
Leishmania 360°: Guidelines for Exosomal Research

 , ,
, ,
Abstract
:1. Introduction
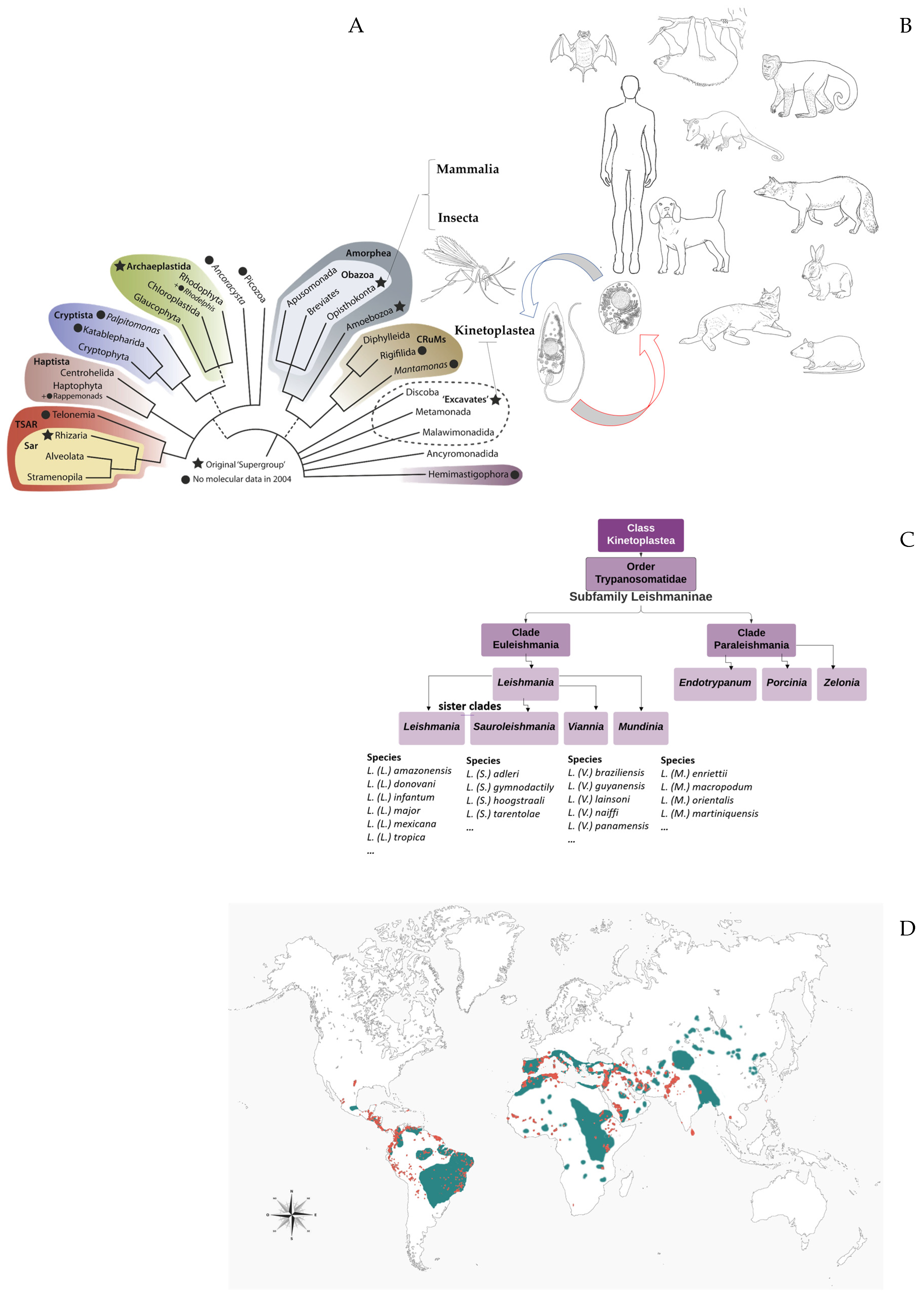
2. Leishmania (Kinetoplastida: Trypanosomatidae): Eclectic Parasites That Modulate the Host Immune Response through Extracellular Communication
2.1. Leishmanine Trypanosomatids Are Unicellular Organisms and Excellent Models to Explain Microbial Virulence
2.2. Host Leishmania Interactions through Extracellular Products
3. Leishmania Extracellular Vesicles (LEVs): Do We Know Everything?
3.1. Trypanosomatids Extracellular Vesicles Studies Requirements
3.2. Current and Expected Advances in Leishmania Extracellular Vesicle Research
3.3. Leishmania Extracellular Vesicles: What We Know So Far
3.4. Drug-Resistant Leishmania Extracellular Vesicles
4. Guidelines for the Production of Exosomes from Parasite Cells Grown in Culture Medium (Exo-Free Serum)
4.1. Guidelines for a Hypothetical Model of Leishmania Extracellular Vesicle Research
4.2. Guidelines for Leishmania Extracellular Vesicle Research
- Optimization of the cell culture and harvest conditions; (b) cell culture and exosome production, and the culture of adherent cells to confluence (e.g., RPMI 1640 medium with 10% ultrapure FBS, which provides the highest level of exosome depletion, 1 mM sodium-pyruvate in 225 cm2 cell culture bottles at 37 °C with 5% CO2); (c) removal of the medium from the confluent cells through the addition of 50 mL of fresh medium; (d) after 3 days, remove the cell-conditioned medium, with two centrifugation steps (300× g for 10 min at 2–8 °C and 2000× g for 30 min at 2–80 °C) prior to pre-enrichment [87].
- Test for exosome release; (b) depending on the cell type and the production efficiency, it may be possible to extract exosomes from the solution directly without any pre-enrichment step (e.g., flow analysis); (c) this is very useful to verify the efficiency of the exosome release by the cell and determine the correct time for exosome harvesting; (d) use of Dynabeads™ (CD9, CD63 or CD81) as exosomal markers, first in the host cell first and then in the LEVs (e.g., flow analysis) [87].
- Key questions asked by the extracellular research community; (b) current recommendations: estimating the size distribution using, e.g., Nano Tracking Analysis (NTA); (c) combining NTA with Transmission Electron Microscopy (TEM); (d) verifying the presence of miRNA/mRNA; (e) using Western Blotting (WB) to target lipid-bound targets such as tetraspanins (3 different types), cytosolic proteins, such as TSG101, annexins, and rabs, the absence or under-representation of the ER (e.g., hsp90B1, calnexin), Golgi (GM130), mitochondrion (cytC) or nucleus (histones); (f) flow cytometry can be used easily to support the WB data (e.g., for membrane-anchored targets such as tetraspanins) using Dynabeads™ as the solid support, which can be introduced and detected in the flow instrument [91].
- Consider the question “can the exosome harvest be used directly or do I need to pre-enrich?”; (b) the flow signal you will obtain may vary from very low/absent to signals that are sufficient for further analysis. Depending on this signal, the researcher may decide to continue to direct isolation with the Dynabeads™, give that they are compatible with many different types of downstream applications [91].
5. Discussion
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Barbosa, F.M.C.; Dupin, T.V.; Toledo, M.D.S.; Reis, N.F.D.C.; Ribeiro, K.; Cronemberger-Andrade, A.; Rugani, J.N.; De Lorenzo, B.H.P.; Brito, R.R.N.E.; Soares, R.P.; et al. Extracellular Vesicles Released by Leishmania (Leishmania) amazonensis Promote Disease Progression and Induce the Production of Different Cytokines in Macrophages and B-1 Cells. Front. Microbiol. 2018, 9, 3056. [Google Scholar] [CrossRef]
- Gioseffi, A.; Hamerly, T.; Van, K.; Zhang, N.; Dinglasan, R.R.; Yates, P.A.; Kima, P.E. Leishmania-infected macrophages release extracellular vesicles that can promote lesion development. Life Sci. Alliance 2020, 3, e202000742. [Google Scholar] [CrossRef]
- Douanne, N.; Dong, G.; Douanne, M.; Olivier, M.; Fernandez-Prada, C. Unravelling the proteomic signature of extracellular vesicles released by drug-resistant Leishmania infantum parasites. PLoS Negl. Trop. Dis. 2020, 14, e0008439. [Google Scholar] [CrossRef]
- Marshall, S.; Kelly, P.H.; Singh, B.K.; Pope, R.M.; Kim, P.; Zhanbolat, B.; Wilson, M.E.; Yao, C. Extracellular release of virulence factor major surface protease via exosomes in Leishmania infantum promastigotes. Parasites Vectors 2018, 11, 355. [Google Scholar] [CrossRef]
- Dong, G.; Wagner, V.; Minguez-Menendez, A.; Fernandez-Prada, C.; Olivier, M. Extracellular vesicles and leishmaniasis: Current knowledge and promising avenues for future development. Mol. Immunol. 2021, 135, 73–83. [Google Scholar] [CrossRef]
- Soto-Serna, L.E.; Diupotex, M.; Zamora-Chimal, J.; Ruiz-Remigio, A.; Delgado-Domínguez, J.; Cervantes-Sarabia, R.B.; Méndez-Bernal, A.; Escalona-Montaño, A.R.; Aguirre-García, M.M.; Becker, I. Leishmania mexicana: Novel Insights of Immune Modulation through Amastigote Exosomes. J. Immunol. Res. 2020, 2020, 8894549. [Google Scholar] [CrossRef] [PubMed]
- Cronemberger-Andrade, A.; Aragão-França, L.; Araujo, C.F.; Rocha, V.J.; Borges-Silva, M.C.; Figueiras, C.P.; Oliveira, P.R.; Freitas, L.A.R.; Veras, P.S.T.; Pontes-de-Carvalho, L. Extracellular vesicles from Leishmania-infected macrophages confer an anti-infection cytokine-production profile to naïve macrophages. PLoS Negl. Trop. Dis. 2014, 8, 11. [Google Scholar] [CrossRef] [Green Version]
- Olajide, J.S.; Cai, J. Perils and Promises of Pathogenic Protozoan Extracellular Vesicles. Front. Cell. Infect. Microbiol. 2020, 10, 371. [Google Scholar] [CrossRef] [PubMed]
- WHO. Leishmaniasis. Available online: https://www.who.int/health-topics/leishmaniasis#tab=tab_1 (accessed on 10 February 2021).
- Kumar, R.; Engwerda, C.R. Vaccines to prevent leishmaniasis. Clin. Transl. Immunol. 2014, 3, e13. [Google Scholar] [CrossRef] [PubMed]
- Aida, V.; Pliasas, V.C.; Neasham, P.J.; North, J.F.; McWhorter, K.L.; Glover, S.R.; Kyriakis, C.S. Novel Vaccine Technologies in Veterinary Medicine: A Herald to Human Medicine Vaccines. Front. Veter- Sci. 2021, 8, 654289. [Google Scholar] [CrossRef]
- Jamshaid, H.; Din, F.U.; Khan, G.M. Nanotechnology based solutions for anti-leishmanial impediments: A detailed insight. J. Nanobiotechnology 2021, 19, 106. [Google Scholar] [CrossRef] [PubMed]
- Kammona, O.; Tsanaktsidou, E. Nanotechnology-aided diagnosis, treatment and prevention of leishmaniasis. Int. J. Pharm. 2021, 605, 120761. [Google Scholar] [CrossRef] [PubMed]
- Dong, G.; Filho, A.L.; Olivier, M. Modulation of Host-Pathogen Communication by Extracellular Vesicles (EVs) of the Protozoan Parasite Leishmania. Front. Cell. Infect. Microbiol. 2019, 9, 100. [Google Scholar] [CrossRef] [PubMed]
- Yáñez-Mó, M.; Siljander, P.R.-M.; Andreu, Z.; Zavec, A.B.; Borras, F.E.; Buzas, E.I.; Buzas, K.; Casal, E.; Cappello, F.; Carvalho, J.; et al. Biological properties of extracellular vesicles and their physiological functions. J. Extracell. Vesicles 2015, 4, 27066. [Google Scholar] [CrossRef] [Green Version]
- Weber, B.; Franz, N.; Marzi, I.; Henrich, D.; Leppik, L. Extracellular vesicles as mediators and markers of acute organ injury: Current concepts. Eur. J. Trauma Emerg. Surg. 2021. [Google Scholar] [CrossRef]
- Carroll-Portillo, A.; Surviladze, Z.; Cambi, A.; Lidke, D.S.; Wilson, B.S.P. Mast cell synapses and exosomes: Membrane contacts for information exchange. Front. Immunol. 2012, 3, 46. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Liu, Y.; Liu, H.; Tang, W.H. Exosomes: Biogenesis, biologic function and clinical potential. Cell Biosci. 2019, 9, 19. [Google Scholar] [CrossRef]
- Doyle, L.M.; Wang, M.Z. Overview of Extracellular Vesicles, Their Origin, Composition, Purpose, and Methods for Exosome Isolation and Analysis. Cells 2019, 8, 727. [Google Scholar] [CrossRef] [Green Version]
- Gabriel, Á.; Bolas, A.S.; Marques, J.; Mourata-Gonçalves, P.; Ruas, P.; Dias-Guerreiro, T.; Santos-Gomes, G. Cutaneous Leishmaniasis: The Complexity of Host’s Effective Immune Response against a Polymorphic Parasitic Disease. J. Immunol. Res. 2019, 2019, 2603730. [Google Scholar] [CrossRef] [Green Version]
- Burki, F.; Roger, A.J.; Brown, M.W.; Simpson, A.G. The New Tree of Eukaryotes. Trends Ecol. Evol. 2020, 35, 43–55. [Google Scholar] [CrossRef] [Green Version]
- Akhoundi, M.; Kuhls, K.; Cannet, A.; Votypka, J.; Marty, P.; Delaunay, P.; Sereno, D. A Historical Overview of the Classification, Evolution, and Dispersion of Leishmania Parasites and Sandflies. PLOS Neglected Trop. Dis. 2016, 10, e0004349. [Google Scholar] [CrossRef] [PubMed]
- Klatt, S.; Simpson, L.; Maslov, D.A.; Konthur, Z. Leishmania tarentolae: Taxonomic classification and its application as a promising biotechnological expression host. PLOS Neglected Trop. Dis. 2019, 13, e0007424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sereno, D. Leishmania (Mundinia) spp.: From description to emergence as new human and animal Leishmania pathogens. New Microbes New Infect. 2019, 30, 100540. [Google Scholar] [CrossRef]
- Espinosa, O.A.; Serrano, M.G.; Camargo, E.; Teixeira, M.M.G.; Shaw, J.J. An appraisal of the taxonomy and nomenclature of trypanosomatids presently classified as Leishmania and Endotrypanum. Parasitology 2016, 145, 430–442. [Google Scholar] [CrossRef]
- Azami-Conesa, I.; Gómez-Muñoz, M.T.; Martínez-Díaz, R.A. A Systematic Review (1990–2021) of Wild Animals Infected with Zoonotic Leishmania. Microorganisms 2021, 9, 1101. [Google Scholar] [CrossRef]
- Ashford, R. The leishmaniases as emerging and reemerging zoonoses. Int. J. Parasitol. 2000, 30, 1269–1281. [Google Scholar] [CrossRef]
- Torres, E.; Quintanilla-Cedillo, M.R.; Ruiz-Esmenjaud, J.; Arenas, R. Leishmaniasis: A review. F1000Research 2017, 6, 750. [Google Scholar] [CrossRef]
- Mann, S.; Frasca, K.; Scherrer, S.; Henao-Martínez, A.F.; Newman, S.; Ramanan, P.; Suarez, J.A. A Review of Leishmaniasis: Current Knowledge and Future Directions. Curr. Trop. Med. Rep. 2021, 8, 121–132. [Google Scholar] [CrossRef]
- Gurunathan, S.; Kang, M.H.; Kim, J.H. A Comprehensive Review on Factors Influences Biogenesis, Functions, Therapeutic and Clinical Implications of Exosomes. Int. J. Nanomedicine. 2021, 16, 1281–1312. [Google Scholar] [CrossRef]
- Marcilla, A.; Jaular, L.M.; Trelis, M.; Menezes-Neto, A.; Osuna, A.; Bernal, D.; Fernandez-Becerra, C.; Almeida, I.; Del Portillo, H.A. Extracellular vesicles in parasitic diseases. J. Extracell. Vesicles 2014, 3, 25040. [Google Scholar] [CrossRef] [Green Version]
- Bifeld, E.; Clos, J. The genetics of Leishmania virulence. Med. Microbiol. Immunol. 2015, 204, 619–634. [Google Scholar] [CrossRef] [PubMed]
- Schwabl, P.; Boité, M.C.; Bussotti, G.; Jacobs, A.; Andersson, B.; Moreira, O.; Freitas-Mesquita, A.L.; Meyer-Fernandes, J.R.; Telleria, E.L.; Traub-Csekö, Y.; et al. Colonization and genetic diversification processes of Leishmania infantum in the Americas. Commun. Biol. 2021, 4, 139. [Google Scholar] [CrossRef]
- Elmahallawy, E.; Alkhaldi, A. Insights into Leishmania Molecules and Their Potential Contribution to the Virulence of the Parasite. Veter- Sci. 2021, 8, 33. [Google Scholar] [CrossRef]
- Forrest, D.M.; Batista, M.; Marchini, F.K.; Tempone, A.J.; Traub-Csekö, Y.M. Proteomic analysis of exosomes derived from procyclic and metacyclic-like cultured Leishmania infantum chagasi. J. Proteom. 2020, 227, 103902. [Google Scholar] [CrossRef]
- Maharana, B.R.; Tewari, A.K.; Singh, V. An overview on kinetoplastid paraflagellar rod. J. Parasit. Dis. Off. Organ Indian Soc. Parasitol. 2015, 39, 589–595. [Google Scholar] [CrossRef] [Green Version]
- Wijesinghe, H.; Gunathilaka, N.; Semege, S.; Pathirana, N.; Manamperi, N.; De Silva, C.; Fernando, D. Histopathology of Cutaneous Leishmaniasis Caused by Leishmania donovani in Sri Lanka. BioMed Res. Int. 2020, 2020, 4926819. [Google Scholar] [CrossRef]
- Nogueira, P.M.; De Menezes-Neto, A.; Borges, V.M.; Descoteaux, A.; Torrecilhas, A.C.; Xander, P.; Revach, O.-Y.; Regev-Rudzki, N.; Soares, R.P. Immunomodulatory Properties of Leishmania Extracellular Vesicles During Host-Parasite Interaction: Differential Activation of TLRs and NF-κB Translocation by Dermotropic and Viscerotropic Species. Front. Cell. Infect. Microbiol. 2020, 10, 380. [Google Scholar] [CrossRef]
- Giraud, E.; Martin, O.; Yakob, L.; Rogers, M. Quantifying Leishmania Metacyclic Promastigotes from Individual Sandfly Bites Reveals the Efficiency of Vector Transmission. Commun. Biol. 2019, 2, 84. [Google Scholar] [CrossRef] [PubMed]
- Lambertz, U.; Silverman, J.M.; Nandan, D.; McMaster, W.R.; Clos, J.; Foster, L.J.; Reiner, N.E. Secreted virulence factors and immune evasion in visceral leishmaniasis. J. Leukoc. Biol. 2012, 91, 887–899. [Google Scholar] [CrossRef]
- Sunter, J.; Gull, K. Shape, form, function and Leishmania pathogenicity: From textbook descriptions to biological understanding. Open Biol. 2017, 7, 170165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Efstathiou, A.; Smirlis, D. Leishmania Protein Kinases: Important Regulators of the Parasite Life Cycle and Molecular Targets for Treating Leishmaniasis. Microorganisms 2021, 9, 691. [Google Scholar] [CrossRef] [PubMed]
- Kalenkiewicz, A.; Cruz, P.; Dolan, M.; Sackett, D.L. Targeting Microtubules in Leishmania using a Combined Computational & Experimental Approach. Biophys. J. 2017, 112, 495a. [Google Scholar] [CrossRef]
- Silva, C.G.; Silva, R.M.M.; Diniz, V.A.; Côrte-Real, S. Ultraestrutura de Parasitos do Gênero Leishmania. In Leishmanioses do Continente Americano; Conceição-Silva, F., Alves, C.R., Eds.; Editora Fiocruz: Rio de Janeiro, Brazil, 2014; pp. 53–67. ISBN 978-85-7541-568-9. [Google Scholar] [CrossRef]
- Halliday, C.; Yanase, R.; Catta-Preta, C.M.C.; Moreira-Leite, F.; Myskova, J.; Pruzinova, K.; Volf, P.; Mottram, J.C.; Sunter, J.D. Role for the flagellum attachment zone in Leishmania anterior cell tip morphogenesis. PLoS Pathog. 2020, 16, e1008494. [Google Scholar] [CrossRef]
- Gadelha, A.P.R.; Cunha-E-Silva, N.L.; De Souza, W. Assembly of the Leishmania amazonensis flagellum during cell differentiation. J. Struct. Biol. 2013, 184, 280–292. [Google Scholar] [CrossRef] [Green Version]
- Beneke, T.; Demay, F.; Hookway, E.; Ashman, N.; Jeffery, H.; Smith, J.; Valli, J.; Becvar, T.; Myskova, J.; Lestinova, T.; et al. Genetic dissection of a Leishmania flagellar proteome demonstrates requirement for directional motility in sand fly infections. PLOS Pathog. 2019, 15, e1007828. [Google Scholar] [CrossRef] [Green Version]
- Emoradin, N.; Edescoteaux, A. Leishmania promastigotes: Building a safe niche within macrophages. Front. Cell. Infect. Microbiol. 2012, 2, 121. [Google Scholar] [CrossRef] [Green Version]
- Hofstatter, P.G.; Ribeiro, G.M.; Porfírio-Sousa, A.L.; Lahr, D.J.G. The Sexual Ancestor of all Eukaryotes: A Defense of the “Meiosis Toolkit”: A Rigorous Survey Supports the Obligate Link between Meiosis Machinery and Sexual Recombination. BioEssays 2020, 42, e2000037. [Google Scholar] [CrossRef]
- Shaik, J.S.; Dobson, D.E.; Sacks, D.L.; Beverley, S. M Leishmania Sexual Reproductive Strategies as Resolved through Computational Methods Designed for Aneuploid Genomes. Genes 2021, 12, 167. [Google Scholar] [CrossRef] [PubMed]
- Young, J.; Kima, P.E. The Leishmania Parasitophorous Vacuole Membrane at the Parasite-Host Interface. Yale. J. Biol. Med. 2019, 92, 511–521. [Google Scholar]
- Rybak, K.; Robatzek, S. Functions of Extracellular Vesicles in Immunity and Virulence. Plant. Physiol. 2019, 179, 1236–1247. [Google Scholar] [CrossRef] [Green Version]
- Soulat, D.; Bogdan, C. Function of Macrophage and Parasite Phosphatases in Leishmaniasis. Front. Immunol. 2017, 8, 1838. [Google Scholar] [CrossRef]
- Silverman, J.M.; Reiner, N.E.M. Leishmania Exosomes Deliver Preemptive Strikes to Create an Environment Permissive for Early Infection. Front. Cell. Infect. Microbiol. 2012, 1, 26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cardoso, L.; Schallig, H.; Persichetti, M.F.; Pennisi, M.G. New Epidemiological Aspects of Animal Leishmaniosis in Europe: The Role of Vertebrate Hosts Other Than Dogs. Pathogens 2021, 10, 307. [Google Scholar] [CrossRef] [PubMed]
- Conceição-Silva, F.; Morgado, F.N. Leishmania Spp-Host Interaction: There Is Always an Onset, but Is There an End? Front. Cell. Infect. Microbiol. 2019, 9, 330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maia, C.; Campino, L. Biomarkers Associated With Leishmania infantum Exposure, Infection, and Disease in Dogs. Front. Cell. Infect. Microbiol. 2018, 8, 302. [Google Scholar] [CrossRef] [PubMed]
- De Morais, R.C.S.; Gonçalves, S.D.C.; Costa, P.L.; Da Silva, K.G.; Da Silva, F.J.; E Silva, R.P.; De Brito, M.E.F.; Filho, S.B.; Torres, F.D.; De Paiva-Cavalcanti, M. Detection of Leishmania infantum in animals and their ectoparasites by conventional PCR and real time PCR. Exp. Appl. Acarol. 2012, 59, 473–481. [Google Scholar] [CrossRef]
- Carvalho, E.M.; Bacellar, O. Imunopatogênese da Leishmaniose Visceral. Gaz. Méd. Bahia 2005, 75, 24–34. [Google Scholar]
- De Freitas, E.; Silva, R.; von Stebut, E. Unraveling the Role of Immune Checkpoints in Leishmaniasis. Front. Immunol. 2021, 12, 620144. [Google Scholar] [CrossRef]
- Christensen, S.M.; Belew, A.T.; El-Sayed, N.M.; Tafuri, W.L.; Silveira, F.T.; Mosser, D.M. Host and parasite responses in human diffuse cutaneous leishmaniasis caused by L. amazonensis. PLOS Neglected Trop. Dis. 2019, 13, e0007152. [Google Scholar] [CrossRef] [Green Version]
- Hong, A.; Zampieri, R.A.; Shaw, J.J.; Floeter-Winter, L.M.; Laranjeira-Silva, M.F. One Health Approach to Leishmaniases: Understanding the Disease Dynamics through Diagnostic Tools. Pathogens 2020, 9, 809. [Google Scholar] [CrossRef]
- Atayde, V.D.; Hassani, K.; Filho, A.D.S.L.; Borges, A.R.; Adhikari, A.; Martel, C.; Olivier, M. Leishmania exosomes and other virulence factors: Impact on innate immune response and macrophage functions. Cell. Immunol. 2016, 309, 7–18. [Google Scholar] [CrossRef] [PubMed]
- Gonçalves, G.A.; Eifler-Lima, V.L.; von Poser, G.L. Revisiting nature: A review of iridoids as a potential antileishmanial class. Phytochem. Rev. 2021, 1–26. [Google Scholar] [CrossRef]
- Giunchetti, R.C.; Martins-Filho, O.A.; Carneiro, C.M.; Mayrink, W.; Marques, M.J.; Tafuri, W.L.; Corrêa-Oliveira, R.; Reis, A.B. Histopathology, parasite density and cell phenotypes of the popliteal lymph node in canine visceral leishmaniasis. Veter- Immunol. Immunopathol. 2008, 121, 23–33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piña-Vázquez, C.; Reyes-López, M.; Ortíz-Estrada, G.; De La Garza, M.; Serrano-Luna, J. Host-Parasite Interaction: Parasite-Derived and -Induced Proteases That Degrade Human Extracellular Matrix. J. Parasitol. Res. 2012, 2012, 748206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mercado-Camargo, J.; Cervantes-Ceballos, L.; Vivas-Reyes, R.; Pedretti, A.; Serrano-García, M.L.; Gómez-Estrada, H. Homology Modeling of Leishmanolysin (GP63) from Leishmania panamensis and Molecular Docking of Flavonoids. ACS Omega 2020, 5, 14741–14749. [Google Scholar] [CrossRef]
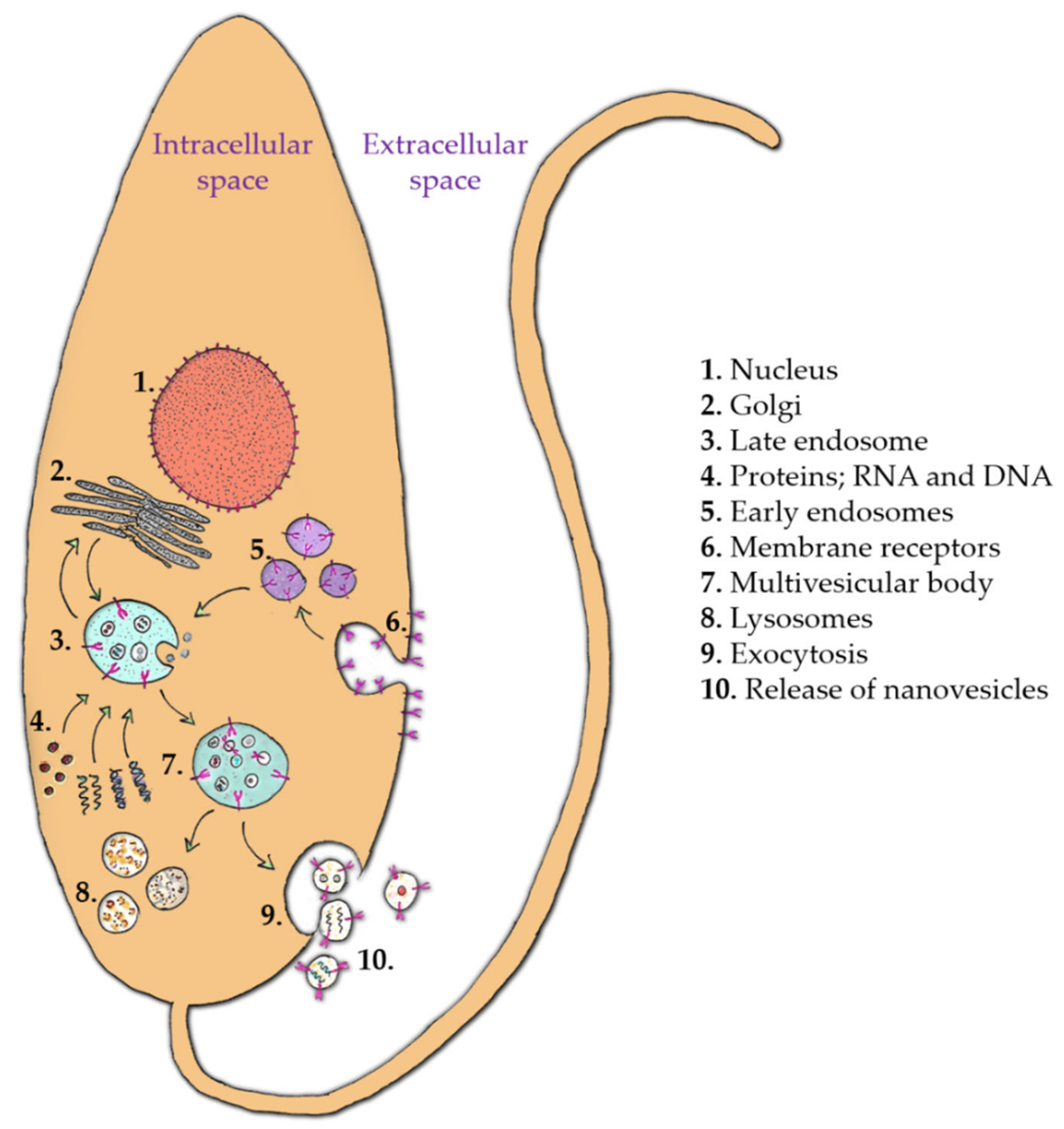
- Silverman, J.M.; Clos, J.; de’Oliveira, C.C.; Shirvani, O.; Fang, Y.; Wang, C.; Foster, L.J.; Reiner, N.E. An exosome-based secretion pathway is responsible for protein export from Leishmania and communication with macrophages. J. Cell Sci. 2010, 123, 842–852. [Google Scholar] [CrossRef] [Green Version]
- Monteiro, L.; Loebenberg, R.; Cotrim, P.; Fotaki, N.; Bou-Cracha, N. A potential nanotechnology based drug delivery for the treatment of Leishmaniases. In Proceedings of the AAPS Annual Meeting, Orlando, FL, USA, 25–29 October; 2015. [Google Scholar]
- Olivier, M.; Zamboni, D.S. Leishmania Viannia guyanensis, LRV1 virus and extracellular vesicles: A dangerous trio influencing the faith of immune response during muco-cutaneous leishmaniasis. Curr. Opin. Immunol. 2020, 66, 108–113. [Google Scholar] [CrossRef]
- Procházková, M.; Füzik, T.; Grybchuk, D.; Falginella, F.L.; Podešvová, L.; Yurchenko, V.; Vácha, R.; Plevka, P. Capsid Structure of Leishmania RNA Virus 1. J. Virol. 2021, 95, e01957-20. [Google Scholar] [CrossRef]
- Atayde, V.D.; Filho, A.D.S.L.; Chaparro, V.; Zimmermann, A.; Martel, C.; Jaramillo, M.; Olivier, M. Exploitation of the Leishmania exosomal pathway by Leishmania RNA virus 1. Nat. Microbiol. 2019, 4, 714–723. [Google Scholar] [CrossRef]
- Castiglioni, P.; Hartley, M.-A.; Rossi, M.; Prevel, F.; Desponds, C.; Utzschneider, D.; Eren, R.O.; Zangger, H.; Brunner, L.; Collin, N.; et al. Exacerbated Leishmaniasis Caused by a Viral Endosymbiont can be Prevented by Immunization with Its Viral Capsid. PLoS Neglected Trop. Dis. 2017, 11, e0005240. [Google Scholar] [CrossRef]
- Saberi, R.; Fakhar, M.; Hajjaran, H.; Ataei-Pirkooh, A.; Mohebali, M.; Taghipour, N.; Hezarjaribi, H.Z.; Moghadam, Y.D.; Bagheri, A. Presence and diversity of Leishmania RNA virus in an old zoonotic cutaneous leishmaniasis focus, northeastern Iran: Haplotype and phylogenetic based approach. Int. J. Infect. Dis. 2020, 101, 6–13. [Google Scholar] [CrossRef]
- Regev-Rudzki, N.; Wilson, D.W.; Carvalho, T.G.; Sisquella, X.; Coleman, B.M.; Rug, M.; Bursac, D.; Angrisano, F.; Gee, M.; Hill, A.F.; et al. Cell-cell communication between malaria-infected red blood cells via exosome-like vesicles. Cell 2013, 153, 1120–1133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szempruch, A.J.; Dennison, L.; Kieft, R.; Harrington, J.M.; Hajduk, S.L. Sending a message: Extracellular vesicles of pathogenic protozoan parasites. Nat. Rev. Genet. 2016, 14, 669–675. [Google Scholar] [CrossRef] [PubMed]
- Gabriel, Á.M.; da Rocha, L.C.; Passero, F.; Alexandre-Pires, G.; Santos-Gomes, G. Morfometria aplicada à micromorfologia para a caracterização de subpopulações heterogéneas de vesículas extracelulares de Leishmania. In Proceedings of the X Jornadas Cientificas do Instituto de Higiene e Medicina Tropical, IHMT–Lisbon, Portugal, December 2019; IHMT-UNL: Lisbon, Portugal, 2019. [Google Scholar]
- Rossi, I.V.; Nunes, M.A.F.; Vargas-Otalora, S.; da Silva Ferreira, T.C.; Cortez, M.; Ramirez, M.I. Extracellular Vesicles during TriTryps infection: Complexity and future challenges. Mol. Immunol. 2021, 132, 172–183. [Google Scholar] [CrossRef] [PubMed]
- Cruz-Saavedra, L.; Muñoz, M.; Patiño, L.H.; Vallejo, G.A.; Guhl, F.; Ramírez, J.D. Slight temperature changes cause rapid transcriptomic responses in Trypanosoma cruzi metacyclic trypomastigotes. Parasites Vectors 2020, 13, 255. [Google Scholar] [CrossRef]
- Atayde, V.D.; Aslan, H.; Townsend, S.; Hassani, K.; Kamhawi, S.; Olivier, M. Exosome Secretion by the Parasitic Protozoan Leishmania within the Sand Fly Midgut. Cell Rep. 2015, 13, 957–967. [Google Scholar] [CrossRef] [Green Version]
- Hassani, K.; Antoniak, E.; Jardim, A.; Olivier, M. Temperature-induced protein secretion by Leishmania mexicana modulates macrophage signalling and function. PLoS Negl. Trop. Dis. 2011, 6, e18724. [Google Scholar] [CrossRef] [Green Version]
- Gabriel Á, M.; Coelho da Rocha, L.; Passero, F.; Santos-Gomes, G. Interações experimentais entre macrófagos de roedores e nanovesículas de Leishmania. In Proceedings of the VIII Jornadas Cientificas do Instituto de Higiene e Medicina Tropical, IHMT, Lisbon, Portugal, 12 December 2017; IHMT-UNL: Lisbon, Portugal, 2017. [Google Scholar]
- Reis, N.F.D.C.; Dupin, T.V.; Costa, C.R.; Toledo, M.D.S.; De Oliveira, V.C.; Popi, A.F.; Torrecilhas, A.C.; Xander, P. Leishmania amazonensis Promastigotes or Extracellular Vesicles Modulate B-1 Cell Activation and Differentiation. Front. Cell. Infect. Microbiol. 2020, 10, 573813. [Google Scholar] [CrossRef]
- Schorey, J.S.; Cheng, Y.; Singh, P.P.; Smith, V.L. Exosomes and other extracellular vesicles in host–pathogen interactions. EMBO Rep. 2015, 16, 24–43. [Google Scholar] [CrossRef] [Green Version]
- Torrecilhas, A.C.; Soares, R.P.; Schenkman, S.; Fernández-Prada, C.; Olivier, M. Extracellular Vesicles in Trypanosomatids: Host Cell Communication. Front. Cell. Infect. Microbiol. 2020, 10, 602502. [Google Scholar] [CrossRef]
- Pérez-Cabezas, B.; Santarem, N.; Cecílio, P.; Silva, C.; Silvestre, R.; Catita, J.; Da Silva, A.C. More than just exosomes: Distinct Leishmania infantum extracellular products potentiate the establishment of infection. J. Extracell. Vesicles 2018, 7, 1541708. [Google Scholar] [CrossRef] [Green Version]
- Gabriel, Á.M.; Pedersen, K.W.; Santos-Gomes, G. Standardization of an extracellular vesicles research checklist in Medical Parasitology. In Proceedings of the XI Jornadas Cientificas do IHMT, Lisbon, Portugal, 10 December 2020; IHMT-UNL: Lisbon, Portugal, 2020. [Google Scholar]
- Théry, C.; Witwer, K.W.; Aikawa, E.; Alcaraz, M.J.; Anderson, J.D.; Andriantsitohaina, R.; Antoniou, A.; Arab, T.; Archer, F.; Atkin-Smith, G.K.; et al. Minimal information for studies of extracellular vesicles 2018 (MISEV2018): A position statement of the International Society for Extracellular Vesicles and update of the MISEV2014. J. Extracell. Vesicles 2018, 7, 1535750. [Google Scholar] [CrossRef] [Green Version]
- Soekmadji, C.; Li, B.; Huang, Y.; Wang, H.; An, T.; Liu, C.; Pan, W.; Chen, J.; Cheung, L.; Falcon-Perez, J.M.; et al. The future of Extracellular Vesicles as Theranostics—An ISEV meeting report. J. Extracell. Vesicles 2020, 9, 1809766. [Google Scholar] [CrossRef] [PubMed]
- Ambertz, U.; Ovando, M.E.O.; Vasconcelos, E.J.; Unrau, P.J.; Myler, P.J.; Reiner, N.E. Small RNAs derived from tRNAs and rRNAs are highly enriched in exosomes from both old and new world Leishmania providing evidence for conserved exosomal RNA Packaging. BMC Genom. 2015, 16, 151. [Google Scholar]
- Pedersen, K.W.; Kierulf, B.; Neurauter, A. Specific and Generic Isolation of Extracellular Vesicles with Magnetic Beads. Methods Mol. Biol. 2017, 1660, 65–87. [Google Scholar] [CrossRef]
- Sabatke, B.; Gavinho, B.; Coceres, V.; de Miguel, N.; Ramirez, M.I. Unveiling the role of EVs in anaerobic parasitic protozoa. Mol. Immunol. 2021, 133, 34–43. [Google Scholar] [CrossRef]
- Khosravi, M.; Mirsamadi, E.S.; Mirjalali, H.; Zali, M.R. Isolation and Functions of Extracellular Vesicles Derived from Parasites: The Promise of a New Era in Immunotherapy, Vaccination, and Diagnosis. Int. J. Nanomed. 2020, 15, 2957–2969. [Google Scholar] [CrossRef] [PubMed]
- Nievas, Y.R.; Lizarraga, A.; Salas, N.; Cóceres, V.M.; De Miguel, N. Extracellular vesicles released by anaerobic protozoan parasites: Current situation. Cell. Microbiol. 2020, 22, e13257. [Google Scholar] [CrossRef]
- Hui, W.W.; Emerson, L.E.; Clapp, B.; Sheppe, A.E.; Sharma, J.; del Castillo, J.; Ou, M.; Maegawa, G.H.B.; Hoffman, C.; Larkin, I.J.; et al. Antigen-encapsulating host extracellular vesicles derived from Salmonella-infected cells stimulate pathogen-specific Th1-type responses in vivo. PLoS Pathog. 2021, 17, e1009465. [Google Scholar] [CrossRef]
- Chawla, B.; Jhingran, A.; Panigrahi, A.; Stuart, K.D.; Madhubala, R. Paromomycin affects translation and vesicle-mediated trafficking as revealed by proteomics of paromomycin–susceptible and resistant Leishmania donovani. PLoS ONE 2011, 6, e26660. [Google Scholar] [CrossRef]
- Kaminski, V.D.L.; Ellwanger, J.; Chies, J.A.B. Extracellular vesicles in host-pathogen interactions and immune regulation—exosomes as emerging actors in the immunological theater of pregnancy. Heliyon 2019, 5, e02355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Brien, K.; Breyne, K.; Ughetto, S.; Laurent, L.C.; Breakefield, X.O. RNA delivery by extracellular vesicles in mammalian cells and its applications. Nat. Rev. Mol. Cell Biol. 2020, 21, 585–606. [Google Scholar] [CrossRef]
- Gill, S.; Catchpole, R.; Forterre, P. Extracellular membrane vesicles in the three domains of life and beyond. FEMS Microbiol. Rev. 2018, 43, 273–303. [Google Scholar] [CrossRef]
- Toledo, M.D.S.; Cronemberger-Andrade, A.; Barbosa, F.M.C.; Reis, N.F.D.C.; Dupin, T.V.; Soares, R.P.; Torrecilhas, A.C.; Xander, P. Effects of extracellular vesicles released by peritoneal B-1 cells on experimental Leishmania (Leishmania) amazonensis infection. J. Leukoc. Biol. 2020, 108, 1803–1814. [Google Scholar] [CrossRef]
- De Jesus, J.B.; Mesquita-Rodrigues, C.; Cuervo, P. Proteomics advances in the study of Leishmania parasites and leishmaniasis. Subcell Biochem. 2014, 74, 323–349. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Vega, A.; Losada-Barragán, M.; Berbert, L.R.; Mesquita-Rodrigues, C.; Bombaça, A.C.S.; Menna-Barreto, R.; Aquino, P.; Carvalho, P.C.; Padrón, G.; de Jesus, J.B.; et al. Quantitative analysis of proteins secreted by Leishmania (Viannia) braziliensis strains associated to distinct clinical manifestations of American Tegumentary Leishmaniasis. J. Proteom. 2020, 232, 104077. [Google Scholar] [CrossRef]
- Noor, Z.; Ahn, S.B.; Baker, M.; Ranganathan, S.; Mohamedali, A. Mass spectrometry-based protein identification in proteomics-a review. Briefings Bioinform. 2020, 22, 1620–1638. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, S.; Dutta, S.; Verma, A.S.; Saha, M.K. Chapter 34—Nanoparticle synthesis harnessing benign green routes. In Verma, Anchal Singh, Animal Biotechnology, 2nd ed.; Ashish, S., Ed.; Academic Press: Cambridge, MA, USA, 2020; pp. 689–708. ISBN 9780128117101. [Google Scholar] [CrossRef]
- Théry, C.; Clayton, A.; Amigorena, S.; Raposo, G. Current Protocols in Cell Biology Ch. 3 Exosomes from Cell Culture Supernatants and Biological Fluids; Bonifacino, J.S., Dasso, M., Harford, J.B., Lippincott-Schwartz, J., Yamada, K.M., Eds.; Wiley & Sons: New Jersey, NJ, USA, 2006; Volume 30, pp. 22–29. [Google Scholar]
- Deolindo, P.; Ramirez, M.I.; Evans-Osses, I. Microvesicles and exosomes as vehicles between protozoan and host cell communication. Soc. Trans. 2013, 41, 252–257. [Google Scholar] [CrossRef] [Green Version]
- Sharma, M.; Morgado, P.; Zhang, H.; Ehrenkaufer, G.; Manna, D.; Singh, U. Characterization of Extracellular Vesicles from Entamoeba histolytica Identifies Roles in Intercellular Communication That Regulates Parasite Growth and Development. Infect. Immun. 2020, 88, e00349-20. [Google Scholar] [CrossRef] [PubMed]
- Diaz-Garrido, N.; Cordero, C.; Olivo-Martinez, Y.; Badia, J.; Baldomà, L. Cell-to-Cell Communication by Host-Released Extracellular Vesicles in the Gut: Implications in Health and Disease. Int. J. Mol. Sci. 2021, 22, 2213. [Google Scholar] [CrossRef] [PubMed]
- Montaner, S.; Galiano, A.; Trelis, M.; Martin-Jaular, L.; Del Portillo, H.A.; Bernal, D.; Marcilla, A. The role of extracellular vesicles in modulating the host immune response during parasitic infections. Front. Immunol. 2014, 5, 433. [Google Scholar] [CrossRef]
- Hellard, E.; Fouchet, D.; Vavre, F.; Pontier, D. Parasite-Parasite Interactions in the Wild: How To Detect Them? Trends Parasitol. 2015, 31, 640–652. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.; Zhou, Z. Progress on the mechanism of extracellular vesicles involves in pathogen infection: Review. Chin. J. Cell. Mol. Immunol. 2020, 36, 467–473. [Google Scholar]
- Babatunde, K.; Subramanian, B.Y.; Ahouidi, A.D.; Murillo, P.M.; Walch, M.; Mantel, P.-Y. Role of Extracellular Vesicles in Cellular Cross Talk in Malaria. Front. Immunol. 2020, 11, 22. [Google Scholar] [CrossRef] [Green Version]
- Corrales, R.M.; Sereno, D.; Mathieu-Daudé, F. Deciphering the Leishmania exoproteome: What we know and what we can learn. FEMS Immunol. Med. Microbiol. 2010, 58, 27–38. [Google Scholar] [CrossRef] [Green Version]
- Gabriel, Á.M.; Rafael-Fernandes, M.; Passero, F.; Pires, G.A.; Santos-Gomes, G. Inserts on comparative scanning electron microscope study of Leishmania exosomes. In Proceedings of the VI WORLDLEISH, Toledo, Spain, 16–20 May 2017. [Google Scholar]
- Wheeler, R.J.; Sunter, J.D.; Gull, K. Flagellar pocket restructuring through the Leishmania life cycle involves a discrete flagellum attachment zone. J. Cell Sci. 2016, 129, 854–867. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jain, S.; Santana, W.; Dolabella, S.; Santos, A.; Souto, E.; Severino, P. Are Nanobiosensors an Improved Solution for Diagnosis of Leishmania? Pharmaceutics 2021, 13, 491. [Google Scholar] [CrossRef]
- Lässer, C.; Eldh, M.; Lötvall, J. Isolation and characterization of RNA-containing exosomes. J. Vis. Exp. 2012, 59, e3037. [Google Scholar] [CrossRef]
- Valverde, J.; Paun, A.; Inbar, E.; Romano, A.; Lewis, M.; Ghosh, K.; Sacks, D. Increased Transmissibility of Leishmania donovani From the Mammalian Host to Vector Sand Flies After Multiple Exposures to Sand Fly Bites. J. Infect. Dis. 2017, 215, 1285–1293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Terrazas, C.; Varikuti, S.; Kimble, J.; Moretti, E.; Boyaka, P.N.; Satoskar, A.R. IL-17A promotes susceptibility during experimental visceral leishmaniasis caused by Leishmania donovani. FASEB journal: Official publication of the Federation of American Societies for Experimental Biology. FASEB J. 2015, 30, 1135–1143. [Google Scholar] [CrossRef] [Green Version]
- Passelli, K.; Billion, O.; Tacchini-Cottier, F. The Impact of Neutrophil Recruitment to the Skin on the 914 Pathology Induced by Leishmania Infection. Front. Immunol. 2021, 12, 649348. [Google Scholar] [CrossRef] [PubMed]
- Ready, P.D. Biology of phlebotomine sand flies as vectors of disease agents. Annu. Rev. Entomol. 2013, 58, 227–250. [Google Scholar] [CrossRef] [PubMed]
- Théry, C. Exosomes: Secreted vesicles and intercellular communications F1000. Biol. Rep. 2011, 3, 15. [Google Scholar] [CrossRef] [PubMed]
- Li, M.-Y.; Liu, L.-Z.; Dong, M. Progress on pivotal role and application of exosome in lung cancer carcinogenesis, diagnosis, therapy and prognosis. Mol. Cancer 2021, 20, 22. [Google Scholar] [CrossRef]
- Olivier, M.; Gregory, D.J.; Forget, G. Subversion mechanisms by which Leishmania parasites can escape the host immune response: A signaling point of view. Clin. Microbiol. Rev. 2005, 18, 293–305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Basmaciyan, L.; Robinson, D.R.; Azas, N.; Casanova, M. (De)glutamylation and cell death in Leishmania parasites. PLoS Neglected Trop. Dis. 2019, 13, e0007264. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Geographic Distribution | Clinical Disease in Humans | Hosts | Experimental Activatory Stimuli | LEVs Isolation Methods | LEVs type and Sub-Cellular Origin | Size (Mean/ Range) | Major LEVs Content | Method of LEVs Analysis |
|---|---|---|---|---|---|---|---|---|---|
| Complex | Vector: Phlebotomine sand fly species | Microparticles, | |||||||
| L. (L.) donovani | Central and southern Sudan, Northern Kenya, south-east Ethiopia, Uganda, Chad, Yemen, North-east India, Bangladesh, Terai region of Nepal, Buthan, China | VL, PKDL | Infect wild and synathropic rodents, wild felids, viverrids. Cats (Felis silvestris catus)? Presumably zoonotic, but reservoir host uncertain/unknown. Human (peridomestic) and Herpestids Mongooses (zoonotic) | Neutral and acid medium; RPMI with HEPES, MES; | 4, 2, 7 | (Exosomes, vesicles); | 30–70 nm | HSP100, 90, 70.4, GP63, histone, chaperonin, proteins; TESA, trans-sialidases, protease transport, metabolic protein | Immunofluorescence (IF) and EM; Proteomic Analysis |
| L. (L.) donovani HSP100/and | |||||||||
| Wildtype (Exosomes) | |||||||||
| From PM, FP, phagolysosome | |||||||||
| L. (L.) infantum (syn. of L. L. chagasi) | Central and western Mediterranean basin, both Europe and North Africa Through Mediterranean basin to Iran, South west Asia, China Central and South America | VL, CL (zoonotic), Infantile visceral leishmaniasis, AIDS associated leishmaniasis | Vectors: Several species of Phlebotomus and Lutzomyia sandflies Infect wild canids, synanthropic rodents, cats and humans Dogs are reservoir hosts (viscerocutaneous leishmaniasis) Landscape peridomestic | Miltefosine/apoptosis indicers, G418; Insect gut; RPMI pepton Yeast; Exo-free medium [77,82,114] | 5,3; 11; 1,4; 8 | Vesicles, Exosome-like Vesicles, Exosome Vesicles | 30–100 nm; 50–120 nm; 122 + −56 nm; 30–450 nm | GP63, ribossomal protein, hsp70 elongation factor-1 α, β tubulin, β-fructofuranosidases; HSP70, HSP83/90, Acetylcholinesterase; GP63, calpan-like cysteine peptidase, HSP70, tryparedoxin peroxidase surface antigen protein; Nucleic acids and proteins (protein levels ranged from 40 ± 76 mg.mL−1, RNA concentration reached mean values of 90 ± 277 ng.µL−1 and DNA of 85 ± 377 ng.µL−1) | Experimental infection [77,82,114] |
| From membrane surface, FP, MVB; Extracellular vesicles, nanovesicles, exosomes [77,82,114] | 115 + −65 nm | ||||||||
| L. (L.) major | Sub-Saharan Africa, Yemen, North Africa, Middle East. South-west Asia, from Algeria to Saudi Arabia. Iran and neighbours, Pakistan, Northwest India. Central Asia from Iran to Uzbekistan, west Africa to Kenya, Sahel belt | CL, Oriental sore (wet form) | Vector: Phlebotomine sand fly species (Phlebotomus) Infect numerous desert mammals, wild rodents, cats, dogs and humans Reservoir hosts: Fat sand-rat Psammomys obesus (epidemic maintained by Meriones shawi), great gerbil Rhombomys opimus, regional gerbils and rodents Relative importance of different hosts to be determined | Neutral and acid medium; Insect gut; | 11; 4 | Microparticles, (Exosomes, vesicles); Exosome-like Vesicles | 30–70 nm; 50–120 nm | GP63, calpan-like cysteine peptidase, HSP70, tryparedoxin peroxidase surface antigen protein; TESA, trans-sialidases, protease transport, metabolic protein | Proteomic Analysis |
| From membrane surface, FP, MVB, PM, phagolysosome | |||||||||
| L. (L.) amazonensis (syn. of L. L. garnhami) | South America, mostly North of the Amazon, East of Andes | CL, DCL, MCL | Vector: Phlebotomine sand fly species (Lutzomyia) Infect terrestrial forest rodents, marsupials, procyonids, wild canids, edentates, sciuridi and humans (sylvatic) Terrestrial rodents and marsupials, Proechymis guyanensis and Proechymis cuvieri (reservoir hosts) | RPMI/20% glucose; Exo-free medium [77,78,79,80,81,82,83,84,85,86,87,88,89,90,91,92,93,94,95,96,97,98,99,100,101,102,103,104,105,106,107,108,109,110,111,112,113,114] | 2,4; 1;8 | L. (L.) amazonensis P (-M2269) (Evs) whole body; Extracellular vesicles, nanovesicles, exosomes [77,82,114] | 180 nm; 30–450 nm | GP63, LPG; Nucleic acids and proteins (protein levels ranged from 40 ± 76 mg·mL−1, RNA concentration reached mean values of 90 ± 277 ng.µL−1 and DNA of 85 ± 377 ng.µL−1) | Experimental infection [77,82,114] |
| L. (L.) mexicana (syn. of L. L. pifanoi) | Southern USA (Texas), Central and South America, East of Andes | CL, DCL, Chiclero ulcer | Vector: Phlebotomine sand fly species (Lutzomyia) Infect rodents, edentates, marsupials, wild canids and humans (sylvatic) Marsupials and terrestrial rodents: Ototylomys phyllotis and Neotoma micropus (reservoir hosts) | Neutral and acid medium | 4 | Microparticles, | 30–70 nm | TESA, trans-sialidases, protease transport, metabolic protein | Proteomic Analysis |
| (Exosomes, vesicles) | |||||||||
| From PM, FP, phagolysosome | |||||||||
| L. (L.) shawi | Brazilian Amazon Region | CL | Vector: Phlebotomine sand fly species (Lutzomyia) Infect monkeys: Cebus apella, Chiropotes satanas; edentates: Choloepus didactylus, Bradypus tridactylus; procyonids: Nasua nasua; humans (sylvatic) Main host uncertain | Exo-free medium [77,82,114] | 1; 8 | Extracellular vesicles, nanovesicles, exosomes [77,82,114] | 30–450 nm | Nucleic acids and proteins (protein levels ranged from 40 ± 76 mg.mL−1, RNA concentration reached mean values of 90 ± 277 ng.µL−1 and DNA of 85 ± 377 ng.µL−1) | Experimental infection [77,82,114] |
| L. (V.) braziliensis | Central & South America | CL, MCL | Vector: Phlebotomine sand fly species (Lutzomyia) Infect wild and synanthropic rodents, marsupials: Didelphis marsupialis and others numerous forest animals; dogs (CL), cats, horses, donkeys Humans. (peridomestc and sylvatic) Presumably zoonotic, but reservoir host unknown Rodents? Marsupials? Dogs? | 199 medium (Gibco, Life Technologies Brand, Grand Island, NY, USA) supplemented with 4.2 mM sodium bicarbonate, 4.2 mM HEPES, 1 mM adenine, 5 μg/mL hemin (bovine type I) (Sigma, St. Louis, MO, USA) and 10% fetal calf serum (FCS) (Gibco, Carlsbad, CA, USA) [1] | 5 | Extracellular vesicles | *** | *** | Proteomic Analysis |
| L. (V.) guyanensis | South America, East of Andes, Guyanas | CL, MCL | Vector: Phlebotomine sand fly species (Lutzomyia) Infect wild rodents, edentates: Choloepus didactylus, Tamandua tetradactyla; marsupials, humans (zoonotic) Sloth Choloepus didactylus (reservoir host) and probably other animals (sylvatic) | Exo-free medium [77,82,114] | 1; 8 | Extracellular vesicles, nanovesicles, exosomes [77,82,114] | 30–450 nm | Nucleic acids and proteins (protein levels ranged from 40 ± 76 mg·ml−1, RNA concentration reached mean values of 90 ± 277 ng.µL−1 and DNA of 85 ± 377 ng.µL−1) | Experimental infection [77,82,114] |
| Parameters | Mandatory Steps (Standardized) | Mandatory if Applicable | Encouraged Steps |
|---|---|---|---|
| Nomenclature | (a) We propose to use the generic term Leishmania extracellular vesicles (LEVs): With demonstration of extracellular (no intact parasites) and vesicular nature per these characterization and function guidelines | No additional information available | 1. Use the generic term Leishmania extracellular vesicles (LEVs) + specification (considering size, density, others) |
| (b) Generic term Leishmania extracellular particles (LEPs): No intact parasites but MISEV guidelines not satisfied | 1. Use a specific term for subcellular origin: Leishmania Ectosomes, microparticles, microvesicles (from plasma membrane), exosomes (from endosomes), with demonstration of the subcellular origin 2. Use other specific term: with definition of specific criteria | ||
| Collection and pre-processing culture conditioned medium (CM) General cell characterization medium used before and during collection (additives, serum, other) | (a) Nature and size of culture vessels, and volume of medium during conditioning | 1. Exact protocol for depletion of LEVs/LEPs from additives in collection medium | No additional information available |
| (b) Number of parasites/mL, or/surface area and % of live/dead cells at time of collection (or at time of seeding with estimation at time of collection) (c) Frequency and interval of CM harvest | 2. Specific culture conditions (treatment, % O2, others) before and during collection | ||
| Collection and pre-processing Storage and recovery | (a) Storage and recovery (e.g., thawing) of culture medium, before LEVs isolation (storage temperature, vessel, time; method of thawing or other sample preparation) | No additional information available | No additional information available |
| (b) Storage and recovery of LEVs after isolation (temperature, vessel, time, additive(s) others) | |||
| LEVs separation and concentration Experimental details of the method | No additional information available | (a) Centrifugation: reference number of tube(s), rotor(s), adjusted k factor(s) of each centrifugation step (= time + speed + rotor, volume/density of centrifugation conditions), temperature, brake settings | No additional information available |
| (b) Density gradient: nature of matrix, method of generating gradient, reference (and size) of tubes, bottom-up (sample at bottom, high density) or top-bottom (sample on top, low density), centrifugation speed and time (with brake specified), method and volume of fraction recovery | |||
| (c) Chromatography: matrix (nature, pore size, others), loaded sample volume, fraction volume, number | |||
| (d) Precipitation: references, ratio vol/vol or weight/vol fluid, time/temperature of incubation, time/speed/temperature of centrifugation | |||
| (e) Filtration: reference of filter type (= nature of membrane, pore size, others, time and speed of centrifugation, volume before/after (in case of concentration) | |||
| (f) Antibody-based: reference of antibodies, mass Ab/amount of LEVs, nature of Ab carrier (bead, surface) and amount of Ab/carrier surface | |||
| Other: all necessary details to allow replication | |||
| Additional step(s) to concentrate | |||
| Additional step(s) to wash matrix and/or sample | |||
| LEVs separation and concentration Specify category of the chosen LEVs separation/concentration method | No additional information available | No additional information available | 1. High recovery, low specificity = mixed LEVs and non-LEVs components |
| 2. Intermediate recovery, intermediate specificity = mixed LEVs with limited non-LEVs components | |||
| 3. Low recovery, high specificity = subtype(s) of LEVs with as little non- LEVs as possible | |||
| 4. High recovery, high specificity = subtype(s) of LEVs with as little non-LEVs as possible | |||
| LEVs characterization (Quantification) | (a) Volume of fluid, and/or cell number, and/or tissue mass used to isolate LEVs | No additional information available | No additional information available |
| (b) Global quantification by at least 2 methods: protein amount, particle number, lipid amount, expressed per volume of initial fluid or number of producing cells/mass of tissue Ratio of the 2 quantification figures | |||
| LEVs characterization (Global characterization) | (a) Transmembrane or GPI-anchored protein localized in cells/parasites at plasma membrane or endosomes | 1. Presence of proteins associated with compartments other than plasma membrane or endosomes | Topology of the relevant functional components |
| (b) Cytosolic protein with membrane-binding or-association capacity Assessment of presence/absence of expected contaminants | 2. Presence of soluble secreted proteins and their likely transmembrane ligands | ||
| LEVs characterization Single LEVs characterization | (a) Images of single LEVs by wide-field and close-up: e.g., EM, scanning probe microscopy, superresolution fluorescence microscopy | No additional information available | No additional information available |
| (b) Non-image-based method analysing large numbers of single LEVs: NTA (Nanoparticle tracking analysis), Tunable resistive pulse sensing (TRPS), Fluorescence Correlation Spectroscopy (FCS), high-resolution flow cytometry, multi-angle light-scattering, Raman spectroscopy, along with others. | |||
| Functional studies | (a) Dose-response assessment | No additional information available | 1. Quantitative comparison of activity of LEVs subtypes (if subtype-specific function claimed) |
| (b) Negative control = nonconditioned medium, biofluid/tissue from control donors, as applicable | |||
| (c) Quantitative comparison of functional activity of total fluid, vs. LEVs-depleted fluid, vs. LEVs (after high recovery/low specificity separation) | 2. Extent of functional activity in the absence of contact between LEVs donor and LEVs recipient | ||
| (d) Quantitative comparison of functional activity of LEVs vs. other LEPs/fractions after low recovery/high specificity separation | |||
| Reporting | Submission of data (proteomic, sequencing, others) to relevant public, curated databases or open-access repositories | Temper LEVs-specific claims when MISEV requirements cannot be entirely satisfied | 1. Submission of methodologic details to LEVs-TRACK (evtrack.org) with EVs-TRACK number provided (strongly encouraged) |
| 2. Data submission to LEVs -specific databases (e.g., EVpedia, Vesiclepedia, exRNA atlas, Eco Carta, exoRBase,) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gabriel, Á.M.; Galué-Parra, A.; Pereira, W.L.A.; Pedersen, K.W.; da Silva, E.O. Leishmania 360°: Guidelines for Exosomal Research. Microorganisms 2021, 9, 2081. https://doi.org/10.3390/microorganisms9102081
Gabriel ÁM, Galué-Parra A, Pereira WLA, Pedersen KW, da Silva EO. Leishmania 360°: Guidelines for Exosomal Research. Microorganisms. 2021; 9(10):2081. https://doi.org/10.3390/microorganisms9102081
Chicago/Turabian StyleGabriel, Áurea Martins, Adan Galué-Parra, Washington Luiz Assunção Pereira, Ketil Winther Pedersen, and Edilene Oliveira da Silva. 2021. "Leishmania 360°: Guidelines for Exosomal Research" Microorganisms 9, no. 10: 2081. https://doi.org/10.3390/microorganisms9102081
APA StyleGabriel, Á. M., Galué-Parra, A., Pereira, W. L. A., Pedersen, K. W., & da Silva, E. O. (2021). Leishmania 360°: Guidelines for Exosomal Research. Microorganisms, 9(10), 2081. https://doi.org/10.3390/microorganisms9102081