The Stable Matching Problem in TBEV Enzootic Circulation: How Important Is the Perfect Tick-Virus Match?


Abstract
:1. Introduction
2. Materials and Methods
2.1. Tick Sampling and Maintenance
2.2. Virus Cultivation
2.3. In Vitro Feeding
2.4. PCR
2.5. Statistical Methods
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Domingo, E.; Holland, J.J. Mutation rates and rapid evolution of RNA viruses. In The Evolutionary Biology of Viruses; Raven Press: New York, NY, USA, 1994; pp. 161–184. [Google Scholar]
- Drake, J.W.; Holland, J.J. Mutation rates among RNA viruses. Proc. Natl. Acad. Sci. USA 1999, 96, 13910–13913. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bogovič, P. Tick-borne encephalitis: A review of epidemiology, clinical characteristics, and management. World J. Clin. Cases 2015, 3, 430–441. [Google Scholar] [CrossRef]
- Völker, I.; Hoffman, B.; Nessler, J.; Baumgärtner, W.; Wohlsein, P. First tick-borne encephalitis in a dog resident in Northern Germany. Berl. Münch. Tierärztl. Wochenschr. 2017, 130, 114–160. [Google Scholar] [CrossRef]
- Süss, J. Tick-borne encephalitis 2010: Epidemiology, risk areas, and virus strains in Europe and Asia—An overview. Ticks Tick Borne Dis. 2011, 2, 2–15. [Google Scholar] [CrossRef] [PubMed]
- Beauté, J.; Spiteri, G.; Warns-Petit, E.; Zeller, H. Tick-borne encephalitis in Europe, 2012 to 2016. Eurosurveillance 2018, 23, 1800201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hellenbrand, W.; Kreusch, T.; Böhmer, M.; Wagner-Wiening, C.; Dobler, G.; Wichmann, O.; Altmann, D. Epidemiology of Tick-Borne Encephalitis (TBE) in Germany, 2001–2018. Pathogens 2019, 8, 42. [Google Scholar] [CrossRef] [Green Version]
- Pavlovsky, E. Fundamentals of the theory of natural focality of transmissible human diseases. Zh. Obshch. Biol. 1946, 7, 3–33. [Google Scholar]
- Michelitsch, A.; Wernike, K.; Klaus, C.; Dobler, G.; Beer, M. Exploring the Reservoir Hosts of Tick-Borne Encephalitis Virus. Viruses 2019, 11, 669. [Google Scholar] [CrossRef] [Green Version]
- Uzcátegui, N.Y.; Sironen, T.; Golovljova, I.; Jääskeläinen, A.; Välimaa, H.; Lundkvist, Å.; Plyusnin, A.; Vaheri, A.; Vapalahti, O. Rate of evolution and molecular epidemiology of tick-borne encephalitis virus in Europe, including two isolations from the same focus 44 years apart. J. Gen. Virol. 2012, 93, 786–796. [Google Scholar] [CrossRef]
- Lambrechts, L.; Halbert, J.; Durand, P.; Gouagna, L.C.; Koella, J.C. Host genotype by parasite genotype interactions underlying the resistance of anopheline mosquitoes to Plasmodium falciparum. Malar. J. 2005, 4, 3. [Google Scholar] [CrossRef] [Green Version]
- Ruzek, D.; Bell-Sakyi, L.; Kopecký, J.; Grubhoffer, L. Growth of tick-borne encephalitis virus (European subtype) in cell lines from vector and non-vector ticks. Virus Res. 2008, 137, 142–146. [Google Scholar] [CrossRef] [PubMed]
- Randolph, S.E.; Storey, K. Impact of Microclimate on Immature Tick-Rodent Host Interactions (Acari: Ixodidae): Implications for Parasite Transmission. J. Med. Entomol. 1999, 36, 741–748. [Google Scholar] [CrossRef] [PubMed]
- Cagnacci, F.; Bolzoni, L.; Rosà, R.; Carpi, G.; Hauffe, H.C.; Valent, M.; Tagliapietra, V.; Kazimirova, M.; Koci, J.; Stanko, M.; et al. Effects of deer density on tick infestation of rodents and the hazard of tick-borne encephalitis. I: Empirical assessment. Int. J. Parasitol. 2012, 42, 365–372. [Google Scholar] [CrossRef] [PubMed]
- Carpi, G.; Cagnacci, F.; Neteler, M.; Rizzoli, A. Tick infestation on roe deer in relation to geographic and remotely sensed climatic variables in a tick-borne encephalitis endemic area. Epidemiol. Infect. 2007, 136, 1416–1424. [Google Scholar] [CrossRef] [PubMed]
- Vor, T.; Kiffner, C.; Hagedorn, P.; Niedrig, M.; Rühe, F. Tick burden on European roe deer (Capreolus capreolus). Exp. Appl. Acarol. 2010, 51, 405–417. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lefèvre, T.; Vantaux, A.; Dabiré, K.R.; Mouline, K.; Cohuet, A. Non-Genetic Determinants of Mosquito Competence for Malaria Parasites. PLoS Pathog. 2013, 9, e1003365. [Google Scholar] [CrossRef]
- Boelke, M.; Bestehorn, M.; Marchwald, B.; Kubinski, M.; Liebig, K.; Glanz, J.; Schulz, C.; Dobler, G.; Monazahian, M.; Becker, S.C. First Isolation and Phylogenetic Analyses of Tick-Borne Encephalitis Virus in Lower Saxony, Germany. Viruses 2019, 11, 462. [Google Scholar] [CrossRef] [Green Version]
- Reed, L.; Muench, H. A simple method of estimating fifty per cent endpoints. Am. J. Epidemiol. 1938, 27, 493–497. [Google Scholar] [CrossRef]
- Liebig, K.; Boelke, M.; Grund, D.; Schicht, S.; Springer, A.; Strube, C.; Chitimia-Dobler, L.; Dobler, G.; Jung, K.; Becker, S.C. Tick populations from endemic and non-endemic areas in Germany show differential susceptibility to TBEV. Sci. Rep. 2020, 10, 1–10. [Google Scholar] [CrossRef]
- Schwaiger, M.; Cassinotti, P. Development of a quantitative real-time RT-PCR assay with internal control for the laboratory detection of tick borne encephalitis virus (TBEV) RNA. J. Clin. Virol. 2003, 27, 136–145. [Google Scholar] [CrossRef]
- Belova, O.A.; Burenkova, L.A.; Karganova, G.G. Different tick-borne encephalitis virus (TBEV) prevalences in unfed versus partially engorged ixodid ticks—Evidence of virus replication and changes in tick behavior. Ticks Tick Borne Dis. 2012, 3, 240–246. [Google Scholar] [CrossRef] [PubMed]
- Belova, O.A.; Litov, A.G.; Kholodilov, I.S.; Kozlovskaya, L.I.; Bell-Sakyi, L.; Romanova, L.; Karganova, G.G. Properties of the tick-borne encephalitis virus population during persistent infection of ixodid ticks and tick cell lines. Ticks Tick Borne Dis. 2017, 8, 895–906. [Google Scholar] [CrossRef] [PubMed]
- Kupča, A.M.; Essbauer, S.; Zoeller, G.; De Mendonça, P.G.; Brey, R.; Rinder, M.; Pfister, K.; Spiegel, M.; Doerrbecker, B.; Pfeffer, M.; et al. Isolation and molecular characterization of a tick-borne encephalitis virus strain from a new tick-borne encephalitis focus with severe cases in Bavaria, Germany. Ticks Tick Borne Dis. 2010, 1, 44–51. [Google Scholar] [CrossRef] [PubMed]
- Heinz, F.-X.; Stiasny, K. Chapter 2b: The molecular and antigenic structure of TBEV. In Tick-Borne Encephalitis—The Book; Global Health Press Pte Ltd.: Singapore, 2019. [Google Scholar] [CrossRef]
- Danet, L.; Beauclair, G.; Berthet, M.; Moratorio, G.; Gracias, S.; Tangy, F.; Choumet, V.; Jouvenet, N. Midgut barriers prevent the replication and dissemination of the yellow fever vaccine in Aedes aegypti. PLoS Negl. Trop. Dis. 2019, 13, e0007299. [Google Scholar] [CrossRef] [Green Version]
- Arias-Goeta, C.; Mousson, L.; Rougeon, F.; Failloux, A.-B. Dissemination and Transmission of the E1-226V Variant of Chikungunya Virus in Aedes albopictus Are Controlled at the Midgut Barrier Level. PLoS ONE 2013, 8, e57548. [Google Scholar] [CrossRef]
- Mitzel, D.N.; Best, S.M.; Masnick, M.F.; Porcella, S.F.; Wolfinbarger, J.B.; Bloom, M.E. Identification of genetic determinants of a tick-borne flavivirus associated with host-specific adaptation and pathogenicity. Virology 2008, 381, 268–276. [Google Scholar] [CrossRef] [Green Version]
- Pettersson, J.H.-O.; Golovljova, I.; Vene, S.; Jaenson, T.G.T. Prevalence of tick-borne encephalitis virus in Ixodes ricinus ticks in northern Europe with particular reference to Southern Sweden. Parasites Vectors 2014, 7, 102. [Google Scholar] [CrossRef] [Green Version]
- Ott, D.; Ulrich, K.; Ginsbach, P.; Öhme, R.; Bock-Hensley, O.; Falk, U.; Teinert, M.; Lenhard, T. Tick-borne encephalitis virus (TBEV) prevalence in field-collected ticks (Ixodes ricinus) and phylogenetic, structural and virulence analysis in a TBE high-risk endemic area in southwestern Germany. Parasites Vectors 2020, 13, 1–15. [Google Scholar] [CrossRef]
- Leggewie, M.; Badusche, M.; Rudolf, M.; Jansen, S.; Börstler, J.; Krumkamp, R.; Huber, K.; Krüger, A.; Schmidt-Chanasit, J.; Tannich, E.; et al. Culex pipiens and Culex torrentium populations from Central Europe are susceptible to West Nile virus infection. One Health 2016, 2, 88–94. [Google Scholar] [CrossRef] [Green Version]
- Vega-Rúa, A.; Marconcini, M.; Madec, Y.; Manni, M.; Carraretto, D.; Gomulski, L.M.; Gasperi, G.; Failloux, A.-B.; Malacrida, A.R. Vector competence of Aedes albopictus populations for chikungunya virus is shaped by their demographic history. Commun. Biol. 2020, 3, 1–13. [Google Scholar] [CrossRef]
- Bennett, K.E.; Flick, D.; Fleming, K.H.; Jochim, R.; Beaty, B.J.; Black, W.C. Quantitative Trait Loci That Control Dengue-2 Virus Dissemination in the Mosquito Aedes aegypti. Genetics 2005, 170, 185–194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ciota, A.T.; Chin, P.A.; Ehrbar, D.J.; Micieli, M.V.; Fonseca, D.M.; Kramer, L.D. Differential Effects of Temperature and Mosquito Genetics Determine Transmissibility of Arboviruses by Aedes aegypti in Argentina. Am. J. Trop. Med. Hyg. 2018, 99, 417–424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steele, G.M.; Nuttall, P.A. Difference in vector competence of two species of sympatric ticks, Amblyomma variegatum and Rhipicephalus appendiculatus, for Dugbe virus (Nairovirus, Bunyaviridae). Virus Res. 1989, 14, 73–84. [Google Scholar] [CrossRef]


{kind=link}
| TBEV Area and Focus (Tick Origin) | Pairing (Virus Isolate) | TBEV Positive Ticks (%) | TBEV Negative Ticks (%) | Odds Ratio | 95%-CI | p-Value |
|---|---|---|---|---|---|---|
| Lower Saxony | Nonsynonymous | 114 (93%) | 8 (7%) | |||
| Synonymous | 61 (97%) | 2 (3%) | 1.85 | (0.33, 18.80) | 0.7204 | |
| Barsinghausen | Nonsynonymous (P19) | 60 (90%) | 7 (10%) | |||
| Synonymous (P51) | 24 (92%) | 2 (8%) | 1.18 | (0.19, 12.7) | 1.0000 | |
| Rauher Busch | Nonsynonymous (P51) | 54 (98%) | 1 (2%) | |||
| Synonymous (P19) | 37 (100%) | 0 (0%) | - | (0.02, -) | 1.0000 | |
| Bavaria | Nonsynonymous | 50 (82%) | 11 (18%) | |||
| Synonymous | 67 (99%) | 1 (1%) | 14.50 * | (2.00, 641.66) | 0.0014 | |
| Haselmühl | Nonsynonymous (HB171) | 35 (85%) | 6 (15%) | |||
| Synonymous (303/16) | 26 (96%) | 1 (4%) | 4.37 | (0.48, 212.53) | 0.2301 | |
| Heselbach | Nonsynonymous (303/16) | 15 (75%) | 5 (25%) | |||
| Synonymous (HB171) | 41 (100%) | 0 (0%) | - * | (2.15, -) | 0.0026 |
| TBEV Area and Focus (Tick Origin) | Pairing (Virus Isolate) | All Data | Outliers Removed | ||||
|---|---|---|---|---|---|---|---|
| Median | Minimum; Maximum | p-Value | Median | Minimum; Maximum | p-Value | ||
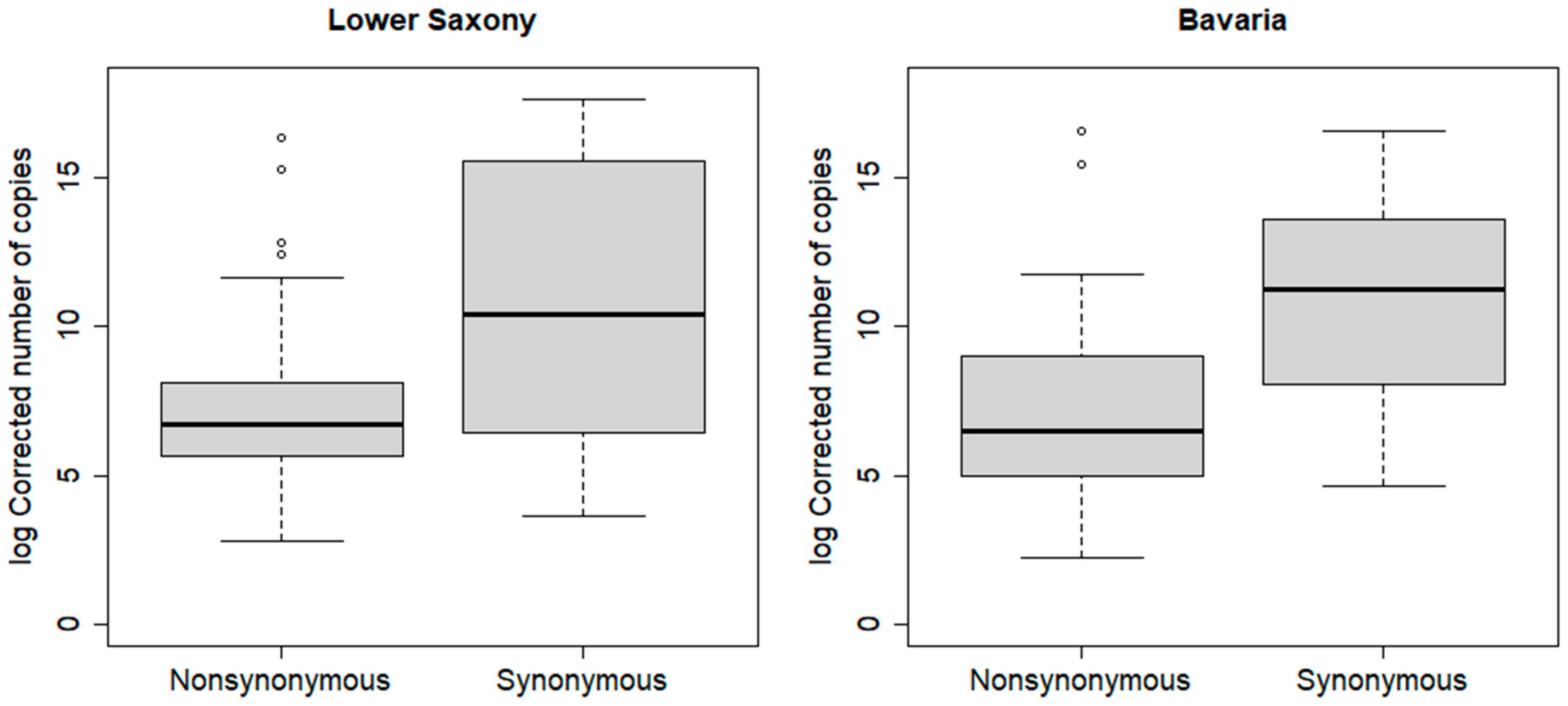
| Lower Saxony | Nonsynonymous | 842 | 16; 12,400,000 | <0.01 * | 781 | 16; 112,000 | 0.66 |
| Synonymous | 34,200 | 38; 46,100,000 | 915 | 38; 106,000 | |||
| Barsinghausen | Nonsynonymous (P19) | 2360 | 105; 250,000 | <0.01 * | 2195 | 105; 112,000 | 0.49 |
| Synonymous(P51) | 67,950 | 58; 46,100,000 | 15,300 | 58; 73,300 | |||
| Rauher Busch | Nonsynonymous (P51) | 376 | 16; 12,400,00038 | <0.01 * | 325 | 16; 107,000 | 0.0525 |
| Synonymous (P19) | 1650 | 38; 15,800,000 | 907 | 38; 106,000 | |||
| Bavaria | Nonsynonymous | 679 | 9; 15,500,000 | <0.01 * | 577 | 9; 128,000 | <0.01 * |
| Synonymous | 76,100 | 103; 15,500,000 | 9995 | 103; 15,500,000 | |||
| Haselmühl | Nonsynonymous (HB171) | 528 | 9; 15,500,000 | <0.01 * | 517 | 9; 128,000 | 0.07 |
| Synonymous (303/16) | 437,000 | 103; 10,100,000 | 2110 | 103; 77,700 | |||
| Heselbach | Nonsynonymous (303/16) | 1320 | 86; 11,900 | <0.01 * | 1320 | 86; 11,900 | <0.01 * |
| Synonymous (HB171) | 21,100 | 587; 15,500,000 | 16,900 | 587; 15,500,000 | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liebig, K.; Boelke, M.; Grund, D.; Schicht, S.; Bestehorn-Willmann, M.; Chitimia-Dobler, L.; Dobler, G.; Jung, K.; Becker, S.C. The Stable Matching Problem in TBEV Enzootic Circulation: How Important Is the Perfect Tick-Virus Match? Microorganisms 2021, 9, 196. https://doi.org/10.3390/microorganisms9010196
Liebig K, Boelke M, Grund D, Schicht S, Bestehorn-Willmann M, Chitimia-Dobler L, Dobler G, Jung K, Becker SC. The Stable Matching Problem in TBEV Enzootic Circulation: How Important Is the Perfect Tick-Virus Match? Microorganisms. 2021; 9(1):196. https://doi.org/10.3390/microorganisms9010196
Chicago/Turabian StyleLiebig, Katrin, Mathias Boelke, Domenic Grund, Sabine Schicht, Malena Bestehorn-Willmann, Lidia Chitimia-Dobler, Gerhard Dobler, Klaus Jung, and Stefanie C. Becker. 2021. "The Stable Matching Problem in TBEV Enzootic Circulation: How Important Is the Perfect Tick-Virus Match?" Microorganisms 9, no. 1: 196. https://doi.org/10.3390/microorganisms9010196
APA StyleLiebig, K., Boelke, M., Grund, D., Schicht, S., Bestehorn-Willmann, M., Chitimia-Dobler, L., Dobler, G., Jung, K., & Becker, S. C. (2021). The Stable Matching Problem in TBEV Enzootic Circulation: How Important Is the Perfect Tick-Virus Match? Microorganisms, 9(1), 196. https://doi.org/10.3390/microorganisms9010196