Adjunct Culture of Non-Starter Lactic Acid Bacteria for the Production of Provola Dei Nebrodi PDO Cheese: In Vitro Screening and Pilot-Scale Cheese-Making


 ,
,  , ,
, ,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cheese Samples and NSLAB Isolation
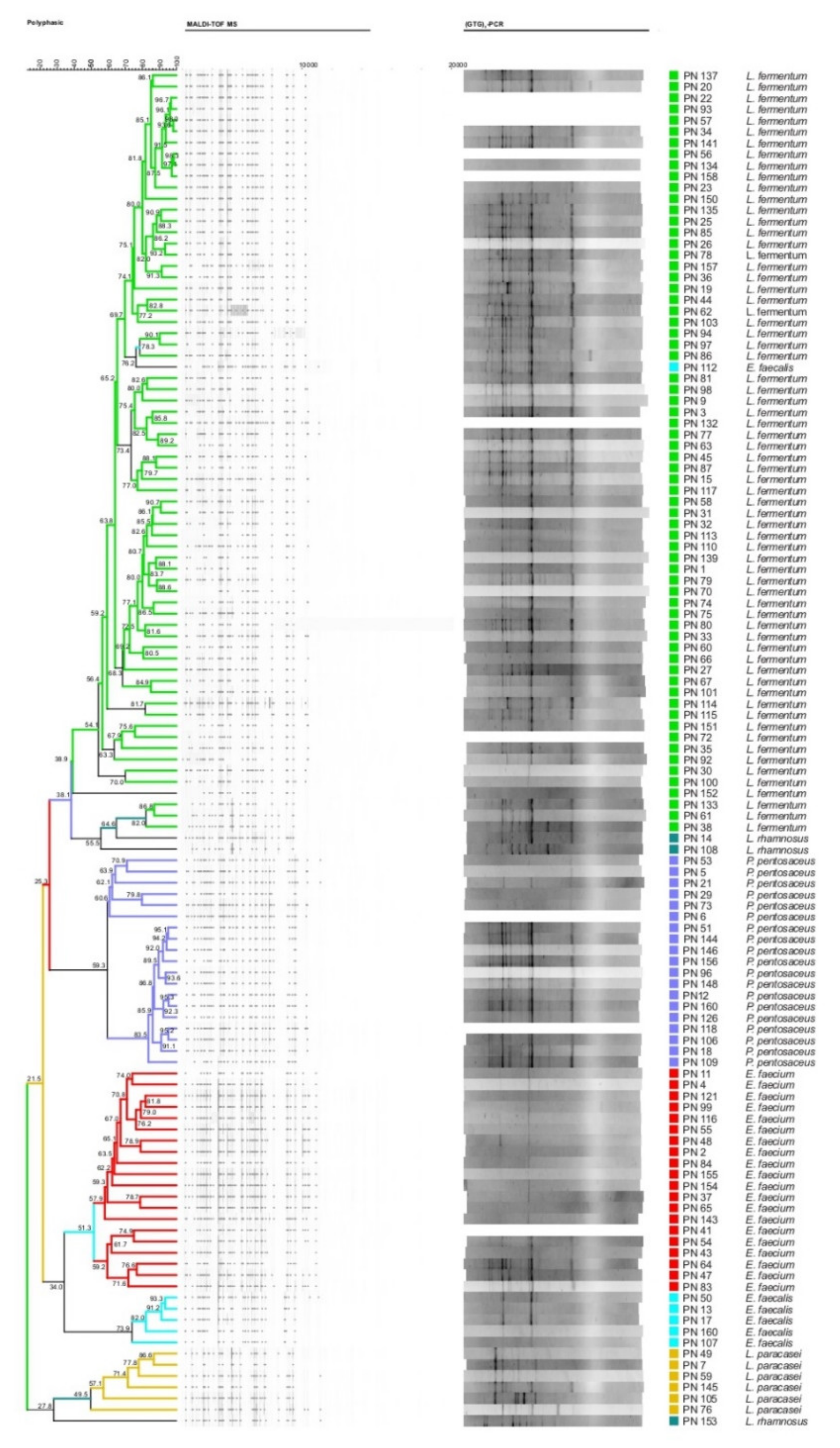
2.2. NSLAB Identification
2.3. Safety Assessment
2.3.1. DNAse, Gelatinase, Haemolytic Activities and Mucin Degradation Ability
2.3.2. Biogenic Amine Production
2.3.3. Antibiotic Susceptibility and Minimum Inhibitory Concentration Determination
2.3.4. Genes for Virulence, Biogenic Amines, and Antibiotic Resistance
2.4. Technological Features
2.4.1. Salt Tolerance
2.4.2. Proteolytic and Lipolytic Activities, Exopolysaccharides and Diacetyl Production
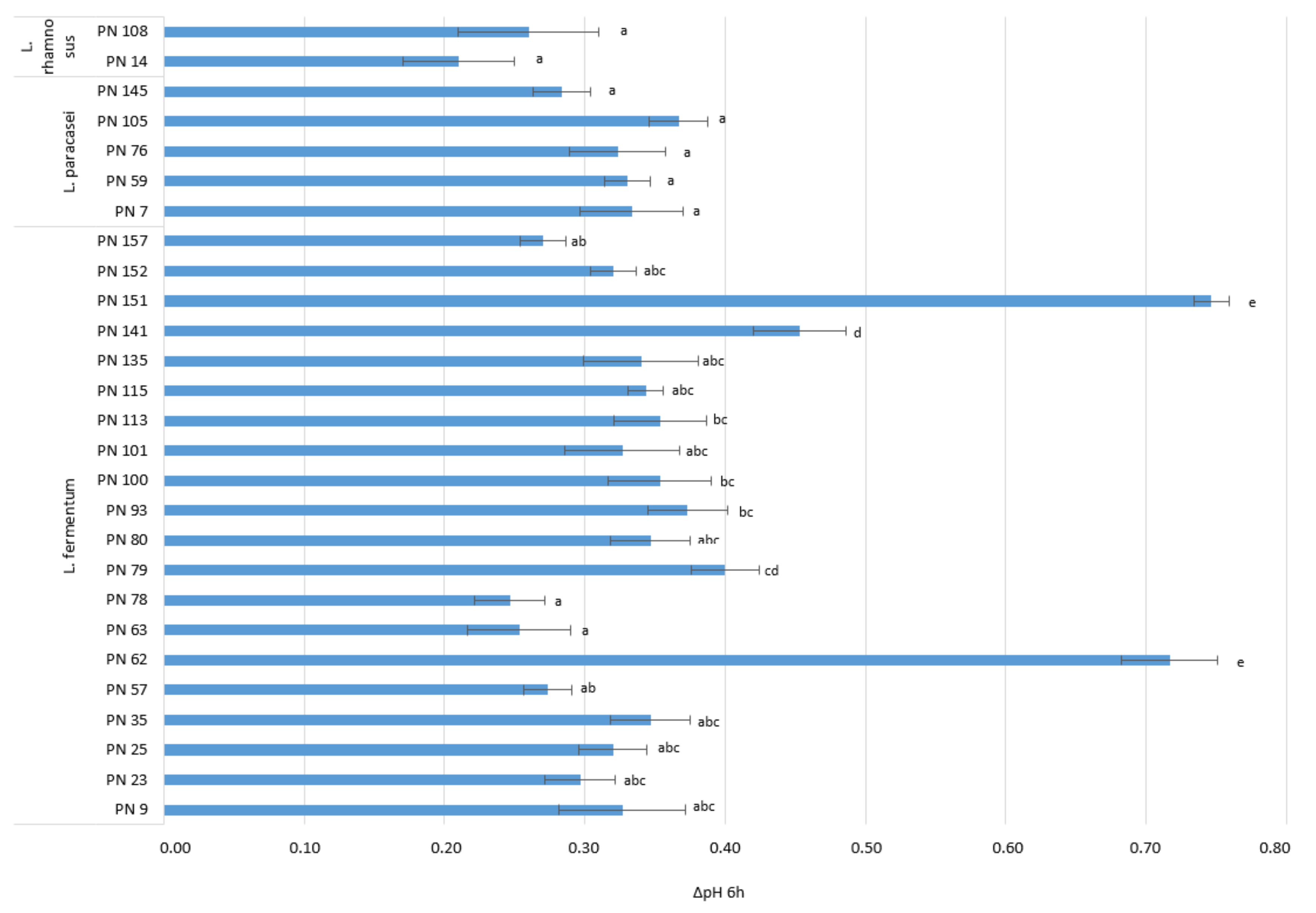
2.4.3. Acidifying Activity
2.4.4. Peptidase Activities
2.5. Experimental Pilot Scale Cheese-Making
2.6. Microbiological Analysis
2.7. Physico-Chemical Analysis
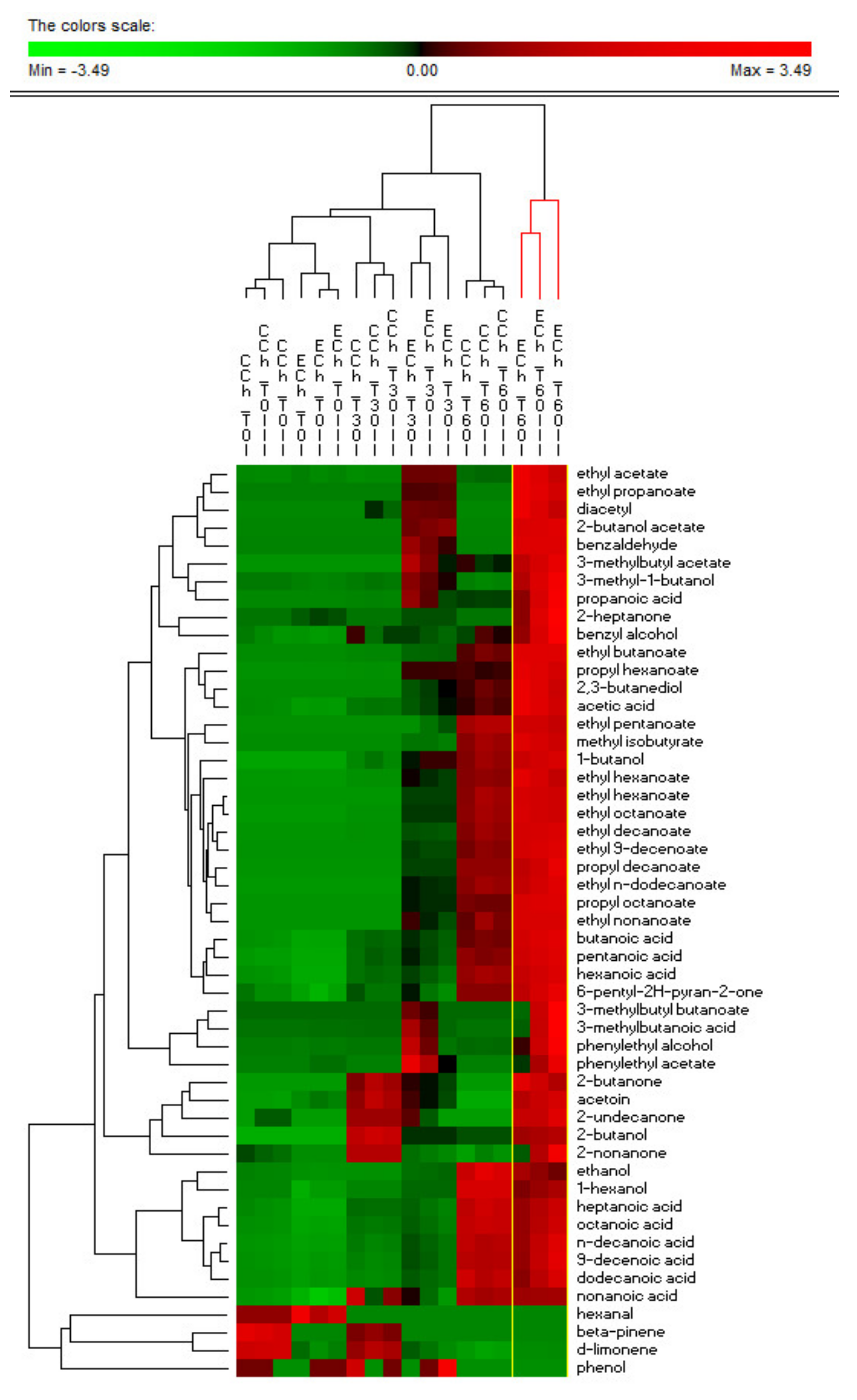
2.8. Analysis of Volatile Organic Compounds (VOCs)
2.9. Statistical Analysis
3. Results
3.1. NSLAB Identification
3.2. Safety Properties
3.3. Technological Features
3.3.1. Salt Tolerance
3.3.2. Proteolytic and Lipolytic Activities, Exopolysaccharides and Diacetyl Production Abilities
3.3.3. Acidifying Activity
3.3.4. Peptidase Activities
3.4. Microbiological Analysis of Control and Experimental Cheeses
3.5. Physico-Chemical Analysis
3.6. Volatile Organic Compounds (VOCs) Detection
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| CCh | control cheese |
| CFE | cell free extract |
| ECh | experimental cheese |
| EPS | exopolysaccharides |
| FFAs | free fatty acids |
| LAB | lactic acid bacteria |
| MALDI-TOF MS | matrix-assisted laser desorption/ionisation time-of-flight mass spectrometry |
| MIC | minimum inhibitory concentration |
| NSLAB | non-starter LAB |
| PDO | Protected Designation of Origin |
| PN | Provola dei Nebrodi |
| pNA | p-nitroanilide |
| QPS | qualified presumption of safety |
| SLAB | starter LAB |
| VOCs | volatile organic compounds |
References
- Meng, Z.; Zhang, L.; Xin, L.; Lin, K.; Yi, H.; Han, X. Technological characterization of Lactobacillus in semihard artisanal goat cheeses from different Mediterranean areas for potential use as nonstarter lactic acid bacteria. J. Dairy Sci. 2018, 101, 2887–2896. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carafa, I.; Nardin, T.; Larcher, R.; Viola, R.; Tuohy, K.; Franciosi, E. Identification and characterization of wild lactobacilli and pediococci from spontaneously fermented mountain cheese. Food Microbiol. 2015, 48, 123–132. [Google Scholar] [CrossRef] [PubMed]
- Di Grigoli, A.; Francesca, N.; Gaglio, R.; Guarrasi, V.; Moschetti, M.; Scatassa, M.L.; Settanni, L.; Bonanno, A. The influence of the wooden equipment employed for cheese manufacture on the characteristics of a traditional stretched cheese during ripening. Food Microbiol. 2015, 46, 81–91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khattab, A.R.; Guirguis, H.A.; Tawfik, S.M.; Farag, M.A. Cheese ripening: A review on modern technologies towards flavor enhancement, process acceleration and improved quality assessment. Trends Food Sci. Technol. 2019, 88, 343–360. [Google Scholar] [CrossRef]
- Beresford, T.P.; Fitzsimons, N.A.; Brennan, N.L.; Cogan, T.M. Recent advances in cheese microbiology. Int. Dairy J. 2001, 11, 259–274. [Google Scholar] [CrossRef]
- Yvon, M.; Rejien, L. Cheese flavour formation by amino acid catabolism. Int. Dairy J. 2001, 11, 185–201. [Google Scholar] [CrossRef]
- McSweeney, P.L.H.; Sousa, M.J. Biochemical pathways for the production of flavour compounds in cheeses during ripening. A review. Le Lait 2000, 80, 293–324. [Google Scholar] [CrossRef]
- Settanni, L.; Moschetti, G. Non-starter lactic acid bacteria used to improve cheese quality and provide health benefits. Food Microbiol. 2010, 27, 691–697. [Google Scholar] [CrossRef]
- Savijoki, K.; Ingmer, H.; Varmanen, P. Proteolytic systems of lactic acid bacteria. Appl. Microbiol. Biotechnol. 2006, 71, 394–406. [Google Scholar] [CrossRef]
- Čaplová, Z.; Pangallo, D.; Kraková, L.; Puškárová, A.; Drahovská, H.; Bučková, M.; Koreňová, J.; Kuchta, T. Detection of genes prtP, pepN, pepX and bcaT involved in formation of aroma-active compounds in lactic acid bacteria from ewes’ cheese. J. Food Nutr. Res. 2018, 57, 195–200. [Google Scholar]
- Curioni, P.M.G.; Bosset, J.O. Key odorants in various cheese types as determined by gas chromatography-olfactometry. Int. Dairy J. 2002, 12, 959–984. [Google Scholar] [CrossRef]
- Gobbetti, M.; De Angelis, M.; Di Cagno, R.; Mancini, L.; Fox, P.F. Pros and cons for using non-starter lactic acid bacteria (NSLAB) as secondary/adjunct starters for cheese ripening. Trends Food. Sci. Technol. 2015, 45, 167–178. [Google Scholar] [CrossRef]
- Pino, A.; Van Hoorde, K.; Pitino, I.; Russo, N.; Carpino, S.; Caggia, C.; Randazzo, C.L. Survival of potential probiotic lactobacilli used as adjunct cultures on Pecorino Siciliano cheese ripening and passage through the gastrointestinal tract of healthy volunteers. Int. J. Food Microbiol. 2017, 252, 42–52. [Google Scholar] [CrossRef] [PubMed]
- Pino, A.; Liotta, L.; Randazzo, C.L.; Todaro, A.; Mazzaglia, A.; De Nardo, F.; Chiofalo, V.; Caggia, C. Polyphasic approach to study physico-chemical, microbiological and sensorial characteristics of artisanal Nicastrese goat’s cheese. Food Microbiol. 2018, 70, 143–154. [Google Scholar] [CrossRef]
- Nguyen, T.L.D.; Van Hoorde, K.; Cnockaert, M.; De Brandt, E.; Aerts, M.; Le, B.T.; Vandamme, P. A description of the lactic acid bacteria microbiota associated with the production of traditional fermented vegetables in Vietnam. Int. J. Food Microbiol. 2013, 163, 19–27. [Google Scholar] [CrossRef] [PubMed]
- Pino, A.; Russo, N.; Van Hoorde, K.; De Angelis, M.; Sferrazzo, G.; Randazzo, C.L.; Caggia, C. Piacentinu Ennese PDO Cheese as Reservoir of Promising Probiotic Bacteria. Microorganisms 2019, 7, 254. [Google Scholar] [CrossRef] [Green Version]
- Muñoz-Atienza, E.; Gómez-Sala, B.; Araújo, C.; Campanero, C.; del Campo, R.; Hernández, P.E. Antimicrobial activity, antibiotic susceptibility and virulence factors of lactic acid bacteria of aquatic origin intended for use as probiotics in aquaculture. BMC Microbiol. 2013, 13, 15. [Google Scholar] [CrossRef] [Green Version]
- Pino, A.; Bartolo, E.; Caggia, C.; Cianci, A.; Randazzo, C.L. Detection of vaginal lactobacilli as probiotic candidates. Sci. Rep. 2019, 9, 3355. [Google Scholar] [CrossRef]
- Bover-Cid, S.; Holzapfel, W.H. Improved screening procedure for biogenic amine production by lactic acid bacteria. Int. J. Food Microbiol. 1999, 53, 33–41. [Google Scholar] [CrossRef]
- EFSA Panel on Additives and Products or Substances used in Animal Feed (FEEDAP). Guidance on the assessment of bacterial susceptibility to antimicrobials of human and veterinary importance. EFSA J. 2012, 10, 2740. [Google Scholar]
- Russo, N.; Caggia, C.; Pino, A.; Coque, T.M.; Arioli, S.; Randazzo, C.L. Enterococcus spp. in Ragusano PDO and Pecorino Siciliano cheese types: A snapshot of their antibiotic resistance distribution. Food Chem. Toxicol. 2018, 120, 277–286. [Google Scholar] [CrossRef] [PubMed]
- Ferrari, I.S.; de Souza, J.V.; Ramos, C.L.; da Costa, M.M.; Schwan, R.F.; Dias, F.S. Selection of autochthonous lactic acid bacteria from goat dairies and their addition to evaluate the inhibition of Salmonella typhi in artisanal cheese. Food Microbiol. 2016, 60, 29–38. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, S.C.; Coelho, M.C.; Todorov, S.D.; Franco, B.D.; Dapkevicius, M.L.; Silva, C.C. Technological properties of bacteriocin-producing lactic acid bacteria isolated from Pico cheese an artisanal cow’s milk cheese. J. Appl. Microbiol. 2014, 116, 573–585. [Google Scholar] [CrossRef] [PubMed]
- González, L.; Sacristán, N.; Arenas, R.; Fresno, J.M.; Tornadijo, E.M. Enzymatic activity of lactic acid bacteria (with antimicrobial properties) isolated from a traditional Spanish cheese. Food Microbiol. 2010, 27, 592–597. [Google Scholar] [CrossRef]
- Association of Official Analytical Chemists (AOAC). Official Methods of Analysis, 21st ed.; AOAC International: Gaithersburg, MD, USA, 2019. [Google Scholar]
- Delgado, F.J.; González-Crespo, J.; Cava, R.; Ramírez, R. Formation of the aroma of a raw goat milk cheese during maturation analysed by SPME–GC–MS. Food Chem. 2011, 129, 1156–1163. [Google Scholar] [CrossRef]
- Montemurro, M.; Celano, G.; De Angelis, M.; Gobbetti, M.; Rizzello, C.G.; Pontonio, E. Selection of non-Lactobacillus strains to be used as starters for sourdough fermentation. Food Microbiol. 2020, 90, 103491. [Google Scholar] [CrossRef]
- Sgarbi, E.; Lazzi, C.; Tabanelli, G.; Gatti, M.; Neviani, E.; Gardini, F. Nonstarter lactic acid bacteria volatilomes produced using cheese components. J. Dairy Sci. 2013, 96, 4223–4234. [Google Scholar] [CrossRef] [Green Version]
- Peralta, G.H.; Bergamini, C.V.; Wolf, I.V.; Perotti, M.C.; Hynes, E.R. Adjunct cultures from non-starter lactic acid bacteria. In Cheese Production, Consumption and Health Benefits; Henriques, M.H.F., Pereira, C.J.D., Eds.; Nova Science Publishers, Inc.: Hauppauge, NY, USA, 2017; Volume 1, pp. 39–96. [Google Scholar]
- Pintado, M.; Da Cruz, A.G.; De Sá, P.B.Z.R. Cheese microbiology. In Dairy Microbiology and Biochemistry: Recent Developments; Özer, B.H., Akdemir-Evrendilek, G., Eds.; CRC Press/Taylor & Francis: Boca Raton, FL, USA, 2014; Volume 1, pp. 113–133. [Google Scholar]
- Smit, G.; Smit, B.A.; Engels, W.J.M. Flavour formation by lactic acid bacteria and biochemical flavour profiling of cheese products. FEMS Microbiol. Rev. 2005, 29, 591–610. [Google Scholar] [CrossRef]
- Van Hoorde, K.; Van Landschoot, A. Application of adjunct-cultures and their influence on the sensory properties of hard and semi-hard cheese varieties. In Book on Beneficial Microbes in Fermented and Functional Foods; Rai, V.R., Bai, J.A., Eds.; CRC Press/Taylor & Francis Group: Boca Raton, FL, USA, 2014; pp. 531–550. [Google Scholar]
- Randazzo, C.L.; De Luca, S.; Todaro, A.; Restuccia, C.; Lanza, C.M.; Spagna, G.; Caggia, C. Preliminary characterization of wild lactic acid bacteria and their abilities to produce flflflavour compounds in ripened model cheese system. J. Appl. Microbiol. 2007, 103, 427–443. [Google Scholar] [CrossRef]
- Randazzo, C.L.; Pitino, I.; Ribbera, A.; Caggia, C. Pecorino Crotonese cheese: Study of bacterial population and flavour compounds. Food Microbiol. 2010, 27, 363–374. [Google Scholar] [CrossRef]
- Guarcello, R.; Carpino, S.; Gaglio, R.; Pino, A.; Rapisarda, T.; Caggia, C.; Marino, G.; Randazzo, C.L.; Settanni, L.; Todaro, M. A large factory-scale application of selected autochthonous lactic acid bacteria for PDO Pecorino Siciliano cheese production. Food Microbiol. 2016, 59, 66–75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carpino, S.; Randazzo, C.L.; Pino, A.; Russo, N.; Rapisarda, T.; Belvedere, G.; Caggia, C. Influence of PDO Ragusano cheese biofilm microbiota on flavour compounds formation. Food Microbiol. 2017, 61, 126–135. [Google Scholar] [CrossRef] [PubMed]
- Aquilanti, L.; Silvestri, G.; Zannini, E.; Osimani, A.; Santarelli, S.; Clementi, F. Phenotypic, genotypic and technological characterization of predominant lactic acid bacteria in Pecorino cheese from central Italy. J. Appl. Microbiol. 2007, 103, 948–960. [Google Scholar] [CrossRef] [PubMed]
- Nandan, A.; Nampoothiri, K.M. Therapeutic and biotechnological applications of substrate specific microbial aminopeptidases. Appl. Microbiol. Biotechnol. 2020, 28, 1–15. [Google Scholar] [CrossRef]
- Piraino, P.; Zotta, T.; Ricciardi, A.; McSweeney, P.L.H.; Parente, E. Acid production, proteolysis, autolytic and inhibitory properties of lactic acid bacteria isolated from pasta filata cheeses: A multivariate screening study. Int. Dairy J. 2008, 18, 81–92. [Google Scholar] [CrossRef]
- Collins, Y.F.; McSweeney, P.L.H.; Wilkinson, M.G. Lipolysis and free fatty acid catabolism in cheese: A review of current knowledge. Int. Dairy J. 2003, 13, 841–866. [Google Scholar] [CrossRef]
- Deeth, H.C. Lipoprotein lipase and lipolysis in milk. Int. Dairy J. 2006, 16, 555–562. [Google Scholar] [CrossRef]
- Thierry, A.; Collins, Y.F.; Abeijón Mukdsi, M.C.; McSweeney, P.L.H.; Wilkinson, M.G.; Spinnler, H.E. Lipolysis and Metabolism of Fatty Acids in Cheese. Cheese 2017, 423–444. [Google Scholar]
- Stefanovic, E.; Kilcawley, K.N.; Roces, C.; Rea, M.C.; O’sullivan, M.; Sheehan, J.J.; McAuliffe, O. Evaluation of the potential of Lactobacillus paracasei adjuncts for flavor compounds development and diversification in short-aged cheddar cheese. Front. Microbiol. 2018, 9, 1506. [Google Scholar] [CrossRef]
- Frank, D.C.; Owen, C.M.; Patterson, J. Solid phase microextraction (SPME) combined with gas chromatography and olfactometry—Mass spectrometry for characterization of cheese aroma compounds. LWT-Food Sci. Technol. 2004, 37, 139–154. [Google Scholar] [CrossRef]
- Drake, M.A.; Miracle, R.E.; McMahon, D.J. Impact of fat reduction on flavour and flavour chemistry of Cheddar cheeses. J. Dairy Sci. 2010, 93, 81. [Google Scholar] [CrossRef] [PubMed]
- Gaglio, R.; Franciosi, E.; Todaro, A.; Guarcello, R.; Alfeo, V.; Randazzo, C.L.; Settanni, L.; Todaro, M. Addition of selected starter/non-starter lactic acid bacterial inoculums to stabilise PDO Pecorino Siciliano cheese production. Food Res. Int. 2020, 136, 109335. [Google Scholar] [CrossRef] [PubMed]
- Guarrasi, V.; Sannino, C.; Moschetti, M.; Bonanno, A.; Di Grigoli, A.; Settanni, L. The individual contribution of starter and non-starter lactic acid bacteria to the volatile organic compound composition of Caciocavallo Palermitano cheese. Int. J. Food Microbiol. 2017, 259, 35–42. [Google Scholar] [CrossRef] [PubMed]
- Shiota, M.; Iwasawa, A.; Suzuki-Iwashima, A.; Iida, F. Effects of Flavor and Texture on the Sensory Perception of Gouda—Type Cheese Varieties during Ripening Using Multivariate Analysis. J. Food Sci. 2015, 80, 2740–2750. [Google Scholar] [CrossRef] [PubMed]
- Ercolini, D.; Frisso, G.; Mauriello, G.; Salvatore, F.; Coppola, S. Microbial diversity in natural whey cultures used for the production of Caciocavallo Silano PDO cheese. Int. J. Food Microbiol. 2008, 124, 164–170. [Google Scholar] [CrossRef]
- Bellesia, F.; Pinetti, A.; Pagnoni, U.G.; Rinaldi, R.; Zucchi, C.; Caglioti, L.; Palyi, G. Volatile components of Grana Parmigiano-Reggiano type hard cheese. Food Chem. 2003, 83, 55–61. [Google Scholar] [CrossRef]
- Di Cagno, R.; Banks, J.; Sheehan, L.; Fox, P.F.; Brechany, E.Y.; Corsetti, A.; Gobbetti, M. Comparison of the microbiological, compositional, biochemical, volatile profile and sensory characteristics of three Italian PDO ewes’ milk cheeses. Int. Dairy J. 2003, 13, 961–972. [Google Scholar] [CrossRef]
- Di Cagno, R.; Upadhyay, V.K.; McSweeney, P.L.H.; Corbo, M.R.; Faccia, M.; Gobbetti, M. Microbiological, compositional and biochemical characterisation of PDO Canestrato Pugliese cheese. Ital. J. Food Sci. 2004, 16, 45–58. [Google Scholar]
- Liu, S.Q.; Holland, R.; Crow, V.L. Esters and their biosynthesis in fermented dairy products: A review. Int. Dairy J. 2004, 14, 923–945. [Google Scholar] [CrossRef]
- Pinho, O.; Mendes, E.; Alves, M.M.; Ferreira, I.M.P.L.V.O. Chemical, Physical, and Sensorial Characteristics of “Terrincho” Ewe Cheese: Changes During Ripening and Intravarietal Comparison. J. Dairy Sci. 2004, 87, 249–257. [Google Scholar] [CrossRef] [Green Version]
- Fernández-García, E.; Serrano, C.; Nuñez, M. Volatile fraction and sensory characteristics of Manchego cheese. 2. Seasonal variation. J. Dairy Res. 2002, 69, 595–604. [Google Scholar] [CrossRef] [PubMed]
- Izco, J.M.; Torre, P. Characterisation of volatile flavour compounds in Roncal cheese extracted by the ‘purge and trap’ method and analysed by GC–MS. Food Chem. 2000, 70, 409–417. [Google Scholar] [CrossRef]
- Bertuzzi, A.S.; McSweeney, P.L.H.; Rea, M.C.; Kilcawley, K.N. Detection of Volatile Compounds of Cheese and Their Contribution to the Flavor Profile of Surface-Ripened Cheese. Compr. Rev. Food Sci. Food 2018, 17, 371–390. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Antimicrobials | Species (No. of Tested Isolates) | Number of Strains which Showed the Indicated MIC Value (mg/L) a | Breakpoints (mg/L) b | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 0.06 | 0.12 | 0.25 | 0.5 | 1 | 2 | 4 | 8 | 16 | 32 | 64 | 128 | 256 | 512 | 1024 | |||
| Ampicillin | L. fermentum (67) | 7 | 13 | 23 | 24 | 2 | |||||||||||
| P. pentosaceus (16) | 13 | 3 | 4 | ||||||||||||||
| L. paracasei (6) | 1 | 5 | 4 | ||||||||||||||
| L. rhamnosus (3) | 3 | 4 | |||||||||||||||
| Gentamicin | L. fermentum (67) | 15 | 11 | 41 | 16 | ||||||||||||
| P. pentosaceus (16) | 2 | 3 | 5 | 6 | 16 | ||||||||||||
| L. paracasei (6) | 2 | 4 | 32 | ||||||||||||||
| L. rhamnosus (3) | 3 | 16 | |||||||||||||||
| Kanamycin | L. fermentum (67) | 3 | 21 | 43 | 32 | ||||||||||||
| P. pentosaceus (16) | 6 | 10 | 64 | ||||||||||||||
| L. paracasei (6) | 6 | 64 | |||||||||||||||
| L. rhamnosus (3) | 3 | 64 | |||||||||||||||
| Streptomycin | L. fermentum (67) | 13 | 21 | 18 | 15 | 64 | |||||||||||
| P. pentosaceus (16) | 16 | 64 | |||||||||||||||
| L. paracasei (6) | 1 | 5 | 64 | ||||||||||||||
| L. rhamnosus (3) | 1 | 2 | 32 | ||||||||||||||
| Erythromycin | L. fermentum (67) | 15 | 21 | 28 | 1 | ||||||||||||
| P. pentosaceus (16) | 4 | 6 | 2 | 4 | 1 | ||||||||||||
| L. paracasei (6) | 4 | 2 | 1 | ||||||||||||||
| L. rhamnosus (3) | 2 | 1 | 1 | ||||||||||||||
| Clindamycin | L. fermentum (67) | 14 | 53 | 1 | |||||||||||||
| P. pentosaceus (16) | 12 | 4 | 1 | ||||||||||||||
| L. paracasei (6) | 4 | 1 | |||||||||||||||
| L. rhamnosus (3) | 3 | 1 | |||||||||||||||
| Tetracycline | L. fermentum (67) | 51 | 16 | 8 | |||||||||||||
| P. pentosaceus (16) | 16 | 8 | |||||||||||||||
| L. paracasei (6) | 6 | 4 | |||||||||||||||
| L. rhamnosus (3) | 3 | 8 | |||||||||||||||
| Chloramphenicol | L. fermentum (67) | 51 | 16 | 4 | |||||||||||||
| P. pentosaceus (16) | 11 | 5 | 4 | ||||||||||||||
| L. paracasei (6) | 6 | 4 | |||||||||||||||
| L. rhamnosus (3) | 2 | 1 | 4 | ||||||||||||||
| Species | Strains | Salt Tolerance * | Proteolytic Activity * | Lipolytic Activity * | EPS Production * | Diacetyl Production ** | ||
|---|---|---|---|---|---|---|---|---|
| 2% | 6% | 10% | ||||||
| Limosilactobacillus fermentum (n = 67) | PN 1; PN 3; PN 22; PN 26; PN 30; PN 31; PN 33; PN 34; PN 36; PN 44; PN 45; PN 56; PN 58; PN 61; PN 66; PN 70; PN 72; PN 75; PN 77; PN 85; PN 86; PN 87; PN 92; PN 97; PN 98; PN 103; PN 134; PN 137; PN 139; PN 150. | + | + | − | − | − | − | − |
| PN 9; PN 23; PN 25; PN 35; PN 57; PN 62; PN 63; PN 78; PN 79; PN 80; PN 93: PN 100; PN 101; PN 113; PN 115; PN 135; PN 141; PN 151; PN 152; PN 157. | + | + | + | + | − | + | ++ | |
| PN 15; PN 20; PN 27; PN 32; PN 67; PN 94; PN 117; PN 132; PN 133. | + | − | − | − | + | − | − | |
| PN 19; PN 38; PN 60; PN 74; PN 110; PN 114. | + | + | − | − | − | + | − | |
| PN 81; PN 158. | + | − | − | + | − | − | + | |
| Pediococcus penstosaceus (n = 10) | PN 5; PN 53; PN 146. | + | − | − | + | + | − | − |
| PN 12; PN 109. | + | − | − | + | − | − | − | |
| PN 16; PN 106; PN 144; PN 148; PN 156. | + | + | − | − | − | − | − | |
| Lacticaseibacillus paracasei (n = 6) | PN 7; PN 59; PN 76; PN 105; PN 145. | + | + | + | + | − | + | +++ |
| PN 49. | + | + | − | + | − | − | − | |
| Lacticaseibacillus rhamnosus (n = 3) | PN 14; PN 108. | + | + | + | + | + | + | ++ |
| PN 153. | + | + | − | + | − | + | − | |
| Species | Strain | Pep N | Pep A | Pep L | Pep M | Pep I |
|---|---|---|---|---|---|---|
| L. fermentum | PN 9 | 76.77 c ± 0.54 | 0.16 hm ± 0.04 | 7.73 n ± 0.14 | 19.23 j ± 0.17 | 35.63 d ± 0.36 |
| PN 23 | 42.08 d ± 0.17 | 0.89 efg ± 0.07 | 0.25 q ± 0.05 | 24.21 fg ± 0.13 | 22.40 i ± 0.20 | |
| PN 25 | 0.5 l ± 0.12 | 2.29 a ± 0.09 | 26.44 d ± 0.33 | 27.74 d ± 0.22 | 24.77 g ± 0.18 | |
| PN 35 | 0.50 l ± 0.06 | 0.70 eghim ± 0.06 | 43.02 a ± 0.14 | 23.57 fg ± 0.32 | 20.18 k ± 0.13 | |
| PN 57 | 10.63 j ± 0.48 | 0.75 eghm ± 0.08 | 25.69 d ± 0.31 | 26.37 e ± 0.26 | 22.54 i ± 0.16 | |
| PN 62 | 17.41 i ± 0.47 | 1.85 b ± 0.09 | 14.11 j ± 0.13 | 18.93 j ± 0.16 | 7.60 q ± 0.21 | |
| PN 63 | 19.45 h ± 0.46 | 1.1 cf ± 0.10 | 19.27 f ± 0.36 | 5.58 pq ± 0.15 | 13.28 n ± 0.14 | |
| PN 78 | 0.4 7 l ± 0.12 | 1.28 c ± 0.14 | 17.57 h ± 0.35 | 18.37 j ± 0.15 | 28.23 f ± 0.09 | |
| PN 79 | 18.54 hi ± 0.38 | 0.70 eghim ± 0.07 | 20.55 eg ± 0.37 | 7.71 n ± 0.16 | 32.83 e ± 0.13 | |
| PN 80 | 6.52 k ± 0.29 | 1.85 b ± 0.10 | 5.95 a ± 0.07 | 30.63 c ± 0.44 | 16.87 m ± 0.26 | |
| PN 93 | 30.67 e ± 0.45 | 1.43 cd ± 0.10 | 10.68 lm ± 0.13 | 4.57 p ± 0.36 | 18.25 l ± 0.22 | |
| PN 100 | 0.49 l ± 0.07 | 0.79 eghm ± 0.12 | 17.37 h ± 0.35 | 21.42 h ± 0.27 | 10.29 a ± 0.14 | |
| PN 101 | 120.75 a ± 0.32 | 0.37 im ± 0.05 | 30.64 b ± 0.37 | 41.83 a ± 0.17 | 39.63 b ± 0.23 | |
| PN 113 | 0.48 l ± 0.07 | 0.71 eglm ± 0.10 | 29.26 c ± 0.05 | 7.12 nq ± 0.12 | 23.74 h ± 0.18 | |
| PN 115 | 0.45 l ± 0.06 | 0.38 hm ± 0.06 | 21.27 e ± 0.16 | 16.72 k ± 0.16 | 24.67 gh ± 0.27 | |
| PN 135 | ND | ND | 10.16 m ± 0.11 | 24.51 f ± 0.23 | 8.97 p ± 0.18 | |
| PN 141 | 0.46 l ± 0.07 | 0.67 eghm ± 0.09 | 13.07 k ± 0.16 | 5.29 pq ± 0.23 | 37.63 c ± 0.34 | |
| PN 151 | 0.44 l ± 0.06 | 0.66 ghm ± 0.10 | 13.22 j ± 0.16 | 16.65 k ± 0.14 | 20.37 jk ± 0.27 | |
| PN 152 | 0.46 l ± 0.16 | 0.64 ghm ± 0.06 | 19.57 fg ± 0.13 | 15.07 l ± 0.16 | 9.21 p ± 0.16 | |
| PN 157 | 0.42 l ± 0.04 | 1.68 bd ± 0.10 | 0.85 pq ± 0.09 | 14.74 l ± 0.17 | 19.69 k ± 0.26 | |
| L. paracasei | PN 7 | 0.45 l ± 0.06 | 1.19 ce ± 0.06 | 7.58 n ± 0.34 | 6.14 oq ± 0.14 | 21.30 j ± 0.25 |
| PN 59 | 26.54 g ± 0.45 | 0.88 efg ± 0.08 | 11.57 l ± 0.28 | 9.42 m ± 0.20 | 19.51 k ± 0.20 | |
| PN 76 | 96.65 b ± 0.34 | 0.48 hilnop ± 0.07 | 43.37 a ± 0.21 | 33.54 b ± 0.33 | 47.61 a ± 0.22 | |
| PN 105 | ND | ND | 0.25 q ± 0.08 | 19.63 j ± 0.31 | 17.69 lm ± 0.17 | |
| PN 145 | 29.32 f ± 0.23 | 0.44 mn ± 0.05 | 30.90 b ± 0.15 | 6.61 no ± 0.36 | 13.66 n ±0.18 | |
| L. rhamnosus | PN 14 | 0.44 l ± 0.07 | 0.83 efo ± 0.09 | 7.75 n ± 0.17 | 23.20 g ± 0.18 | 17.78 lm ± 0.19 |
| PN 108 | 0.76 l ± 0.07 | 0.39 mp ± 0.07 | 1.50 p ± 0.30 | 6.58 noq ± 0.21 | 6.48 r ± 0.22 |
| Media | ECh | CCh | ||||
|---|---|---|---|---|---|---|
| 0 | 30 | 60 | 0 | 30 | 60 | |
| PCA | 9.50 d ± 0.14 | 8.93 c ± 0.07 | 8.63 bc ± 0.08 | 8.34 b ± 0.05 | 8.84 c ± 0.07 | 7.96 a ± 0.10 |
| MRS | 9.29 c ± 0.07 | 8.77 b ± 0.14 | 8.74 b ± 0.06 | 7.80 a ± 0.05 | 7.88 a ± 0.10 | 7.63 a ± 0.09 |
| VRBA * | 1.19 ab ± 0.11 | 1.36 b ± 0.09 | 1.04 a ± 0.07 | 1.73 c ± 0.07 | 1.85 c ± 0.09 | 1.09 ab ± 0.08 |
| M17 | 9.23 d ± 0.06 | 8.87 c ± 0.09 | 8.13 b ± 0.06 | 7.72 a ± 0.07 | 7.84 a ± 0.07 | 7.60 a ± 0.10 |
| SDA | 2.86 b ± 0.07 | 2.30 a ± 0.13 | 2.27 a ± 0.07 | 2.32 a ± 0.08 | 2.26 a ± 0.07 | 2.18 a ± 0.07 |
| MSA | 1.64 b ± 0.10 | 1.13 a ± 0.09 | 1.04 a ± 0.07 | 2.29 c ± 0.12 | 1.24 a ± 0.06 | 1.03 a ± 0.09 |
| KAA | 5.23 a ± 0.09 | 5.38 a ± 0.11 | 5.31 a ± 0.06 | 5.26 a ± 0.10 | 5.34 a ± 0.09 | 5.28 a ± 0.06 |
| Parameters | ECh | CCh | ||||
|---|---|---|---|---|---|---|
| 0 | 30 | 60 | 0 | 30 | 60 | |
| pH | 5.65 b ± 0.02 | 5.52 b ± 0.01 | 5.43 a ± 0.08 | 5.81 b ± 0.03 | 5.66 a ± 0.01 | 5.48 a ± 0.10 |
| Moisture | 44.81 c ± 0.15 | 40.76 b ± 0.03 | 34.58 a ± 0.05 | 44.25 c ± 0.12 | 40.88 b ± 1.09 | 34.86 a ± 1.29 |
| Protein | 26.98 a ± 0.22 | 28.66 a ± 0.08 | 30.08 b ± 0.06 | 27.25 a ± 0.14 | 28.34 a ± 0.44 | 29.80 a ± 0.59 |
| Fat | 23.56 a ± 0.11 | 25.78 a ± 0.03 | 30.03 b ± 0.04 | 23.62 a ± 0.19 | 25.90 a ± 0.34 | 29.90 b ± 0.70 |
| Salt | 1.54 a ± 0.20 | 2.01 b ± 0.02 | 1.83 b ± 0.01 | 1.40 a ± 0.04 | 2.04 b ± 0.03 | 1.91 b ± 0.02 |
| Ash | 2.89 a ± 0.28 | 2.88 a ± 0.03 | 3.33 b ± 0.01 | 2.78 a ± 0.03 | 2.70 a ± 0.07 | 3.47 b ± 0.08 |
| Cheese Samples | ||||||
|---|---|---|---|---|---|---|
| VOCs | ECh | CCh | ||||
| 0 | 30 | 60 | 0 | 30 | 60 | |
| Organic acids | ||||||
| Hexanoic acid | 5.67 d ± 0.007 | 17.23 c ± 2.616 | 49.67 a ± 3.714 | 9.61 d ± 0.662 | 15.37 c ± 0.556 | 36.65 b ± 1.647 |
| Butanoic acid | 2.14 d ± 0.007 | 11.70 c ± 1.680 | 38.21 a ± 2.454 | 4.53 d ± 0.363 | 9.07 c ± 0.597 | 20.17 b ± 0.950 |
| Octanoic acid | 2.04 c ± 0.144 | 6.02 b ± 0.960 | 18.62 a ± 3.102 | 3.68 bc ± 0.271 | 5.44 bc ± 0.135 | 20.62 a ± 0.596 |
| Acetic acid | 0.59 d ± 0.095 | 5.00 bc ± 0.904 | 18.49 a ± 2.221 | 1.59 d ± 0.165 | 2.75 cd ± 0.117 | 7.27 b ± 0.800 |
| n-Decanoic acid | 0.89 b ± 0.063 | 2.57 b ± 0.421 | 8.10 a ± 1.809 | 1.21 b ± 0.075 | 1.91 b ± 0.169 | 7.63 a ± 0.341 |
| Propanoic acid | ND | 0.74 b ± 0.387 | 1.91 a ± 0.868 | ND | 0.03 b ± 0.005 | 0.39 b ± 0.010 |
| Nonanoic acid | 0.10 c ± 0.052 | 0.38 bc ± 0.133 | 0.79 a ± 0.008 | 0.25 c ± 0.015 | 0.71 ab ± 0.287 | 0.83 a ± 0.032 |
| Heptanoic acid | 0.07 c ± 0.002 | 0.23 b ± 0.043 | 0.72 a ± 0.109 | 0.16 c ± 0.02 b | 0.24 b ± 0.006 | 0.81 a ± 0.053 |
| 3-Methylbutanoic acid | 0.01 a ± 0.007 | 0.57 a ± 0.516 | 1.44 a ± 1.342 | ND | 0.04 a ± 0.006 | ND |
| Pentanoic acid | 0.06 d ± 0.002 | 0.26 c ± 0.041 | 0.76 a ± 0.049 | 0.09 d ± 0.003 | 0.21 c ± 0.020 | 0.47 b ± 0.016 |
| Dodecanoic acid | 0.07 b ± 0.007 | 0.19 b ± 0.027 | 0.65 a ± 0.162 | 0.10 b ± 0.006 | 0.12 b ± 0.004 | 0.69 a ± 0.058 |
| 9-Decenoic acid | 0.07 b ± 0.011 | 0.21 b ± 0.035 | 0.69 a ± 0.179 | 0.09 b ± 0.012 | 0.13 b ± 0.009 | 0.62 a ± 0.012 |
| Alcohols | ||||||
| Ethanol | 0.30 b ± 0.068 | 1.30 b ± 0.130 | 4.58 a ± 0.812 | 0.71 b ± 0.054 | 0.37 b ± 0.016 | 7.80 a ± 0.970 |
| 3-Methyl-1-butanol | 0.15 d ± 0.026 | 1.12 b ± 0.315 | 2.76 a ± 0.800 | 0.24 cd ± 0.004 | 0.23 cd ± 0.026 | 0.15 d ± 0.046 |
| Phenylethyl alcohol | 0.20 b ± 0.018 | 1.21 ab ± 0.947 | 2.36 a ± 1.470 | 0.19 b ± 0.006 | 0.24 b ± 0.001 | 0.35 b ± 0.007 |
| Benzyl alcohol | 0.12 b ± 0.018 | 0.32 b ± 0.044 | 1.14 a ± 0.423 | 0.17 b ± 0.048 | 0.37 b ± 0.111 | 0.44 b ± 0.090 |
| 2-Butanol | ND | 0.07 c ± 0.003 | 0.15 b ± 0.009 | ND | 0.18 a ± 0.013 | 0.06 c ± 0.002 |
| 1-Butanol | ND | 0.07 c ± 0.006 | 0.17 a ± 0.010 | ND | 0.02 d ± 0.002 | 0.11 b ± 0.007 |
| 2,3-Butanediol | ND | 0.04 c ± 0.008 | 0.19 a ± 0.030 | ND | ND | 0.07 b ± 0.007 |
| 1-Hexanol | 0.01 e ± 0.006 | 0.03 c ± 0.000 | 0.08 b ± 0.009 | 0.02 de ± 0.001 | 0.02 cd ± 0.002 | 0.12 a ± 0.002 |
| Phenol | 0.01 a ± 0.007 | 0.01 a ± 0.013 | ND | 0.01 a ± 0.006 | 0.01 a ± 0.010 | ND |
| Esters | ||||||
| Ethyl hexanoate | 0.11 d ± 0.012 | 10.12 c ± 1.033 | 34.23 a ± 4.790 | 0.47 d ± 0.039 | 1.47 d ± 0.139 | 19.8 b ± 0.682 |
| Ethyl octanoate | 0.08 d ± 0.002 | 3.98 c ± 0.023 | 14.14 a ± 0.523 | 0.14 d ± 0.014 | 0.52 d ± 0.002 | 9.31 b ± 0.808 |
| Ethyl butanoate | 0.03 d ± 0.002 | 0.65 c ± 0.049 | 5.16 a ± 0.044 | 0.06 d ± 0.002 | 0.14 d ± 0.018 | 2.12 b ± 0.286 |
| Ethyl decanoate | 0.04 d ± 0.003 | 0.57 c ± 0.030 | 2.74 a ± 0.081 | 0.04 d ± 0.001 | 0.07 d ± 0.003 | 1.65 b ± 0.171 |
| Propyl hexanoate | ND | 0.68 b ± 0.006 | 2.08 a ± 0.180 | ND | 0.02 c ± 0.005 | 0.69 b ± 0.042 |
| Ethyl acetate | 0.07 c ± 0.021 | 0.50 b ± 0.008 | 1.14 a ± 0.197 | 0.05 c ± 0.005 | 0.05 c ± 0.007 | 0.16 c ± 0.010 |
| Ethyl n-dodecanoate | ND | 0.26 c ± 0.015 | 0.90 a ± 0.100 | ND | ND | 0.57 b ± 0.044 |
| Ethyl propanoate | ND | 0.33 b ± 0.016 | 1.06 a ± 0.135 | ND | ND | ND |
| 3-Methylbutyl acetate | ND | 0.40 b ± 0.177 | 0.76 a ± 0.131 | ND | ND | 0.22 b ± 0.038 |
| Ethyl hexanoate | ND | 0.15 c ± 0.003 | 0.58 a ± 0.023 | ND | ND | 0.39 b ± 0.045 |
| 2-Butanol acetate | ND | 0.26 b ± 0.026 | 0.56 a ± 0.010 | ND | ND | ND |
| Propyl octanoate | ND | 0.12 c ± 0.006 | 0.46 a ± 0.011 | ND | ND | 0.21 b ± 0.004 |
| Methyl isobutyrate | ND | 0.03 c ± 0.000 | 0.37 a ± 0.029 | ND | ND | 0.23 b ± 0.022 |
| Ethyl 9-decenoate | ND | 0.07 c ± 0.008 | 0.34 a ± 0.016 | ND | ND | 0.18 b ± 0.018 |
| Phenylethyl acetate | 0.01 a ± 0.007 | 0.15 a ± 0.102 | 0.14 a ± 0.105 | ND | ND | ND |
| Ethyl nonanoate | ND | 0.0 c ± 0.005 | 0.14 a ± 0.001 | ND | ND | 0.07 b ± 0.011 |
| Ethyl pentanoate | ND | 0.01 b ± 0.012 | 0.10 a ± 0.007 | ND | ND | 0.08 c ± 0.007 |
| Propyl decanoate | ND | 0.02 c ± 0.001 | 0.08 a ± 0.013 | ND | ND | 0.05 b ± 0.000 |
| 3-Methylbutyl butanoate | ND | 0.02 a ± 0.015 | 0.07 a ± 0.072 | ND | ND | ND |
| Ketones | ||||||
| 2-Butanone | ND | 0.10 b ± 0.016 | 0.30 a ± 0.059 | ND | 0.21 c ± 0.039 | ND |
| Diacetyl | ND | 0.59 b ± 0.03 | 1.42 a ± 0.280 | ND | 0.16 c ± 0.160 | ND |
| Acetoin | 0.10 c ± 0.018 | 0.24 b ± 0.048 | 0.59 a ± 0.064 | 0.04 c ± 0.008 | 0.48 a ± 0.073 | ND |
| 2-Nonanone | 0.04 b ± 0.002 | 0.07 b ± 0.018 | 0.41 a ± 0.291 | 0.11 ab ± 0.028 | 0.41 a ± 0.001 | 0.03 b ± 0.030 |
| 2-Heptanone | 0.04 b ± 0.009 | 0.04 b ± 0.002 | 0.39 a ± 0.194 | ND | ND | ND |
| 2-Undecanone | ND | 0.01 c ± 0.009 | 0.04 a ± 0.003 | 0.01 c ± 0.005 | 0.03 b ± 0.002 | ND |
| Aldehydes | ||||||
| Hexanal | 0.05 a ± 0.012 | ND | ND | 0.03 b ± 0.001 | ND | ND |
| Benzaldehyde | ND | 0.03 b ± 0.014 | 0.07 a ± 0.001 | ND | ND | ND |
| Terpenes | ||||||
| beta-Pinene | ND | ND | ND | 0.11 a ± 0.008 | 0.05 b ± 0.004 | ND |
| d-Limonene | 0.23 cd ± 0.092 | 0.27 c ± 0.109 | 0.16 cd ± 0.005 | 1.53 a ± 0.034 | 1.15 b ± 0.128 | 0.04 d ± 0.039 |
| Others | ||||||
| 6-Pentyl-2H-pyran-2-one | 0.03 cd ± 0.003 | 0.05 c ± 0.012 | 0.15 a ± 0.02 | 0.04 d ± 0.001 | 0.05 c ± 0.007 | 0.10 b ± 0.001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Randazzo, C.L.; Liotta, L.; Angelis, M.D.; Celano, G.; Russo, N.; Hoorde, K.V.; Chiofalo, V.; Pino, A.; Caggia, C. Adjunct Culture of Non-Starter Lactic Acid Bacteria for the Production of Provola Dei Nebrodi PDO Cheese: In Vitro Screening and Pilot-Scale Cheese-Making. Microorganisms 2021, 9, 179. https://doi.org/10.3390/microorganisms9010179
Randazzo CL, Liotta L, Angelis MD, Celano G, Russo N, Hoorde KV, Chiofalo V, Pino A, Caggia C. Adjunct Culture of Non-Starter Lactic Acid Bacteria for the Production of Provola Dei Nebrodi PDO Cheese: In Vitro Screening and Pilot-Scale Cheese-Making. Microorganisms. 2021; 9(1):179. https://doi.org/10.3390/microorganisms9010179
Chicago/Turabian StyleRandazzo, Cinzia Lucia, Luigi Liotta, Maria De Angelis, Giuseppe Celano, Nunziatina Russo, Koenraad Van Hoorde, Vincenzo Chiofalo, Alessandra Pino, and Cinzia Caggia. 2021. "Adjunct Culture of Non-Starter Lactic Acid Bacteria for the Production of Provola Dei Nebrodi PDO Cheese: In Vitro Screening and Pilot-Scale Cheese-Making" Microorganisms 9, no. 1: 179. https://doi.org/10.3390/microorganisms9010179
APA StyleRandazzo, C. L., Liotta, L., Angelis, M. D., Celano, G., Russo, N., Hoorde, K. V., Chiofalo, V., Pino, A., & Caggia, C. (2021). Adjunct Culture of Non-Starter Lactic Acid Bacteria for the Production of Provola Dei Nebrodi PDO Cheese: In Vitro Screening and Pilot-Scale Cheese-Making. Microorganisms, 9(1), 179. https://doi.org/10.3390/microorganisms9010179