, 264, 114461. [Google Scholar] [CrossRef]
- Liu, Q.; Wang, C.; Guo, G.; Huo, W.J.; Zhang, Y.L.; Pei, C.X.; Zhang, S.L.; Wang, H. Effects of branched-chain volatile fatty acids supplementation on growth performance, ruminal fermentation, nutrient digestibility, hepatic lipid content and gene expression of dairy calves. Anim. Feed Sci. Technol. 2018, 237, 27–34. [Google Scholar] [CrossRef]
- San Martin, D.; Orive, M.; Iñarra, B.; Garcia-Rodriguez, A.; Goiri, I.; Atxaerandio, R.; Urkiza, J.; Zufia, J. Spent coffee ground as second-generation feedstuff for dairy cattle. Biomass Convers. Biorefin. 2020, 1–11. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Lauber, C.L.; Walters, W.A.; Berg-Lyons, D.; Lozupone, C.A.; Turnbaugh, P.J.; Fierer, N.; Knight, R. Global patterns of 16S rRNA diversity at a depth of millions of sequences per sample. Proc. Natl. Acad. Sci. USA 2011, 108, 4516–4522. [Google Scholar] [CrossRef] [Green Version]
- Magoč, T.; Salzberg, S.L. FLASH: Fast length adjustment of short reads to improve genome assemblies. Bioinformatics 2011, 27, 2957–2963. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [Green Version]
- Edgar, R.C. Search and clustering orders of magnitude faster than BLAST. Bioinformatics 2010, 26, 2460–2461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McDonald, D.; Price, M.N.; Goodrich, J.; Nawrocki, E.P.; DeSantis, T.Z.; Probst, A.; Andersen, G.L.; Knight, R.; Hugenholtz, P. An improved Greengenes taxonomy with explicit ranks for ecological and evolutionary analyses of bacteria and archaea. ISME J. 2012, 6, 610–618. [Google Scholar] [CrossRef] [PubMed]
- Association of Official Analytical Chemists (AOAC). Official Methods of Analysis, 18th ed.; AOAC: Gaitherburg, MD, USA, 2007. [Google Scholar]
- Van Soest, P.J.; Robertson, J.B.; Lewis, B.A. Methods for dietary fiber, neutral detergent fiber, and nonstarch polysaccharides in relation to animal nutrition. J. Dairy Sci. 1991, 74, 3583–3597. [Google Scholar] [CrossRef]
- Robertson, J.B.; Van Soest, P.J. The detergent system of analysis. In The Analysis of Dietary Fibre in Food; James, W.P.T., Theander, O., Eds.; Marcel Dekker: New York, NY, USA, 1981; pp. 123–158. [Google Scholar]
- MAPA. Métodos Oficiales del MAPA; BOE 52, 7167; MAPA: Madrid, Spain, 1995. [Google Scholar]
- International Organization for Standarization (ISO). Coffee and Coffee Products—Determination of the Caffeine Content Using High Performance Liquid Chromatography (HPLC)—Reference Method; ISO 20481:2008; ISO: Geneva, Switzerland, 2008. [Google Scholar]
- Singleton, V.L.; Rossi, J.A. Colorimetry of total phenolics with phosphomolybdic-phosphotungstic acid reagents. Am. J. Enol. Vitic. 1965, 16, 144–158. [Google Scholar]
- Pérez Hernández, L.M.; Chávez-Quiroz, K.; Medina-Juárez, L.A.; Gámez-Meza, N. Phenolic characterization, melanoidins, and antioxidant activity of some commercial coffees from coffea arabica and coffea canephora. J. Chem. Mex. Soc. 2012, 56, 430–435. [Google Scholar]
- SAS. Enterprise’s Guide; SAS: Cary, NC, USA, 2017. [Google Scholar]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [Green Version]
- Le Cao, K.-A.; Rohart, F.; Gonzalez, I.; Dejean, S.; Gautier, B.; Bartolo, F.; Monget, P.; Coquery, J.; Yao, F.Z.; Liquet, B. mixOmics: Omics Data Integration Project. R Package Version. 2016. Available online: https://CRAN.R-project.org/package=mixOmics (accessed on 1 June 2020).
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018; Available online: http://www.R-project.org/ (accessed on 1 June 2020).
- Campbell, T.W.; Bartley, E.E.; Bechtle, R.M.; Dayton, A.D. Coffee Grounds. I. Effects of coffee grounds on ration digestibility and diuresis in cattle, on in vitro rumen fermentation, and on rat growth. J. Dairy Sci. 1976, 59, 1452–1460. [Google Scholar] [CrossRef]
- Bartley, E.E.; Ibbetson, R.W.; Chyba, L.J.; Dayton, A.D. Coffee Grounds. II. Effects of coffee grounds on performance of milking dairy cows and feedlot cattle, and on rumen fermentation and dry matter removal rate. J. Anim. Sci. 1978, 47, 791–799. [Google Scholar] [CrossRef]
- Givens, D.I.; Barber, W.P. In vivo evaluation of spent coffee grounds as a ruminant feed. Agric. Wastes 1986, 18, 69–72. [Google Scholar] [CrossRef]
- Xu, C.C.; Cai, Y.; Zhang, J.G.; Ogawa, M. Fermentation quality and nutritive value of a total mixed ration silage containing coffee grounds at ten or twenty percent of dry matter. J. Anim. Sci. 2007, 85, 1024–1029. [Google Scholar] [CrossRef] [Green Version]
- Jiménez-Zamora, A.; Pastoriza, S.; Rufián-Henares, J.A. Revalorization of coffee by-products. Prebiotic, antimicrobial and antioxidant properties. LWT Food Sci. Technol. 2015, 61, 12–18. [Google Scholar] [CrossRef]
- Vítězová, M.; Jančiková, S.; Dordević, D.; Vítěz, T.; Elbl, J.; Hanišáková, N.; Jampílek, J.; Kushkevych, I. The Possibility of Using Spent Coffee Grounds to Improve Wastewater Treatment Due to Respiration Activity of Microorganisms. Appl. Sci. 2019, 9, 3155. [Google Scholar] [CrossRef] [Green Version]
- Pérez-Burillo, S.; Pastoriza, S.; Fernández-Arteaga, A.; Luzón, G.; Jiménez-Hernández, N.; D’Auria, G.; Francino, M.P.; Rufián-Henares, J.A. Spent Coffee Grounds Extract, Rich in Mannooligosaccharides, Promotes a Healthier Gut Microbial Community in a Dose-Dependent Manner. J. Agric. Food Chem. 2019, 67, 2500–2509. [Google Scholar] [CrossRef] [PubMed]
- Bhandarkar, N.S.; Mouatt, P.; Goncalves, P.; Thomas, T.; Brown, L.; Panchal, S.K. Modulation of gut microbiota by spent coffee grounds attenuates diet-induced metabolic syndrome in rats. FASEB J. 2020, 34, 4783–4797. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, X.; Yang, C.; Guan, L.L.; Wang, J.; Xue, M.; Liu, J.X. Persistence of Cellulolytic Bacteria Fibrobacter and Treponema After Short-Term Corn Stover-Based Dietary Intervention Reveals the Potential to Improve Rumen Fibrolytic Function. Front. Microbiol. 2018, 9, 1363. [Google Scholar] [CrossRef] [PubMed]
- Zhou, M.; Chen, Y.; Guan, L.L. Rumen bacteria. In Rumen Microbiology: From Evolution to Revolution; Puniya, A., Singh, R., Kamra, D., Eds.; Springer: New Delhi, India, 2015; pp. 79–95. [Google Scholar]
- Moreira, A.S.P.; Nunes, F.M.; Domingues, M.R.; Coimbra, M.A. Coffee melanoidins: Structures, mechanisms of formation and potential health impacts. Food Funct. 2012, 3, 903–915. [Google Scholar] [CrossRef]
- Morales, F.J.; Somoza, V.; Fogliano, V. Physiological relevance of dietary melanoidins. Amino Acids 2012, 42, 1097–1109. [Google Scholar] [CrossRef]
- Delgado-Andrade, C.; Fogliano, V. Dietary advanced glycosylation end-products (dAGEs) and melanoidins formed through the Maillard reaction: Physiological consequences of their intake. Annu. Rev. Food Sci. Technol. 2018, 9, 271–291. [Google Scholar] [CrossRef]
- Monlau, F.; Sambusiti, C.; Barakat, A.; Quéméneur, M.; Trably, E.; Steyer, J.P.; Carrère, H. Do furanic and phenolic compounds of lignocellulosic and algae biomass hydrolysate inhibit anaerobic mixed cultures? A comprehensive review. Biotechnol. Adv. 2014, 32, 934–951. [Google Scholar] [CrossRef]
- Ramón-Gonçalves, M.; Alcaraz, L.; Pérez-Ferreras, S.; León-González, M.E.; Rosales-Conrado, N.; López, F.A. Extraction of polyphenols and synthesis of new activated carbon from spent coffee grounds. Sci. Rep. 2019, 9, 17706. [Google Scholar] [CrossRef] [Green Version]
- Akin, D.E. Forage Cell Wall Degradation and ρ-Coumaric, Ferulic, and Sinapic Acids1. Agron. J. 1982, 74, 424–428. [Google Scholar] [CrossRef]
- Filosa, S.; Di Meo, F.; Crispi, S. Polyphenols-gut microbiota interplay and brain neuromodulation. Neural Regen. Res. 2018, 13, 2055–2059. [Google Scholar] [PubMed]
- Chesson, A.; Provan, G.J.; Russell, W.R.; Scobbie, L.; Richardson, A.J.; Stewart, C. Hydroxycinnamic acids in the digestive tract of livestock and humans. J. Sci. Food Agric. 1999, 79, 373–378. [Google Scholar] [CrossRef]
- Patra, A.K.; Park, T.; Braun, H.-S.; Geiger, S.; Pieper, R.; Yu, Z.; Aschenbach, J.R. Dietary Bioactive Lipid Compounds Rich in Menthol Alter Interactions Among Members of Ruminal Microbiota in Sheep. Front. Microbiol. 2019, 10, 2038. [Google Scholar] [CrossRef]
- Russell, J.B.; Wallace, R.J. Energy Yielding and consuming reactions. In The Rumen Microbial Ecosystem; Hobson, P.N., Steward, C.S., Eds.; Blackie Academic & Professional: London, UK, 1997; pp. 246–282. [Google Scholar]
- Tapio, I.; Fischer, D.; Blasco, L.; Tapio, M.; Wallace, R.J.; Bayat, A.R.; Ventto, L.; Kahala, M.; Negussie, E.; Shingfield, K.J.; et al. Taxon abundance, diversity, co-occurrence and network analysis of the ruminal microbiota in response to dietary changes in dairy cows. PLoS ONE 2017, 12, e0180260. [Google Scholar] [CrossRef] [Green Version]
- Wolff, S.M.; Ellison, M.J.; Hao, Y.; Cockrum, R.R.; Austin, K.J.; Baraboo, M.; Burch, K.; Jin Lee, H.; Maurer, T.; Patil, R.; et al. Diet shifts provoke complex and variable changes in the metabolic networks of the ruminal microbiome. Microbiome 2017, 5, 60. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Diversity Measurements | Treatment a | SEM | p-Value | ||||
|---|---|---|---|---|---|---|---|
| Control | 3 | 5 | 10 | Linear | Quadratic | ||
| Observed OTU | 4347 | 4501 | 4659 | 4685 | 445.3 | 0.116 | 0.491 |
| Chao1 | 7025 | 7330 | 7607 | 7650 | 698.8 | 0.069 | 0.393 |
| Coverage (%) | 97.8 | 97.5 | 97.4 | 97.4 | 0.35 | 0.034 | 0.278 |
| Phylogenetic diversity | 124 | 128 | 130 | 135 | 8.3 | 0.007 | 0.897 |
| Shannon | 7.82 | 8.33 | 8.33 | 8.72 | 0.4547 | 0.001 | 0.412 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Goiri, I.; Díaz de Otálora, X.; Ruiz, R.; Rey, J.; Atxaerandio, R.; Lavín, J.L.; San Martin, D.; Orive, M.; Iñarra, B.; Zufia, J.; et al. Spent Coffee Grounds Alter Bacterial Communities in Latxa Dairy Ewes. Microorganisms 2020, 8, 1961. https://doi.org/10.3390/microorganisms8121961
Goiri I, Díaz de Otálora X, Ruiz R, Rey J, Atxaerandio R, Lavín JL, San Martin D, Orive M, Iñarra B, Zufia J, et al. Spent Coffee Grounds Alter Bacterial Communities in Latxa Dairy Ewes. Microorganisms. 2020; 8(12):1961. https://doi.org/10.3390/microorganisms8121961
Chicago/Turabian StyleGoiri, Idoia, Xabier Díaz de Otálora, Roberto Ruiz, Jagoba Rey, Raquel Atxaerandio, Jose Luis Lavín, David San Martin, Mikel Orive, Bruno Iñarra, Jaime Zufia, and et al. 2020. "Spent Coffee Grounds Alter Bacterial Communities in Latxa Dairy Ewes" Microorganisms 8, no. 12: 1961. https://doi.org/10.3390/microorganisms8121961
APA StyleGoiri, I., Díaz de Otálora, X., Ruiz, R., Rey, J., Atxaerandio, R., Lavín, J. L., San Martin, D., Orive, M., Iñarra, B., Zufia, J., Urkiza, J., & García-Rodríguez, A. (2020). Spent Coffee Grounds Alter Bacterial Communities in Latxa Dairy Ewes. Microorganisms, 8(12), 1961. https://doi.org/10.3390/microorganisms8121961