Performance of a Four-Antigen Staphylococcus aureus Vaccine in Preclinical Models of Invasive S. aureus Disease

Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains
2.2. Animal Studies
2.2.1. Surgical Site Infection Mouse Model
2.2.2. Murine Bacteremia Model
2.2.3. Murine Pyelonephritis Model
2.2.4. Rat Endocarditis Model
Standard Model
Refined Model
2.3. Assessment of Immune Responses to Vaccination
2.3.1. Opsonophagocytic Activity Assay
2.3.2. Competitive Luminex Immunoassay
3. Results
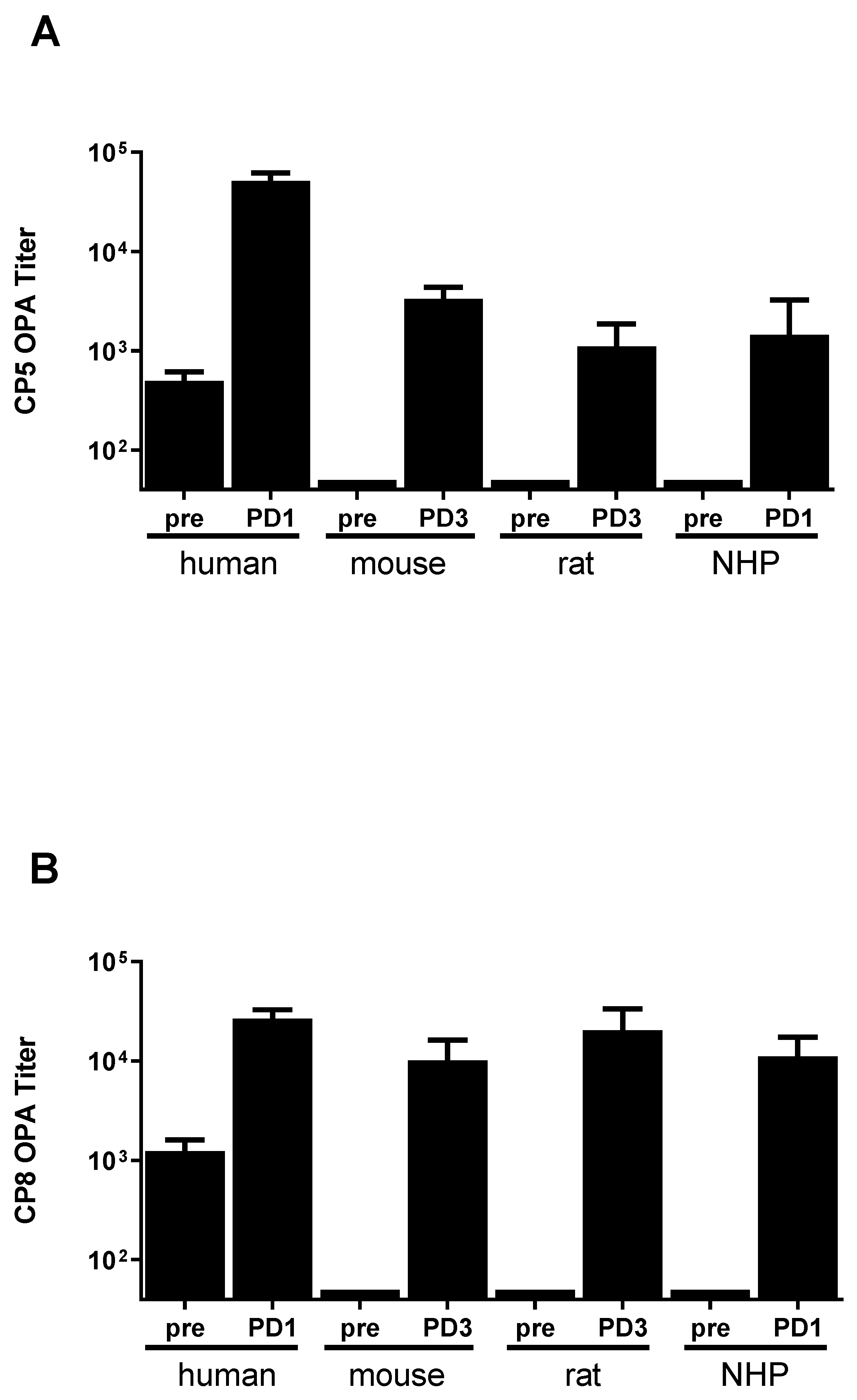
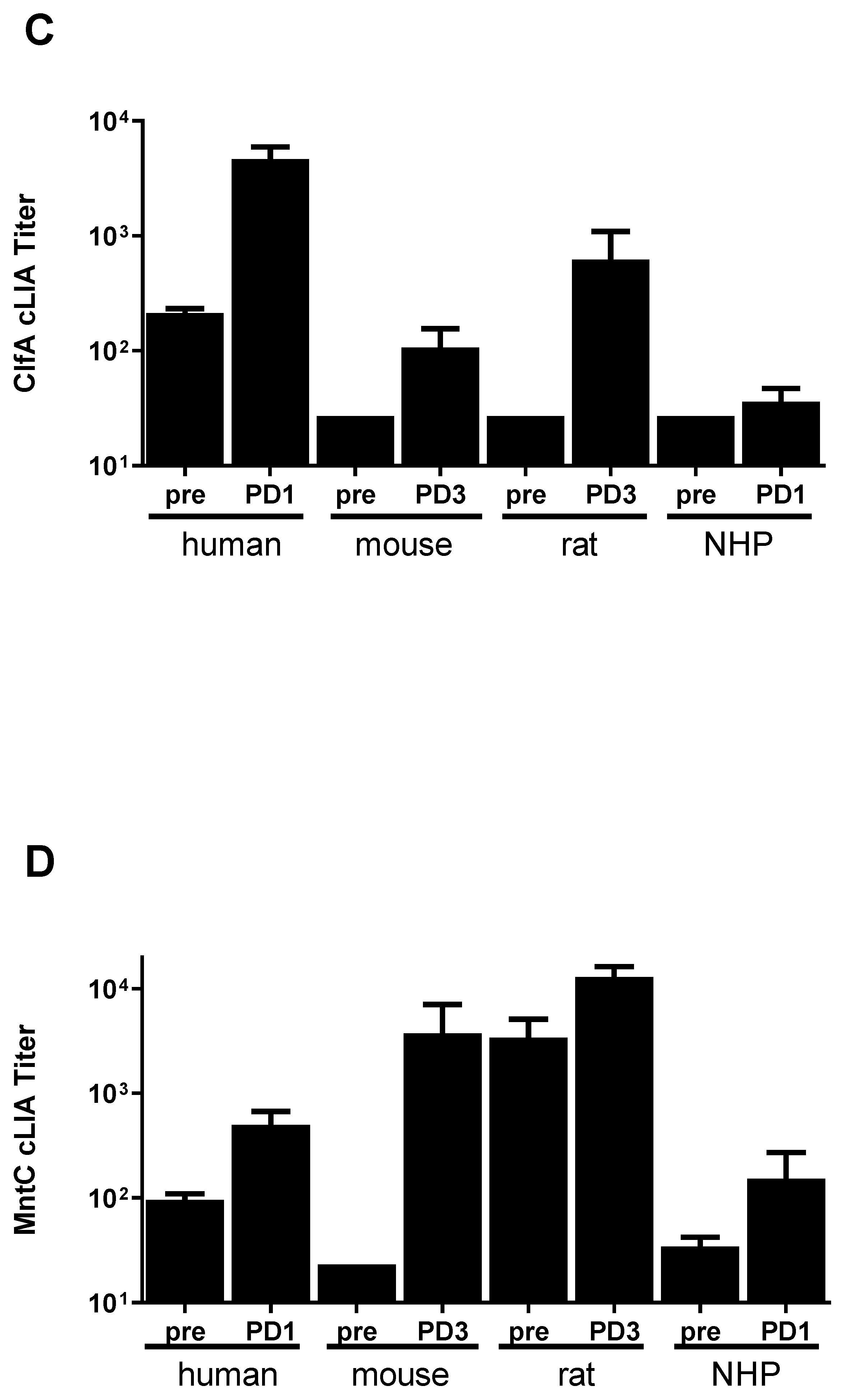
3.1. SA4Ag Is Immunogenic in Mice, Rats and Non-Human Primates
3.2. Immunization with SA4Ag Reduces Bacterial Burden in a Murine Model of Surgical Site Infection
3.3. Immunization with SA4Ag Protects against MRSA Challenge in a Murine Bacteremia Model
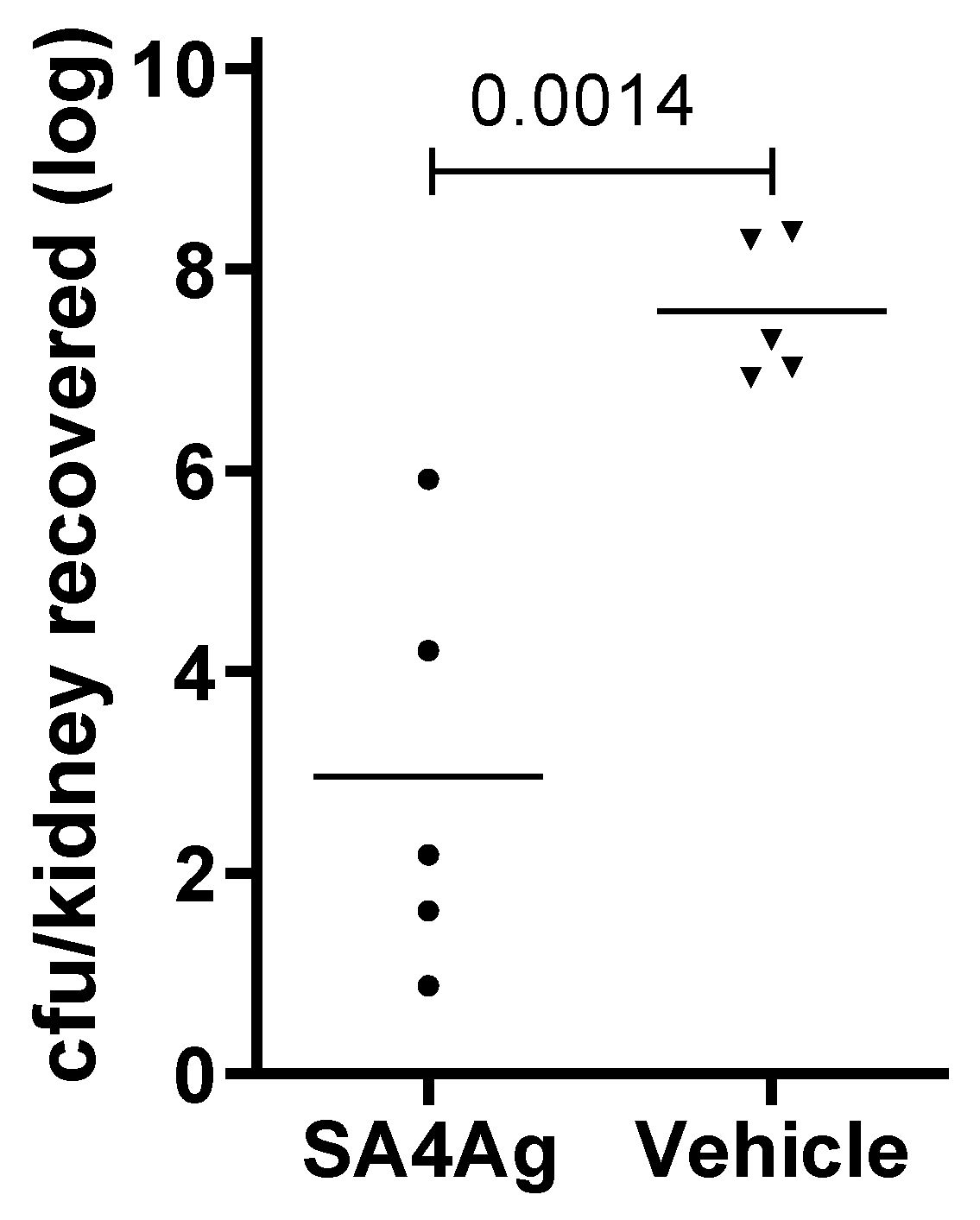
3.4. Immunization with SA4Ag Reduces Bacterial Burden in a Murine Pyelonephritis Model
3.5. Immunization with SA4Ag Protects against Both CP5- and CP8-Expressing S. aureus in a Rat Endocarditis Model
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kluytmans, J.; van Belkum, A.; Verbrughn, H. Nasal carriage of Staphylococcus aureus: Epidemiology, underlying mechanisms, and associated risks. Clin. Microbiol. Rev. 1997, 10, 505–520. [Google Scholar] [CrossRef]
- Lowy, F.D. Staphylococcus aureus infections. N. Engl. J. Med. 1998, 339, 520–532. [Google Scholar] [CrossRef] [PubMed]
- Noskin, G.A.; Rubin, R.J.; Schentag, J.J.; Kluytmans, J.; Hedblom, E.C.; Jacobson, C.; Smulders, M.; Gemmen, E.; Bharmal, M. National trends in Staphylococcus aureus infection rates: Impact on economic burden and mortality over a 6-year period (1998–2003). Clin. Infect. Dis. 2007, 45, 1132–1140. [Google Scholar] [CrossRef] [PubMed]
- Carrel, M.; Perencevich, E.N.; David, M.Z. USA300 Methicillin-Resistant Staphylococcus aureus, United States, 2000–2013. Emerg. Infect. Dis. 2015, 21, 1973–1980. [Google Scholar] [CrossRef] [PubMed]
- Malachowa, N.; Whitney, A.R.; Kobayashi, S.D.; Sturdevant, D.E.; Kennedy, A.D.; Braughton, K.R.; Shabb, D.W.; Diep, B.A.; Chambers, H.F.; Otto, M.; et al. Global changes in Staphylococcus aureus gene expression in human blood. PLoS ONE 2011, 6, e18617. [Google Scholar] [CrossRef] [Green Version]
- Bode, L.; Kluytmans, J.; Wertheim, H.; Bogaers, D.; Vandenbroucke-Grauls, C.; Roosendaal, R.; Troelstra, A.; Box, A.; Voss, A.; Van Der Tweel, I.; et al. Preventing surgical-site infections in nasal carriers of Staphylococcus aureus. N. Engl. J. Med. 2010, 362, 9–17. [Google Scholar] [CrossRef] [Green Version]
- Kalmeijer, M.D.; Coertjens, H.; Van Nieuwland-Bollen, P.M.; Bogaers-Hofman, D.; De Baere, G.A.J.; Stuurman, A.; Van Belkum, A.; Kluytmans, J.A.J.W. Surgical site infections in orthopedic surgery: The effect of mupirocin nasal ointment in a double-blind, randomized, placebo-controlled study. Clin. Infect. Dis. 2002, 35, 353–358. [Google Scholar] [CrossRef]
- Simor, A.E.; Phillips, E.; McGeer, A.; Konvalinka, A.; Loeb, M.; Devlin, H.R.; Kiss, A. Randomized controlled trial of chlorhexidine gluconate for washing, intranasal mupirocin, and rifampin and doxycycline versus no treatment for the eradication of methicillin-resistant Staphylococcus aureus colonization. Clin. Infect. Dis. 2007, 44, 178–185. [Google Scholar] [CrossRef]
- Simor, A.E.; Stuart, T.L.; Louie, L.; Watt, C.; Ofner-Agostini, M.; Gravel, D.; Mulvey, M.; Loeb, M.; McGeer, A.; Bryce, E.; et al. Mupirocin-resistant, methicillin-resistant Staphylococcus aureus strains in Canadian hospitals. Antimicrob. Agents Chemother. 2007, 51, 3880–3886. [Google Scholar] [CrossRef] [Green Version]
- Hawkins, J.; Kodali, S.; Matsuka, Y.V.; McNeil, L.K.; Mininni, T.; Scully, I.L.; Vernachio, J.H.; Severina, E.; Girgenti, D.; Jansen, K.U.; et al. A recombinant clumping factor A-containing vaccine induces functional antibodies to Staphylococcus aureus that are not observed after natural exposure. Clin. Vaccine Immunol. 2012, 19, 1641–1650. [Google Scholar] [CrossRef] [Green Version]
- Anderson, A.S.; Scully, I.L.; Timofeyeva, Y.; Murphy, E.; McNeil, L.K.; Mininni, T.; Nuñez, L.; Carriere, M.; Singer, C.; Dilts, D.A.; et al. Staphylococcus aureus manganese transport protein C is a highly conserved cell surface protein that elicits protective immunity against S. aureus and Staphylococcus epidermidis. J. Infect. Dis. 2012, 205, 1688–1696. [Google Scholar] [CrossRef] [PubMed]
- Brady, R.A.; Leid, J.G.; Camper, A.K.; Costerton, J.W.; Shirtliff, M.E. Identification of Staphylococcus aureus proteins recognized by the antibody-mediated immune response to a biofilm infection. Infect. Immun. 2006, 74, 3415–3426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Handke, L.D.; Hawkins, J.C.; Miller, A.A.; Jansen, K.U.; Anderson, A.S. Regulation of Staphylococcus aureus MntC Expression and Its Role in Response to Oxidative Stress. PLoS ONE 2013, 8, e77874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nanra, J.S.; Buitrago, S.M.; Crawford, S.; Ng, J.; Fink, P.S.; Hawkins, J.; Scully, I.L.; McNeil, L.K.; Aste-Amézaga, J.M.; Cooper, D.; et al. Capsular polysaccharides are an important immune evasion mechanism for Staphylococcus aureus. Hum. Vaccines Immunother. 2012, 9, 480–487. [Google Scholar] [CrossRef] [Green Version]
- Thakker, M.; Park, J.S.; Carey, V.; Lee, J.C. Staphylococcus aureus serotype 5 capsular polysaccharide is antiphagocytic and enhances bacterial virulence in a murine bacteremia model. Infect. Immun. 1998, 66, 5183–5189. [Google Scholar] [CrossRef] [Green Version]
- Fattom, A.; Schneerson, R.; Szu, S.C.; Vann, W.F.; Shiloach, J.; Karakawa, W.W.; Robbins, J.B. Synthesis and immunologic properties in mice of vaccines composed of Staphylococcus aureus type 5 and type 8 capsular polysaccharides conjugated to Pseudomonas aeruginosa exotoxin A. Infect Immun. 1990, 58, 2367–2374. [Google Scholar] [CrossRef] [Green Version]
- Kuklin, N.A.; Clark, D.J.; Secore, S.; Cook, J.; Cope, L.D.; McNeely, T.; Noble, L.; Brown, M.J.; Zorman, J.K.; Wang, X.M.; et al. A novel Staphylococcus aureus vaccine: Iron surface determinant B induces rapid antibody responses in rhesus macaques and specific increased survival in a murine S. aureus sepsis model. Infect. Immun. 2006, 74, 2215–2223. [Google Scholar] [CrossRef] [Green Version]
- Joshi, A.; Pancari, G.; Cope, L.; Bowman, E.P.; Cua, D.; Proctor, R.A.; McNeely, T. Immunization with Staphylococcus aureus iron regulated surface determinant B (IsdB) confers protection via Th17/IL17 pathway in a murine sepsis model. Hum. Vaccines Immunother. 2012, 8, 336–346. [Google Scholar] [CrossRef] [Green Version]
- Pozzi, C.; Olaniyi, R.; Liljeroos, L.; Galgani, I.; Rappuoli, R.; Bagnoli, F. Vaccines for Staphylococcus aureus and Target Populations. Curr. Top. Microbiol. Immunol. 2017, 409, 491–528. [Google Scholar] [CrossRef]
- Jansen, K.U.; Girgenti, D.Q.; Scully, I.L.; Anderson, A.S. Vaccine review: “Staphyloccocus aureus vaccines: Problems and prospects”. Vaccine 2013, 31, 2723–2730. [Google Scholar] [CrossRef]
- Scully, I.L.; Timofeyeva, Y.; Keeney, D.; Matsuka, Y.V.; Severina, E.; McNeil, L.K.; Nanra, J.; Hu, G.; Liberator, P.A.; Jansen, K.U.; et al. Demonstration of the preclinical correlate of protection for Staphylococcus aureus clumping factor A in a murine model of infection. Vaccine 2015, 33, 5452–5457. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, L.; Lee, J.C. Murine Models of Bacteremia and Surgical Wound Infection for the Evaluation of Staphylococcus aureus Vaccine Candidates. Methods Mol. Biol. 2016, 1403, 409–418. [Google Scholar] [CrossRef] [PubMed]
- Schennings, T.; Heimdahl, A.; Coster, K.; Flock, J.I. Immunization with fibronectin binding protein from Staphylococcus aureus protects against experimental endocarditis in rats. Microb. Pathog. 1993, 15, 227–236. [Google Scholar] [CrossRef] [PubMed]
- Chambers, H.F.; Korzeniowski, O.M.; Sande, M.A. Staphylococcus aureus endocarditis: Clinical manifestations in addicts and nonaddicts. Medicine 1983, 62, 170–177. [Google Scholar] [CrossRef] [PubMed]
- Gribenko, A.V.; Mosyak, L.; Ghosh, S.; Parris, K.; Svenson, K.; Moran, J.; Chu, L.; Li, S.; Liu, T.; Woods, V.L.; et al. Three-dimensional structure and biophysical characterization of Staphylococcus aureus cell surface antigen-manganese transporter MntC. J. Mol. Biol. 2013, 425, 3429–3445. [Google Scholar] [CrossRef]
- Gribenko, A.V.; Parris, K.; Mosyak, L.; Lidia, M.; Handke, L.; Hawkins, J.C.; Severina, E.; Matsuka, Y.V.; Anderson, A.S. High Resolution Mapping of Bactericidal Monoclonal Antibody Binding Epitopes on Staphylococcus aureus Antigen MntC. PLoS Pathog. 2016, 12, e1005908. [Google Scholar] [CrossRef] [Green Version]
- Frenck, R.W., Jr.; Creech, C.B.; Sheldon, E.; Seiden, D.J.; Kankam, M.K.; Baber, J.; Zito, E.; Hubler, R.; Eiden, J.J.; Severs, J.M.; et al. Safety, tolerability, and immunogenicity of a 4-antigen Staphylococcus aureus vaccine (SA4Ag): Results from a first-in-human randomised, placebo-controlled phase 1/2 study. Vaccine 2017, 35, 375–384. [Google Scholar] [CrossRef]
- Li, X.; Wang, X.; Thompson, C.D.; Park, S.; Park, W.B.; Lee, J.C. Preclinical efficacy of clumping factor A in prevention of Staphylococcus aureus infection. mBio 2016, 7, e02232-15. [Google Scholar] [CrossRef] [Green Version]
- Zhu, D.; Tuo, W. QS-21: A Potent Vaccine Adjuvant. Nat. Prod. Chem. Res. 2016, 3, e113. [Google Scholar] [CrossRef] [Green Version]
- Bigaeva, E.; Doorn, E.; Liu, H.; Hak, E. Meta-Analysis on Randomized Controlled Trials of Vaccines with QS-21 or ISCOMATRIX Adjuvant: Safety and Tolerability. PLoS ONE 2016, 11, e0154757. [Google Scholar] [CrossRef] [Green Version]
- Nissen, M.D.; Marshall, H.S.; Richmond, P.; Shakib, S.; Jiang, Q.; Cooper, D.; Rill, D.; Baber, J.; Eiden, J.J.; Gruber, W.; et al. A randomized phase I study of the safety and immunogenicity of three ascending dose levels of a 3-antigen Staphylococcus aureus vaccine (SA3Ag) in healthy adults. Vaccine 2015, 33, 1846–1854. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fattom, A.; Matalon, A.; Buerkert, J.; Taylor, K.; Damaso, S.; Boutriau, D. Efficacy profile of a bivalent Staphylococcus aureus glycoconjugated vaccine in adults on hemodialysis: Phase III randomized study. Hum. Vaccines Immunother. 2015, 11, 632–641. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fowler, V.G.; Allen, K.B.; Moreira, E.D.; Moustafa, M.; Isgro, F.; Boucher, H.W.; Corey, G.R.; Carmeli, Y.; Betts, R.; Hartzel, J.S.; et al. Effect of an investigational vaccine for preventing Staphylococcus aureus infections after cardiothoracic surgery: A randomized trial. JAMA 2013, 309, 1368–1378. [Google Scholar] [CrossRef] [PubMed]
- Haley, K.P.; Skaar, E.P. A battle for iron: Host sequestration and Staphylococcus aureus acquisition. Microbes Infect. 2012, 14, 217–227. [Google Scholar] [CrossRef] [Green Version]
- Stranger-Jones, Y.K.; Bae, T.; Schneewind, O. Vaccine assembly from surface proteins of Staphylococcus aureus. Proc. Natl. Acad. Sci. USA 2006, 103, 16942–16947. [Google Scholar] [CrossRef] [Green Version]
- Bagnoli, F.; Fontana, M.R.; Soldaini, E.; Mishra, R.P.N.; Fiaschi, L.; Cartocci, E.; Nardi-Dei, V.; Ruggiero, P.; Nosari, S.; De Falco, M.G.; et al. Vaccine composition formulated with a novel TLR7-dependent adjuvant induces high and broad protection against Staphylococcus aureus. Proc. Natl. Acad. Sci. USA 2015, 112, 3680–3685. [Google Scholar] [CrossRef] [Green Version]
- Torre, A.; Bacconi, M.; Sammicheli, C.; Galletti, B.; Laera, D.; Fontana, M.R.; Grandi, G.; De Gregorio, E.; Bagnoli, F.; Nuti, S.; et al. Four-Component Staphylococcus aureus Vaccine 4C-Staph Enhances Fcgamma Receptor Expression in Neutrophils and Monocytes and Mitigates S. aureus Infection in Neutropenic Mice. Infect. Immun. 2015, 83, 3157–3163. [Google Scholar] [CrossRef] [Green Version]
- Levy, J.; Licini, L.; Haelterman, E.; Moris, P.; Lestrate, P.; Damaso, S.; Van Belle, P.; Boutriau, D. Safety and immunogenicity of an investigational 4-component Staphylococcus aureus vaccine with or without AS03 adjuvant: Results of a randomized phase I trial. Hum. Vaccines Immunother. 2015, 11, 620–631. [Google Scholar] [CrossRef] [Green Version]
- ClinicalTrials.gov. Available online: https://clinicaltrials.gov/ct2/show/NCT02388165?term=NCT02388165&draw=2&rank=1 (accessed on 25 April 2020).
- Pfizer Inc. Available online: https://investors.pfizer.com/investor-news/press-release-details/2018/Independent-Data-Monitoring-Committee-Recommends-Discontinuation-of-the-Phase-2b-STRIVE-Clinical-Trial-of-Staphylococcus-aureus-Vaccine-Following-Planned-Interim-Analysis/default.aspx (accessed on 25 April 2020).




{kind=link}
{kind=link}
{kind=link}
| Strain Name | Capsule Type | Source | MRSA/MSSA | ST/CC | Comments |
|---|---|---|---|---|---|
| CDC3 (USA300) | 5 | ID | MRSA | ST8/CC8 | PVL+ |
| PFESA0241 (USA300) | 5 | ID | MRSA | ST8/CC8 | Mec IV, PVL+ TSST-neg |
| PFESA0158 | 5 | ID | MSSA | ST28/CC25 | |
| PFESA0186 | 8 | Carriage | MSSA | ST57/CC30 | |
| Reynolds | 5 | ID | MSSA | ST25/CC25 |
| Trial | Vaccine | Number of Animals | Mean log CFU/mL (95% CI) | p Value vs. Control |
|---|---|---|---|---|
| 1 | SA4Ag | 20 | 4.65 (3.90–5.40) | 0.0393 |
| Vehicle | 20 | 5.72 (4.99–6.45) | ||
| 2 | SA4Ag | 15 | 6.42 (5.07–7.77) | 0.0330 |
| Vehicle | 13 | 7.89 (6.95–8.83) | ||
| 3 | SA4Ag | 17 | 5.95 (4.60–7.31) | 0.2899 |
| Vehicle | 16 | 6.45 (5.11–7.81) | ||
| Meta-analysis | SA4Ag | 48 | 6.29 (5.57–7.01) | 0.0126 |
| Vehicle | 43 | 7.36 (6.75–7.97) |
| Challenge | Trial | Vaccine | Number of Animals | Mean log CFU/mL (95% CI) | p Value vs. Control |
|---|---|---|---|---|---|
| S. aureus CDC3 (USA300) | 1 | SA4Ag | 10 | 4.65 (4.33–4.97) | 0.0056 |
| Vehicle | 10 | 5.35 (4.96–5.73) | |||
| 2 | SA4Ag | 10 | 4.49 (4.07–4.90) | 0.0147 | |
| Vehicle | 10 | 5.24 (4.76–5.72) | |||
| 3 | SA4Ag | 10 | 3.63 (3.25–4.01) | 0.0056 | |
| Vehicle | 10 | 4.47 (4.00–4.94) | |||
| Meta-analysis | SA4Ag | 30 | 4.25 (4.00–4.51) | <0.0001 | |
| Vehicle | 30 | 5.02 (4.75–5.29) | |||
| S. aureus PFESA0241 (USA300) | 1 | SA4Ag | 10 | 4.17 (3.83–4.51) | 0.0491 |
| Vehicle | 10 | 4.68 (4.25–5.12) | |||
| 2 | SA4Ag | 10 | 4.26 (3.93–4.59) | 0.0157 | |
| Vehicle | 10 | 4.82 (4.48–5.15) | |||
| 3 | SA4Ag | 10 | 4.38 (3.98–4.78) | 0.0455 | |
| Vehicle | 10 | 4.88 (4.54–5.22) | |||
| Meta-analysis | SA4Ag | 30 | 4.27 (4.09–4.45) | 0.0002 | |
| vehicle | 30 | 4.79 (4.60–4.98) |
| Trial | Vaccine | Number of Animals | Mean Log CFU/mL (95% CI) | p Value vs. Control |
|---|---|---|---|---|
| 1 | SA4Ag | 16 | 6.52 (5.20–7.84) | 0.0319 |
| Vehicle | 14 | 7.88 (7.19–8.58) | ||
| 2 | SA4Ag | 15 | 6.42 (5.07–7.77) | 0.0330 |
| Vehicle | 13 | 7.89 (6.95–8.83) | ||
| 3 | SA4Ag | 17 | 5.95 (4.60–7.31) | 0.2899 |
| Vehicle | 16 | 6.45 (5.11–7.81) | ||
| Meta-analysis | SA4Ag | 48 | 6.29 (5.57–7.01) | 0.0126 |
| Vehicle | 43 | 7.36 (6.75–7.97) |
| Experiment | Vaccine | Number of Infected Animals | Number of Uninfected Animals | p Value vs. Control |
|---|---|---|---|---|
| 1 | SA4Ag | 0 | 11 | 0.0983 |
| Vehicle | 4 | 9 | ||
| 2 | SA4Ag | 1 | 11 | 0.0730 |
| Vehicle | 6 | 7 | ||
| Meta-analysis | SA4Ag | 1 | 22 | <0.0001 |
| Vehicle | 10 | 16 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Scully, I.L.; Timofeyeva, Y.; Illenberger, A.; Lu, P.; Liberator, P.A.; Jansen, K.U.; Anderson, A.S. Performance of a Four-Antigen Staphylococcus aureus Vaccine in Preclinical Models of Invasive S. aureus Disease. Microorganisms 2021, 9, 177. https://doi.org/10.3390/microorganisms9010177
Scully IL, Timofeyeva Y, Illenberger A, Lu P, Liberator PA, Jansen KU, Anderson AS. Performance of a Four-Antigen Staphylococcus aureus Vaccine in Preclinical Models of Invasive S. aureus Disease. Microorganisms. 2021; 9(1):177. https://doi.org/10.3390/microorganisms9010177
Chicago/Turabian StyleScully, Ingrid L., Yekaterina Timofeyeva, Arthur Illenberger, Peimin Lu, Paul A. Liberator, Kathrin U. Jansen, and Annaliesa S. Anderson. 2021. "Performance of a Four-Antigen Staphylococcus aureus Vaccine in Preclinical Models of Invasive S. aureus Disease" Microorganisms 9, no. 1: 177. https://doi.org/10.3390/microorganisms9010177
APA StyleScully, I. L., Timofeyeva, Y., Illenberger, A., Lu, P., Liberator, P. A., Jansen, K. U., & Anderson, A. S. (2021). Performance of a Four-Antigen Staphylococcus aureus Vaccine in Preclinical Models of Invasive S. aureus Disease. Microorganisms, 9(1), 177. https://doi.org/10.3390/microorganisms9010177